植物脂肪氧化酶的特性與功能
2019-04-22 11:34:02謝鴻光許惠濱潘麗燕江敏榕魏林艷張建福
福建稻麥科技 2019年1期
謝鴻光 ,許惠濱 ,潘麗燕 ,連 玲 ,江敏榕 ,魏林艷 ,張建福
(1.福建省農業科學院水稻研究所,福建福州350018;2.農業部華南雜交水稻種質創新與分子育種重點實驗室/福州(國家)水稻改良分中心/福建省作物分子育種工程實驗室/福建省水稻分子育種重點實驗室/福建省作物種質創新與分子育種省部共建國家重點實驗室培育基地/雜交水稻國家重點實驗室華南研究基地/水稻國家工程實驗室,福建福州350003)
脂肪氧化酶(LOXs,亞油酸:氧化還原酶)含有非血紅素鐵或錳,是一類專一催化多元不飽和脂肪酸(如亞油酸、亞麻酸和花生四烯酸)加氧反應的雙加氧酶,其催化的多元不飽和脂肪酸含有順,順-1,4-戊二烯結構,生成的多元不飽和脂肪酸的氫過氧化物含有共軛雙鍵[1]。LOXs大量存在于動物、植物和微生物中[2],在植物中,它主要催化含有18個碳的亞油酸 (Linoleic acid)和亞麻酸(Linolenic acid)的反應[3];在動物中,它則是以20個碳的花生四烯酸 (Arachidonic acid)為底物。通常把由LOXs及其后續一系列反應催化多元不飽和脂肪酸(PUFA)的代謝過程稱為LOX途徑(Lipoxygenase pathway)[4],植物體內 LOX途徑的主要過程如圖1所示。
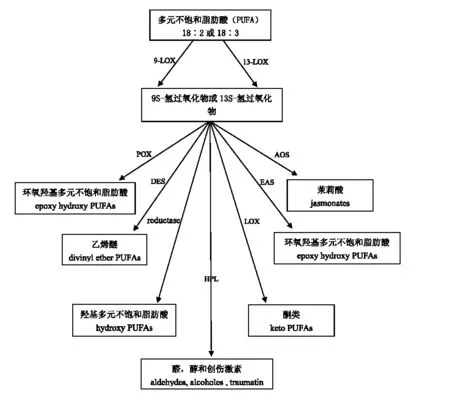
圖1 植物體內LOX途徑
目前的研究發現在大豆、花生、擬南芥、黃瓜、大麥、豌豆、番茄和水稻等植物中均存在LOXs,其表達受各種因素影響,如生長階段、激素、非生物脅迫和真菌感染等[5]。此外,從不同的植物分離出的LOXs基因具有不同的組織特異性表達,不同的植物種類及其不同的生長階段LOXs的活性也隨之變化[3]。亞細胞結果表明,植物中的LOXs主要分布于脂體、微粒體、葉綠體、液泡、質膜和胞漿中,同時在脂體膜和質膜上也有發現,它們主要以跨膜蛋白的形式存在[6,7]。在植物中,LOXs催化底物產生手性與位置專一性不同的氫過氧化基,因此可將其分為3種類型[7]:第1類是催化底物生成9S-氫過氧化物(9-HPOD)的9-氫過氧化亞油酸(9-LOX),如番茄果實和馬鈴薯塊莖中的LOX、豌豆種子以及水稻種胚中的LOX3;第2類是催化底物生成13S-氫過氧化物(13-HPOD)的13-氫過氧化亞油酸(13-LOX),如大豆種子的LOX1和豌豆種子的LOX2;第3類是9/13-LOX,它是一種非傳統的LOX,生成等量的9-HPOD和13-HPOD[8],如大豆種子中的LOX2和LOX3。
近年來,伴隨越來越多研究小組的加入,人們對植物LOXs及其反應產物氧化脂(oxylipins)的了解越來越深入。此外,克隆了一系列植物LOXs基因序列,為進一步揭示它們的系統發育并闡明LOXs基因序列與結構、區域選擇性和活性關系提供了可能。綜述了有關植物LOXs的生理學作用。
1 調控植物生長發育
研究表明,在大豆生長發育過程中,其體內的LOXs含量不斷變化著,當其組織處于快速生長階段時,其體內的LOXs活性很高,因此,研究者認為在大豆生長發育過程中,LOXs發揮重要的調控作用。此外,證明LOXs調控植物的生長發育的證據還來自Porta等人[9],他們在處于生長發育的菜豆根瘤中檢測到LOXs活性,并驗證到其相應的mRNA的表達,當根瘤生長停止的時候,LOXs及其相應的mRNA的含量都是不斷地下降,直到消失為止,因此,Porta 等人推測菜豆根瘤的發育可能受LOXs的調控。
馬鈴薯LOX1基因是一個具有塊莖專一性表達的基因,其編碼產物具有9-LOX活性,原位雜交結果證實其主要在發育的塊莖中表達。2001年,Kolomiets等[10]通過研究轉反義LOX1基因的馬鈴薯植株,結果發現其塊莖的產量明顯下降,而且形狀畸形,表明LOX1參與調控塊莖的發育和生長,并可能與LOXs的反應產物羥脂(oxylipin)有關。當然,也不排除LOXs調控種子萌發和根瘤發育的可能性[3]。
2 調節植物組織衰老
在植物中,脂質過氧化及其產生的自由基導致其膜降解進而引起組織衰老,而脂肪氧化酶及其催化產生的氫過氧化物可能直接參與了以上反應[11]。LOXs還參與啟動了組織的膜脂過氧化作用[12],在促進酶的失活反應的同時,還抑制葉綠體的光化學活性,最終加速細胞膜的降解[13];其催化產生的自由基加劇細胞組分降解,促進組織衰老。
研究發現,LOXs與植物果實成熟和衰老密切相關,但對其生理作用機理的研究還不是很透徹。我們知道乙烯(C2H4)調節果實后熟進程,人為調控其合成可實現此目的。而C2H4的合成需要有自由基及過氧化物的參與,LOXs催化所產生的超氧自由基正好可以參與該反應[14]。另外,茉莉酸(JA)和脫落酸(ABA)也是促進植物組織衰老的生長調節劑,它們的合成同樣需要有LOXs的參與[3,15]。番茄果實成熟伴隨著LOXs活性的增強,人為施加LOXs能夠使果實組織電導率增加,從而促進果實的成熟與衰老;研究人員還發現外源施加乙烯促使番茄果實 LOXs活性增強[16]。獼猴桃的果實采摘后初始階段,伴隨著LOXs活性不斷增強,啟動了膜脂的過氧化反應,不斷消耗底物亞麻酸和亞油酸,且不斷釋放出游離的脂肪酸,最終引起亞麻酸和亞油酸的積累[17]。此外,在蘋果儲存過程中,LOXs活性與乙烯合成量呈正相關,它不僅啟動果實成熟反應,還誘導合成乙烯,加速果實的成熟與衰老[7]。
3 防御病蟲害
前人的研究表明,LOX途徑與植物對病蟲害的防衛反應息息相關。其產生的茉莉酸(JA)是參與植物轉導防御反應的重要的信號因子;此外,生成的多種脂肪酸的氧化物,包括二乙烯乙醚類(divinylethers)和醛類(aldehydes),都具有抗病菌和蟲害的活性[18-20]。
當植物受到害蟲與病原菌侵染時,引起其體內的LOXs活性及其代謝產物的含量不斷地提高,包括脂氫過氧化物、氧化自由基和醛、酮等揮發性物質等,它們都可以實現抑制病蟲生長以及抵御病原菌侵染的目的,具體表現為在抗病植株體內LOXs的活性比敏感植株高很多。當大豆葉片受傷后,其體內的LOXs酶活性急劇增強[21];后面的研究又證實水稻稻瘟病菌[22]以及煙草 P.parasitica[23]都可以誘導LOXs的表達,這說明許多植物中LOXs的表達均受蟲害和病原微生物的誘導。以上現象有兩種可能的解釋:① LOX途徑產生的代謝物具有抗真菌活性,能夠抑制真菌的生長;② 啟動LOX途徑誘導植物過敏性反應(Hypersensitive reaction,HR)。前人的研究表明,LOXs活性增高和HR反應具有相關性,其可能原因是LOXs催化不飽和脂肪酸進而生成的脂氫過氧化物很不穩定,且極易通過均裂反應而生成大量的活性氧自由基,參與細胞膜脂的過氧化,從而破壞質膜,最終導致細胞壞死[24,25]。同時HR的進行需要激活9-LOX和 13-LOX[26],其中,9-LOX主要負責脂肪酸的過氧化,而13-LOX則負責JA的合成,這進一步說明HR的發生依賴于LOXs的存在。
4 作為儲藏蛋白
LOXs普遍存在于植物種子中,然而,尚未出現相關的研究表明其對種子發育發揮特定的功能。在非種子表達的植物中,LOXs可以以營養生長儲藏蛋白(vegetative storage protein, VSP)的形式存在,如大豆葉肉細胞液泡中營養器官的儲藏蛋白就是LOXs。在大豆種子發育過程中,3種LOX的含量占全部可溶性蛋白的1%,到萌發的早期它們就完全消失。同時在大豆子葉中出現三種新的LOX同工酶,它們均不是位于脂體膜上,也不參與儲藏脂質的降解。以上結果表明植物種子LOXs可能具有種子儲藏蛋白的功能[3]。
5 參與種子儲藏
在植物的生活史中,種子是重要的階段之一,且在植物繁衍中發揮著不可替代的作用。在儲藏過程中,通常隨著儲藏時間的延長,種子會不斷老化劣變,活力不斷下降。產生此現象最主要的一個原因是細胞膜完整性的喪失,而細胞膜完整性喪失的關鍵因素則是過氧化物、自由基以及膜脂過氧化代謝產物的產生和毒害作用,它們是引起植物組織衰老和種子劣變的主要原因[27]。脂體(lipid body,又稱油體,oil body)是一種由磷脂單分子層包裹的儲藏甘油三酯的球體結構,也是儲藏脂類聚集的主要細胞器,而LOXs則是定位在脂體膜上催化儲藏脂類轉運和降解的限速酶。
6 在食品加工中的應用
LOXs普遍存在于各種食品原料中,一方面其催化的生化反應可能影響食品加工品質;另一方面,其代謝產物可能降低食品的營養價值,主要是通過直接與食品中的有效成分(氨基酸和蛋白質)結合,因此,研究人員建議在食品加工和儲藏過程中應盡可能降低LOXs含量與活性[28]。
大豆由于其富含營養而備受青睞,而在大豆的加工過程中,LOXs通過分解不飽和脂肪酸產生小分子揮發性物質醛、酮和醇等,這就是豆腥味產生的原因所在。為此,前人大量研究如何通過降低大豆LOXs活性來消除豆制品的豆腥味,最后發現,豆腥味的產生與大豆體內LOXs的活性存在正相關。研究發現通過改變介質的pH值、加入醛水解酶、微波處理和有機溶劑萃取等均可降低豆腥味。但這些方法卻增加了豆制品的加工成本,還引起大豆蛋白水溶性下降。因此,應該從根本上解決此問題,即培育低LOXs含量的大豆品種。1999年,麻浩等[29]利用誘變技術培育了一個純合的LOX缺陷型大豆新品種。研究發現,蠶豆粉或大豆粉中的LOXs可以與面粉中的類胡蘿卜素發生偶聯反應漂白面團,因此,可以往面粉中添加些許的蠶豆粉或大豆粉,其中的LOXs不但可以漂白面粉,還能夠提高焙烤食品的質量。LOXs還具有使食品發生顏色變化的作用,比如苜蓿加工飼料中的葉黃素及其它有色類胡蘿卜素的降解,冷凍和加工蔬菜中葉綠素的破壞,還有添加于食品中的色素的降解等[30]。而LOXs提高烘焙食品質量的原理主要包括兩個方面:一、在面筋蛋白質中形成二硫鍵-S-S,從而引起蛋白質構象變化并使帶電基團轉向蛋白質表面,改變面團的流變性;二、氧化面筋蛋白,釋放結合的脂肪,防止脂肪的結合反應,促進游離脂肪數量增加,有效改進面包的柔軟度[31]。
另外,LOXs還應用于茶葉加工過程。人們在紅茶和烏龍茶的發酵過程中利用LOXs來催化亞麻酸和亞油酸的氧化分解,生成茶葉特有的香氣成分,包括已烯醇、已烯醛和正己醛等。
7 討論
脂質過氧化是所有生物系統中常見的一種代謝反應,出現在生長發育過程中,調控生物和非生物脅迫的響應機制。脂質過氧化的初始階段可以通過化學反應促使其發生,也可來自酶的催化作用。在植物中,這種反應主要是由脂肪氧化酶催化的。LOXs亞型的數量、結構和功能的多樣性使植物能夠適當地響應環境的挑戰。JA及其十八烷酸前體是第一個具有信使功能的氧化脂蛋白。越來越多的研究表明這些化合物在植物生長發育和脅迫反應中起“主開關”作用。此外,其他的氧化脂蛋白作為公認的信號分子也逐漸受到關注。
近幾年來,對LOX通路復雜性及其生理意義的了解顯著增加。對不同植物LOXs的研究為揭示它們的機制奠定了基礎。反向遺傳學方法為脂肪氧化酶功能的研究提供了初步的見解。此外,氧化脂蛋白生物合成突變體的產生,以及編碼LOX途徑相關酶基因的持續過表達和失活,有望拓寬我們對脂肪氧化酶參與植物發育與逆境適應的生理功能的認識。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
