不同鹽脅迫條件下東方杉的生長及生理響應研究
2019-04-16 02:48:24馬曉華宮莉霞張旭樂項德強
西南林業大學學報 2019年2期
孔 強 馬曉華 宮莉霞 張旭樂 項德強 鄭 堅
( 1. 樂清市林業局,浙江 溫州 325600;2. 浙江省亞熱帶作物研究所,浙江 溫州 325005)
東方杉(Taxodium mucronatum×Cryptomeria fortunei)為杉科落羽杉屬,半常綠的高大喬木,是1963年我國著名林木育種家葉培忠教授將中國柳杉和墨西哥落羽杉進行跨屬雜交育成的杉科新樹種[1]。東方杉樹形優美,適應性十分廣泛,其具有生長迅速、耐鹽堿和耐水濕能力強等特點,且具有較強的抗風能力[2],因此東方杉近年在沿海防護林和生態公益林的建設中逐步受到青睞。
有研究表明,沿海地帶土壤主要表現為鹽堿含量高、表層板結、海水倒灌反鹽,十分不利于植物的生長發育[3]。目前,我國擁有大面積的鹽堿地,僅海岸帶、灘涂地等就達670多萬hm2,且有逐年遞增的趨勢[4]。在我國人口不斷增加,耕地面積日趨緊張和淡水資源不斷減少的嚴峻形勢下,鹽堿地的開發利用已是國內外生物學者迫切需要解決的研究課題。在鹽堿地土壤對植物的危害中通常認為在無機鹽對植物生長的危害是最顯著的,大部分植物或農作物都對NaCl極為敏感,即使是在低鹽度條件下的 50 mmol/L NaCl對于具有一定耐鹽能力的水稻(Oryza sativa)也具有致死作用[5]。本研究對2年生東方杉鹽脅迫試驗,測定和分析其在鹽脅迫環境下的各項形態及生理指標,以期探討鹽脅迫條件下東方杉的生長及生理變化,為開發及選育耐鹽性強的植物資源提供依據。
1 材料與方法
1.1 試驗材料
2015 年4 月,于浙江溫州(119°37′~121°18′E;27°06′~28°36′N)智能溫室進行東方杉耐鹽性試驗,選用健壯且長勢一致(苗高為(62±0.5)cm,地徑為(68±0.5)mm)的2年生東方杉,在光強為1000 μmol /(m2·s), 溫度為 28 ℃/25 ℃(晝/夜),光周期為12 h,濕度為(70 ± 5)%的條件下進行水培試驗研究。營養液配比按照霍格蘭營養液作一定調整后配制。營養液配方如下:Ca(NO3)2·4H2O 2.00 mmol/L,KH2PO40.10 mmol /L,MgSO4·7H2O 0.50 mmol /L,KCl 0.10 mmol/L,K2SO40.70 mmol/L,H3BO310.00 μmol/L,MnSO4·H2O 0.50 μmol/L,ZnSO4·7H2O 1.00 μmol/L,CuSO4·5H2O 0.20 μmol/L,(NH4)6Mo7O240.01 μmol/L,Fe-EDTA 100 μmol /L。
1.2 試驗方法
東方杉植株適應生長2個月后,開始進行不同濃度的鹽處理。選用容積為10 L的塑料桶,每桶1株,培養液體積為7.5 L。試驗設置0‰(CK)、3‰(T1)、6‰(T2)3個鹽脅迫濃度,每個濃度重復5次。在處理前后的培養過程中每天用pH計測定營養液pH值,隔天用 1 mol/L H3PO4或 1 mol/L KOH調節pH,保持營養液pH為5.8;保持24 h連續通氣,每8 d換1次營養液。分別于處理后的第 10、20、30、40 和 50天上午 7:00—8:00采取每株葉位相近的當年生葉片,用封口袋裝并做好標記后置于冰盒中迅速帶回實驗室,用液氮冷凍后于-80℃冰箱保存,用于測定其各項生理和生化指標。
1.3 測定方法
1.3.1 生長指標測定
分別在試驗處理第10、20、30、40、50天時用鋼直尺(精度0.1 mm)測量各處理東方杉的固定枝長長度,按公式(1)計算東方杉固定枝條相對生長長度,即生長增量。
相對生長長度=試驗測量時長度-試驗處理前長度(1)
1.3.2 生理指標的測定
分別于試驗處理第10、20、30、40、50天采集0.2 g脅迫后的東方杉葉片,采用茚三酮比色法[6]用UV-2550分光光度計(島津公司,日本)測定葉片內的脯氨酸含量;根據Armon[7]的方法,于663 nm和645 nm測定葉綠素含量;采用NBT比色法[8]測定超氧化物歧化酶(SOD)活性;參照Buege等[9]的方法用硫代巴比妥酸法測定丙二醛(MDA)含量;采用電導法[10]測定相對電導率(REC)。
1.4 分析方法
所有數據采用SPSS 16.0進行統計分析,用Duncan’s新復極差法進行多重比較。
2 結果與分析
2.1 鹽迫條件下植物存苗率及葉色變化
鹽脅迫對東方杉存苗率和葉形態的影響結果見表1。
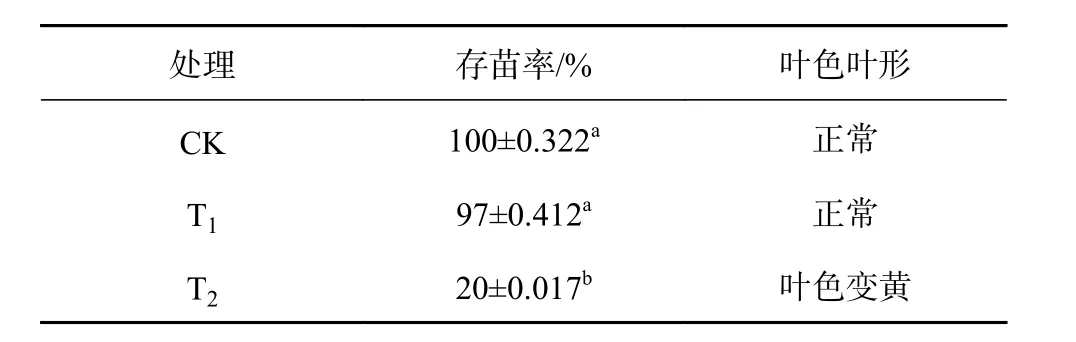
表1 鹽脅迫對東方杉存苗率和葉形態的影響Table 1 Effects of salt cooperative force on the rate and form of T. mucronatum×C. fortunei seedlings
由表1可知,鹽脅迫50 d后,不同鹽脅迫存苗率的處理間差異達顯著水平(P<0.05),T1的存苗率與CK無顯著差異,T2的存苗率顯著地低于CK與T1(P<0.05)。東方杉在T1脅迫下處理50 d后的葉色葉形表現生長,與CK無顯著差異;在T2脅迫下處理50 d后,葉色出現變黃的現象。
2.2 鹽脅迫條件下植東方杉固定枝條相對生長長度變化
由圖1可知,不同鹽濃度處理條件下的東方杉生長速度不同,在鹽脅迫處理的前期,T1枝條的生長未受到顯著抑制,而在脅迫理至50 d時,相比CK,T1枝條的生長出現緩慢減弱的現象。而T2枝條從第20天開始呈現出顯著地生長抑制現象(P<0.05);脅迫至第50天時,T2脅迫下東方杉枝條生長是CK的18.1%。
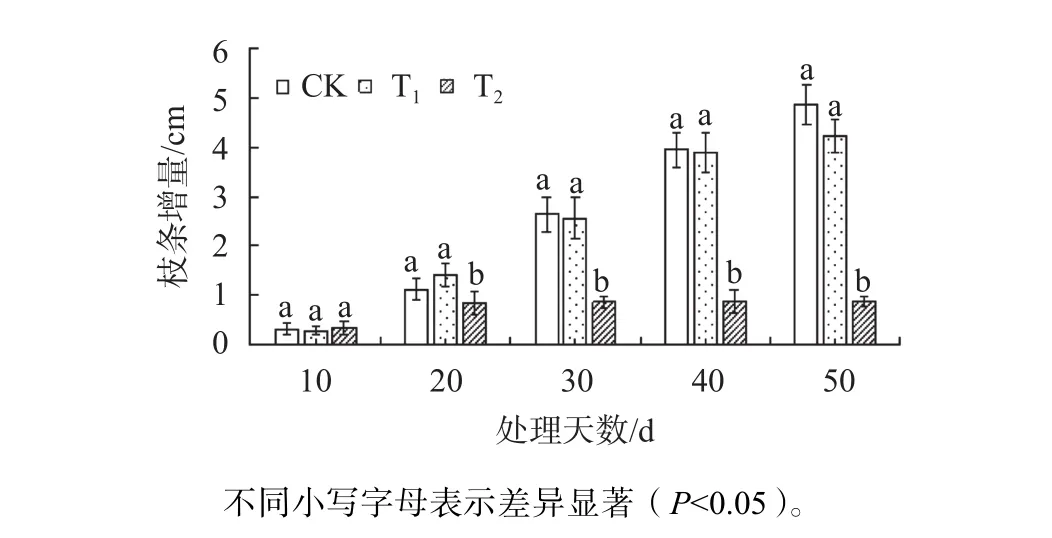
圖1 不同鹽脅迫下東方杉的相對生長長度變化Fig. 1 Changes of relative growth length of T. mucronatum×C. fortunei under different salt stress conditions
2.3 鹽脅迫條件下東方杉葉片中葉綠素含量變化
由圖2可知,在試驗脅迫的前40 d,T1新葉中的葉綠素含量與CK相比未出現顯著差異;在脅迫第50天時,T1葉片葉綠素含量顯著低于CK(P<0.05),是CK的88.9%。而在T2脅迫下,20 d后其葉綠素含量顯著低于CK和T1(P<0.05),隨著處理時間的增加;在脅迫至第50天時,T2新葉葉綠素含量是CK的37.6%。
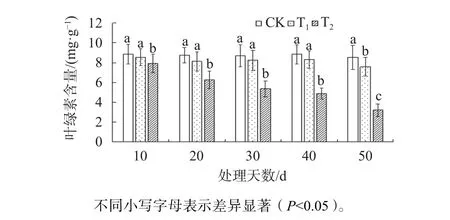
圖2 不同鹽脅迫下東方杉葉綠素含量變化Fig. 2 Changes of chlorophyll content of T. mucronatum×C. fortunei under different salt stress conditions
2.4 鹽脅迫條件下東方杉葉片MDA含量與REC的變化
由圖3可知,T2脅迫下MDA含量隨著處理時間的增加而增加,且顯著高于CK和T1(P<0.05),在脅迫至第50天時,分別是CK和T1的3.49和2.60倍。試驗40 d前,T1的MDA含量與CK無顯著差異;在第50天時,T1的MDA含量顯著高于 CK(P<0.05),是 CK的 1.34倍。在 T2下,東方杉REC值顯著高于CK與T1(P<0.05),在試驗至第50天時,分別是CK和T1的1.97和1.51倍。試驗40 d前,T1的REC與CK差異不顯著;在第50天時,T1的REC顯著高于CK(P<0.05),是CK的1.30倍。
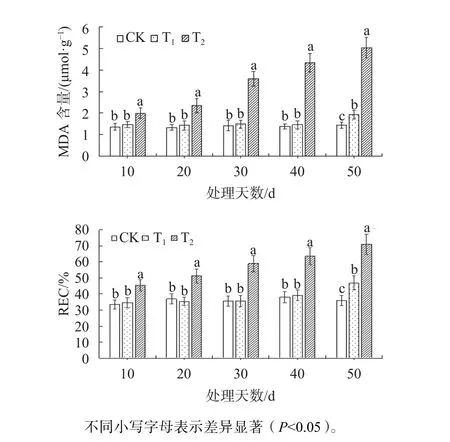
圖3 不同鹽脅迫下東方杉MDA含量與REC變化Fig. 3 Changes of MDA content and REC of T. mucronatum×C. fortunei under different salt stress conditions
2.5 鹽脅迫條件下東方杉葉片中脯氨酸含量的變化
如圖4可知,T1和T2的脯氨酸含量在第20天時顯著高于CK(P<0.05);在脅迫30 d后,T1脅迫的脯氨酸含量有所下降,但仍顯著高于CK(P<0.05),而T2的脯氨酸含量繼續增加,顯著高于CK與T1的脯氨酸含量(P<0.05),是CK的2.12倍;在脅迫50 d后,T1的脯氨酸含量仍顯著高于CK(P<0.05),T2的脯氨酸含量為CK的2.70倍。
2.6 鹽脅迫條件下東方杉葉片SOD活性的變化
由圖5可知,隨著處理時間的增加,T1和T2的SOD活性均顯著高于CK(P<0.05)。在脅迫至第40天時,T2的SOD活性達到最高值,分別是CK與T1的1.34與1.25倍;在脅迫至第50天時,T2的SOD活性下降。

圖4 不同鹽脅迫下東方杉脯氨酸含量變化Fig. 4 Changes of proline content of T. mucronatum×C. fortunei under different salt stress conditions
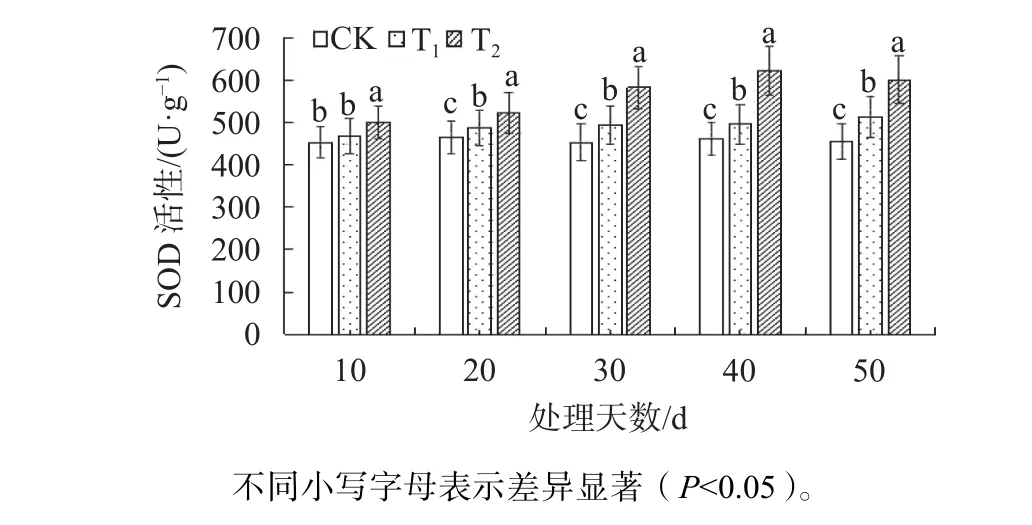
圖5 不同鹽脅迫條件下東方杉SOD活性變化Fig. 5 Changes of SOD activity of T. mucronatum×C. fortunei under different salt stress conditions
3 結論與討論
鹽脅迫是限制植物生長發育的主要環境因素之一。高鹽度對植物的毒害可以導致植物生長緩慢、產量降低甚至植株死亡[11]。在鹽脅迫條件下,植物體內的主要生理生化過程, 如水分吸收、光合作用、能量和物質代謝等均會受到不同程度的抑制。總體來說,鹽脅迫主要通過細胞膜滲透脅迫、離子毒害及營養失衡等對植物進行生長抑制。本研究發現,在T2鹽脅迫條件下,東方杉的枝條生長量減少,生長受到顯著抑制,而在較低的鹽濃度(T1)條件下,東方杉的枝條增量與對照組差異不顯著。這表明東方杉枝條在高濃度的鹽脅迫條件下,植物體為維持滲透壓的穩定,加劇了滲透調節物質的合成和積累,這就導致了植物體內的能量消耗增加,進一步限制了植物的生長[12]。
葉綠素是與植物光合作用相關的最重要的色素,鹽脅迫可能破壞植物葉片內的葉綠體功能結構,從而使葉綠素合成減少,含量降低下降,最終導致植株光合力減弱[13]。低濃度的鹽脅迫可以刺激植物體內的保護機制,促使葉綠素含量增加[14]。本研究發現,低濃度鹽脅迫處理初期,東方杉葉片內的葉綠素含量與CK差異不顯著,而隨著處理時間的增加,T1鹽脅迫條件下的東方杉葉片葉綠素含量略微下降;而T2鹽脅迫條件下的東方杉葉綠素含量一直顯著低于CK,表明鹽脅迫處理損害了東方杉的葉綠素合成路徑或合成器官,致使其葉片內的葉綠素合成受阻,葉綠素含量降低[15]。
細胞膜是鹽脅迫對植物造成傷害的原初位點,尤其是質膜,質膜在受到鹽脅迫傷害后會發生一系列的脅變,它的透性、滲透調節等功能都會因受到傷害而發生變化[16]。MDA是植物體內膜脂過氧化的最終產物,也是膜系統受傷害程度的重要標志之一,REC是反映細胞膜滲透率的重要指標。本研究發現,在低濃度的鹽脅迫條件下,東方杉體內的MDA含量和REC與CK差異不顯著,這表明東方杉作為耐鹽性較強的植物之一,在較低的鹽脅迫條件下,其細胞膜并未受到明顯傷害,但鹽脅迫時間的延長,細胞膜亦出現了輕微損傷。而T2對東方杉植株產生了顯著傷害,其MDA含量和REC相比CK顯著增加。這與錢瓊秋等[17]的研究相似,表明不同濃度的鹽脅迫和不同的處理時間可對植物細胞膜造成不同程度的傷害。
許多研究表明,植物在鹽脅迫下其滲透保護物質含量會增加,如茉莉酸、脯氨酸等[18]。本研究發現,東方杉在鹽處理下葉片中脯氨酸含量顯著高于CK,表明在東方杉體內脯氨酸可作為滲透調節物及膜和酶的保護物質對其起保護作用。在逆境脅迫條件下,SOD作為植物抗氧化保護酶系統重要的組成成分之一。它的變化可以反映細胞清除活性氧的能力,它可以能直接清除H2O2和O2.-,減少活性氧積累對植物造成的傷害[19],本研究發現鹽脅迫處理條件下的東方杉其SOD活性顯著高于CK。這表明鹽脅迫導致了東方杉體內活性氧含量的增加,而東方杉啟用自身保護機制通過增強SOD活性來清除活性氧,維持體內活性氧的動態平衡。本研究對東方杉在鹽脅迫下枝條增量、生理生化等分析比較,發現東方杉在3‰的鹽度生境下均能較好生長,只隨著處理時間的增加而出現生長略微受到抑制的現象,而在6‰的鹽度下其生長明顯受到抑制,直到植株死亡。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
