中國大鯢熱激蛋白基因Hsp90的克隆及極端溫度脅迫下的表達研究
2019-03-22 12:56:50李國勇許抗抗楊文佳駱建林
江蘇農(nóng)業(yè)科學(xué) 2019年24期
李國勇,許抗抗,楊文佳,駱建林,曹 宇,李 燦
(貴陽學(xué)院大學(xué)科技園/貴州省山地珍稀動物與經(jīng)濟昆蟲重點實驗室,貴州貴陽 550005)
熱激蛋白(heat shock protein,簡稱Hsp)是細胞或生物體在受到外界環(huán)境的刺激(例如高溫、干旱、饑餓、病原物侵入、重金屬離子等)誘導(dǎo)后產(chǎn)生的一類應(yīng)激性蛋白,它廣泛存在于動物、植物和微生物體內(nèi)[1-2]。Hsp可以促進蛋白質(zhì)的正確折疊和組裝,阻止變性蛋白聚體,從而將變性蛋白降解或重折疊,在維持生物體正常的生理代謝活動中起著重要作用。根據(jù)分子量大小、同源性和分子能可將熱激蛋白分為6個家族,即 Hsp10、Hsp90、Hsp70、Hsp60、Hsp40和 分 子 量 小Hsp等[3]。
Hsp90蛋白的分子量約為82~90 ku,它是生物體內(nèi)最豐富的蛋白質(zhì)之一,在正常情況下約占胞質(zhì)蛋白的1%~2%。Hsp90家族高度保守,其蛋白質(zhì)包含3個結(jié)構(gòu)域:N-端保守ATP(腺嘌呤核苷三磷酸)結(jié)合結(jié)構(gòu)域、中間結(jié)構(gòu)域和C-端多肽結(jié)合結(jié)構(gòu)域。Hsp90具有多種生物學(xué)功能,它不僅參與熱害、干旱、高鹽等脅迫響應(yīng),還可以參與組裝、穩(wěn)定和激活轉(zhuǎn)錄因子、激素受體以及蛋白激酶等關(guān)鍵信號蛋白[4]。目前,關(guān)于HSP90基因的研究多集中于昆蟲,已有多種昆蟲的Hsp90基因被克隆鑒定。研究發(fā)現(xiàn),這些基因在應(yīng)對極端溫度、重金屬離子以及殺蟲劑等各類非生物逆境脅迫中發(fā)揮著關(guān)鍵作用[5-11]。
中國大鯢(Andrias davidianus)俗稱娃娃魚,屬于兩棲綱(Amphibia)有尾目(Caudata)隱鰓鯢科(Cryptobranchidae)大鯢屬(Andrias),是現(xiàn)存?zhèn)€體最大的兩棲動物,為中國珍稀、瀕危物種,是國家二級保護動物,已被列入瀕危野生動植物種國際貿(mào)易公約(簡稱CITES公約)附錄Ⅰ中[12-15]。由于大鯢具有極高的營養(yǎng)價值、經(jīng)濟價值和科研價值,其養(yǎng)殖在中國華中、西南及西北等地區(qū)迅速發(fā)展,并逐步形成了適度規(guī)模化的地方優(yōu)勢特色。但是受到近幾年來全球氣候變暖的影響,極端天氣也頻繁出現(xiàn),對大鯢免疫系統(tǒng)的正常運行產(chǎn)生了嚴重影響,從而導(dǎo)致大鯢疾病的頻發(fā)。
本研究以高溫、低溫2種極端溫度脅迫下的大鯢為研究對象,分別克隆獲得CgHsp901、CgHsp902這個2個基因的全長序列,分析不同組織中這2個基因的表達變化,試圖了解該基因在大鯢應(yīng)對極端溫度脅迫中所發(fā)揮的作用,為深入開展大鯢繁殖與疾病預(yù)防研究奠定基礎(chǔ)。
1 材料與方法
1.1 材料準備與高溫處理
大鯢幼體來源于貴州省大鯢養(yǎng)殖基地,挑選平均體質(zhì)量為250~300 g的幼體大鯢,用長25 cm、寬10 cm、高13 cm的塑料盆飼養(yǎng)在溫度分別為0、5、15、20、25℃的人工培養(yǎng)箱中,每個溫度梯度飼養(yǎng)1盆共4尾幼體大鯢,飼養(yǎng)時間分別為24、48 h。為了模擬大鯢的洞穴生存環(huán)境,培養(yǎng)箱內(nèi)不開光照燈,試驗重復(fù)3遍。處理完成后,統(tǒng)計各溫度下大鯢幼體的存活率,對大鯢的心臟、肝臟、胃、脾、肌肉、皮膚和腸道7個不同組織進行取樣。樣品取出后馬上放入液氮中凍存,然后置于-81℃冰箱中保存?zhèn)溆谩?/p>
1.2 主要試劑
TRIzol試劑(TRIzol Regent,Invitrogen公司);反轉(zhuǎn)錄試劑盒(PrimeScript RT Reagent Kit,TaKaRa公 司);Taq酶(TaKaRa公司);膠回收試劑盒(Gel Extraction Kit,Omega公司);載體(pCEM-T EasyVector,Promega公司);感受態(tài)細胞DH5α(北京全式金公司);qPCR試劑(Go Taq qPCR Master Mix,Promega公司)。
1.3 試驗方法
1.3.1 大鯢幼體組織總RNA的提取及第一鏈cDNA的合成根據(jù)TRIzol試劑盒說明書提取大鯢幼體組織的總RNA,用瓊脂糖凝膠電泳和NanoDrop 2000核酸蛋白濃度測定儀檢測總RNA的完整性和濃度。取1μg總RNA,按照反轉(zhuǎn)錄試劑盒的操作說明合成第一鏈cDNA,稀釋2倍后保存于-20℃冰箱備用。
1.3.2 Hsp90基因全長cDNA的克隆 根據(jù)貴州省山地珍稀動物與經(jīng)濟昆蟲省級重點實驗室前期測定的大鯢轉(zhuǎn)錄組數(shù)據(jù),利用NCBI(美國國立生物技術(shù)信息中心)站點的ORF(開放閱讀框)Finder軟件(http://www.ncbi.nlm.hih.gov/gorf/or-fig.cgi)對所獲得的Hsp90基因進行分析,發(fā)現(xiàn)1個具有完整開放閱讀框的cDNA序列,用Primer Premier 5.0軟件設(shè)計特異性引物(表1)進行PCR擴增。RT-PCR反應(yīng)體系如下:2.5μL 10×PCR緩沖液,2.5μLMgCl2,2μL dNTPs,各1μL上下游引物,1.0μL cDNA,0.25μL Taq酶以及15μL ddH2O。

表1 引物相關(guān)信息
擴增條件如下:95℃預(yù)變性5 min;95℃變性30 s,56℃退火30 s,72℃延伸2.5 min,35個循環(huán);72℃延伸10 min。PCR產(chǎn)物經(jīng)1.5% 瓊脂糖凝膠電泳檢測后,利用Gel Extraction Mini Kit回收目的條帶并連接至pGEM-T Easy載體上,再將其轉(zhuǎn)化到DH5α大腸桿菌感受態(tài)細胞中,經(jīng)藍白斑篩選和PCR鑒定后進行測序驗證。本試驗中所用引物和DNA測序由成都擎科生物技術(shù)有限公司完成。
1.3.3 序列分析 利用DNAMAN 6.0(Lynnon Biosoft)對測序結(jié)果進行編輯和分析,并采用BLAST(http://www.ncbi.nlm.gov/BLAST/)工具進行同源性比對分析。利用在線數(shù)據(jù)庫Prosite(http://prosite.expasy.org/)分析保守區(qū)域。利用ProtParam(http://web.expasy.org/)和NetNGlyc 1.0 server(http://www.cbs.dtu.dk/services/NetNGlyc/)推測該氨基酸序列的理化性質(zhì)和N-糖基化位點。
1.3.4 Hsp90基因在溫度脅迫下的相對表達量 采用實時定量PCR技術(shù)檢測不同溫度脅迫后幼體大鯢Hsp90基因的相對表達量。根據(jù)“1.1”節(jié)的方法,用不同溫度處理大鯢,分別取7個組織,提取總RNA并反轉(zhuǎn)錄合成cDNA用于實時定量PCR,每個處理設(shè)4次重復(fù)。使用在線軟件Primer 3.0(http://frodo.wi.mit.edu/)設(shè)計定量PCR引物,內(nèi)參基因選取EF1α基因,引物序列信息見表1。PCR反應(yīng)體系如下:10μLGo Taq qPCR Master Mix、1μL cDNA、7μL nucleasefreewater(去核酸酶的水)和各1μL上下游引物,混勻,輕微離心,放入CFX96TM Real-Time System實時定量PCR儀(Bio-Rad)中進行擴增,反應(yīng)條件如下:95℃預(yù)變性2 min;95℃變性15 s,60℃退火30 s,72℃延伸30 s,共40個循環(huán);于60~95℃進行熔解曲線分析以保證反應(yīng)的特異性。用2-ΔΔCT方法計算目的基因的相對表達量[16]。
1.4 數(shù)據(jù)分析
采用SPSS 17.0軟件進行數(shù)據(jù)分析,應(yīng)用單因素方差分析(ANOVA)對不同溫度脅迫后大鯢幼體的存活率及其體內(nèi)Hsp90基因的相對表達量進行差異顯著性分析。
2 結(jié)果與分析
2.1 中國大鯢CgHsp901、CgHsp902基因的cDNA克隆及序列分析
采用RT-PCR技術(shù),從中國大鯢體內(nèi)克隆了熱激蛋白90基因,命名為CgHsp901、CgHsp902。CgHsp901基因cDNA全長為2 619 bp,開放閱讀框為2 388 bp,編碼795個氨基酸;CgHsp902基因cDNA全長3 024 bp,開放閱讀框為2 184 bp,編碼727個氨基酸。2個基因都具有Hsp90家族特有的標簽序列,與其他兩棲動物等物種的Hsp90有較高的同源性。2個基因的理論分子量都為90 ku,等電點分別為4.81、5.32。由CgHsp901推導(dǎo)的氨基酸序列包含Hsp90家族的4個簽名序列:第95~124位的NKEIFLRELISNSSDALDKIR、第161~169位的LGTIAKSGT、第403~412位的IKLYVRRVFI、第429~433位的GVVDSDDLPLNISRE(圖1)。由CgHsp902推導(dǎo)的氨基酸序列包含Hsp90家族的5個簽名序列:第39~59位 的 NKEIFLRELISNSSDALDKIR、第 106~114 位 的LGTIAKSGT、第129~145位的MIGQFGVGFYSAYLVAE、第336~345 位 的 IKLYVRRVFI、第 362~376 位 的GVVDSEDLPLNISRE,以及氨基酸C-端末尾基序MEEVD(圖2)。利用NetNGlyc 1.0 server進行分析可知,CgHsp901存在5個N-糖基化位點,分別為N143、N253、N477、N543和N548;而CgHsp902存在4個N-糖基化位點,分別為N150、N386、N492和N717。
2.2 2個基因在不同溫度脅迫下的應(yīng)激表達

利用實時熒光定量PCR(RT-qPCR)檢測這2個基因在中國大鯢心臟、肝臟、胃、胰腺、肌肉、皮膚和腸道7個不同組織中低溫(0、5℃)和高溫(20、25℃)脅迫下mRNA表達量的差異,結(jié)果顯示,大鯢幼體經(jīng)低溫處理24、48 h后,CgHsp901 mRNA的相對表達量在腸道、胰腺、胃和肝臟中不高,甚至在胰腺和肝臟中的部分表達量顯著下調(diào)(圖3、圖4)。但在皮膚、肌肉和心臟組織中,CgHsp901 mRNA的相對表達量整體上顯著升高。0℃處理24 h后,CgHsp901 mRNA在皮膚中的相對表達量是同溫度下腸道、胰腺中表達量的400倍,是肝臟組織中表達量的40倍;肌肉、心臟組織中CgHsp901 mRNA的相對表達量同樣升高,但是只有對照組的1~2倍。當處理溫度升高到5℃后,CgHspP901 mRNA在皮膚、肌肉和心臟組織中的表達量較0℃處理時顯著下降。
另外,當大鯢幼體處于20、25℃高溫脅迫時,該基因在大部分組織中的mRNA相對表達量都上調(diào)。其中在25℃高溫處理后,其在肌肉、胃、皮膚、心臟和肝臟中的表達量顯著上調(diào)。另外,對圖3、圖4比較可以看出,隨著處理時間的延長,CgHsp901基因的mRNA相對表達量沒有明顯升高,說明處理時間長短不影響CgHsp901基因的表達。以上結(jié)果說明,高溫、低溫脅迫能夠顯著誘導(dǎo)CgHsp901基因在大鯢體內(nèi)表達,且mRNA表達量與脅迫程度成正比。

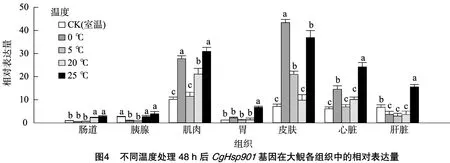
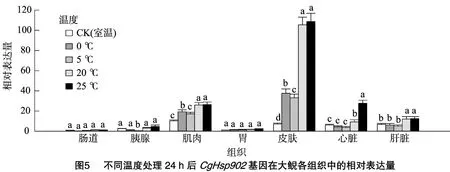
本研究對CgHsp902基因同樣進行了低溫(0、5℃)和高溫(20、25℃)脅迫處理,結(jié)果顯示,CgHsp902基因在腸道、胰腺和胃組織中的相對表達量明顯低于其在肌肉、皮膚、心臟和肝臟組織中的表達量。與CgHsp901基因一樣,CgHsp902基因在低溫、高溫脅迫下在皮膚、肌肉、心臟和肝臟中的mRNA相對表達量整體表現(xiàn)為顯著升高,而且表達量比CgHsp901基因高(圖5、圖6)。這同樣說明,高溫、低溫脅迫能夠顯著誘導(dǎo)CgHsp901基因在大鯢體內(nèi)的表達,且mRNA表達量與脅迫程度成正比。
3 討論
大鯢喜棲息于山澗水深、水質(zhì)清涼、泥潭水流急湍及巖石空洞較多的山溪中,對溫度變化非常敏感,棲息地氣候溫涼濕潤,年平均氣溫為12~17℃,最冷月平均氣溫2℃以上,最熱月平均氣溫在27℃以下,低溫和高溫暴露都可能造成其感染病菌,甚至死亡,從而影響種群發(fā)展[12]。本試驗發(fā)現(xiàn),在0、5℃的低溫及20、25℃高溫下分別處理24、48 h后,并未發(fā)現(xiàn)大鯢死亡。在溫度脅迫下大鯢只是將胃中內(nèi)容物吐出,低溫時不吃不動,高溫時則到處游蕩。從這個結(jié)果可以看出,大鯢對短時低溫或高溫環(huán)境有較強的適應(yīng)性。

熱激蛋白可以分為組成型熱激蛋白和誘導(dǎo)型熱激蛋白。組成型熱激蛋白在正常生理條件下的細胞中就大量存在,高溫等刺激不會誘導(dǎo)其大量表達,大多與維持細胞基本生理功能和形態(tài)建成密切相關(guān),而誘導(dǎo)型熱激蛋白在正常生理條件下不存在或表達量低,但在高溫等逆境刺激下能被強烈誘導(dǎo)表達,具有保護細胞的功能。熱激蛋白90在海洋魚類適應(yīng)海水溫度變化以及體內(nèi)外滲透壓變化等不利條件方面起著重要的作用。熱激蛋白不僅是熱脅迫時大量表達的蛋白,也是其他環(huán)境脅迫時大量表達的蛋白,如低溫、干旱、高滲透壓及重金屬等環(huán)境因素都能促使生物體表達熱激蛋白。在很多昆蟲種類中,Hsp90得到了大量的研究。然而有關(guān)大鯢熱激蛋白方面的研究很少,關(guān)于大鯢Hsp90的熱激蛋白基因家族中Hsp90B1、Hsp90AA1基因的研究幾乎未見報道。本試驗利用RT-PCR技術(shù)克隆了大鯢的Hsp90 cDNA片段,為進一步明確低溫和高溫脅迫與Hsp90基因表達量之間的相互關(guān)系奠定了基礎(chǔ)。本試驗克隆出的CgHsp901、CgHsp902在正常條件下檢測到少量存在于肌肉、皮膚、心臟和肝臟中,而當大鯢在0℃或25℃下脅迫24、48 h后,2個基因在皮膚中的表達量與對照組相比分別上升了5、20倍,說明這2個基因誘導(dǎo)表達的都是誘導(dǎo)型熱激蛋白。而大鯢在低溫、高溫的長時間脅迫下能夠全部存活,很可能是皮膚中CgHsp901、CgHsp902基因上調(diào),在抵抗低溫和高溫脅迫過程中發(fā)揮了重要作用,也可以說明在遇到溫度脅迫時皮膚是抵御其影響的第一道屏障。另外肌肉、心臟和肝臟組織在低溫和高溫脅迫下CgHsp901、CgHsp902基因的相對表達量也高于對照組,并且它們的平均值都顯著高于腸道、胃和胰腺組織中的相對表達量。這說明前三者在應(yīng)對溫度脅迫的時候也發(fā)揮了一定的作用,而后面3個組織則不起作用。
