豚草和三裂葉豚草不同植株部位種子萌發與入侵擴散關系
2019-03-16 02:54:42董合干王寒月王瑞麗馬倩倩趙文軒李秋霞
生態學報 2019年24期
關鍵詞:差異
劉 延,董合干,2,劉 彤,*,王寒月,王瑞麗,馬倩倩,趙文軒,李秋霞
1 石河子大學生命科學學院,石河子 832003 2 伊犁州農村能源環境工作站,伊寧 835000
生物入侵已經成為世界各國面臨的突出生態和經濟問題[1-4]。豚草(AmbrosiaartemisiifoliaLinn.)和三裂葉豚草(A.trifidaLinn.)現已被列入《全國農業植物檢疫性有害生物名單》和《中華人民共和國進境植物檢疫性有害生物名錄》,是世界上公認的有害植物[5]。目前,這兩種入侵植物廣泛分布于我國東北、湖南、北京和新疆等地區[6-8],嚴重危害生物多樣性,破壞生態系統,已給入侵地的農業和畜牧業生產造成巨大經濟損失。另外,其生長季釋放的花粉嚴重威脅人類健康[9-10],影響旅游業發展。因此,制定有效防控技術,一直是遏制這兩種入侵物種進一步擴散的現實問題和理論問題。
種子或果實在植物上的著生位置可影響種子質量、形態、休眠和萌發、擴散和幼苗建立[11]。黃帚橐吾(Ligulariavirgaurea)不同植株部位生產的種子的萌發和幼苗生長對策不同[12]。Moravcova等研究發現Heracleummantegazzianum不同植株部位生產的種子的萌發特性決定該物種入侵棲息地的能力[13]。Volis研究發現Triticumdicoccoide小穗上、下部分的種子在大小和萌發率上存在差異[14]。Koodziejek發現Peucedanumoreoselinum不同植株位置的種子(花序邊緣和中央)的質量和萌發率等均不同[15]。植物可通過調控不同植株部位種子的擴散和休眠能力,將種群密度調控在引起自疏的密度之下[16]。作者在野外觀測時發現,單位面積豚草密度可達到76—179株/m2,三裂葉豚草47—64株/m2;豚草平均單株結實2—3萬粒,三裂葉豚草單株可產種子0.3—0.5萬粒。這兩個物種每年生產數量如此巨大的種子,種群內如何調控自身密度,避免同胞競爭和高密度下死亡,依然保持非常強的種群入侵能力,這些問題值得深入研究。
目前,國內外對于豚草和三裂葉豚草的研究主要集中在對農田經濟作物的影響[17-18],防除[19-22],外界環境因子對種子萌發的影響[23-26],入侵機制(主要包括繁殖體壓力假說)[27],種子萌發、休眠特性[28-29]及擴散[30]等,然而,關于豚草和三裂葉豚草不同植株部位種子的萌發特性研究還未見報道。為此,本研究對豚草和三裂葉豚草不同植株部位種子開展了不同溫度條件下的萌發試驗,現將研究結果報道如下。
1 材料與方法
1.1 試驗材料
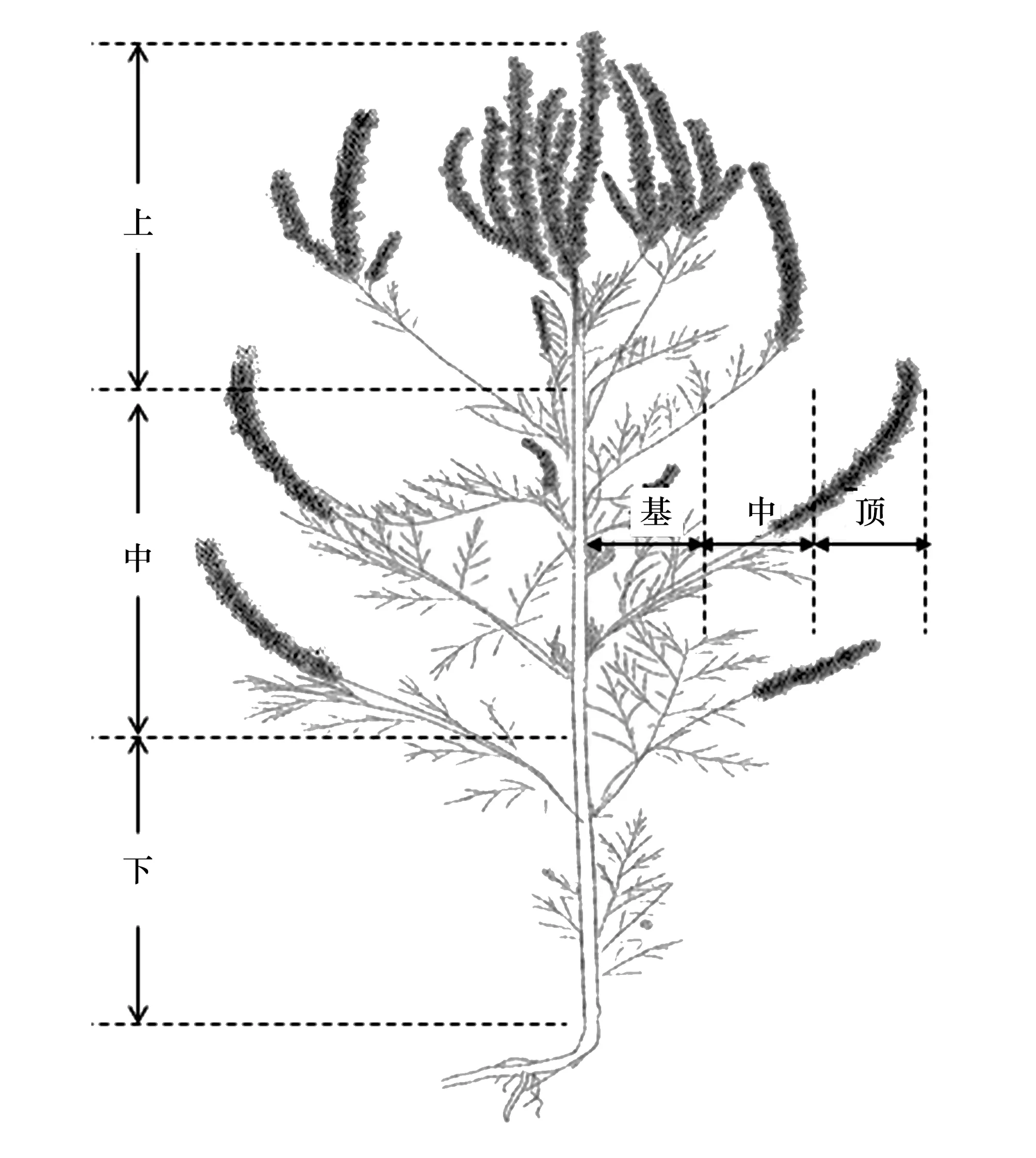
圖1 植株種子取樣部位示意圖 Fig.1 Diagrammatic sketch for sample position of seeds of Ambrosia artemisiifolia and A. trifida
在2016年10月赴新疆維吾爾自治區新源縣進行調查和采樣。在樣地隨機選取豚草和三裂葉豚草植株各30株。先根據植株高度,將植株平均劃分成三段,以相對于地面的高低定義為植株的上、中、下部;再根據不同的枝條長度,將枝條平均劃分成三段,以相對于植株主莖的遠近定義為頂、中、基部,將植株共分為9個部位(圖1)。
首先記錄單株豚草或三裂葉豚草不同植株部位所生產的種子數,并記錄相應的單株種子總數。如果種子散落,則根據相應部位的種子特征進行補充、核算;然后將30株豚草或三裂葉豚草相同植株部位生產的種子混合均勻,帶回實驗室進行試驗。
1.2 試驗方法
1.2.1不同植株部位種子的形態特征觀測
從豚草和三裂葉豚草植株各部位采集的種子中隨機抽取30粒,用游標卡尺測量種子的長度和寬度,重復3次,共測90粒。從每個物種各部位采集的種子中隨機選取100粒,用萬分之一天平進行稱重,重復3次,計算百粒重。
1.2.2不同植株部位種子的萌發試驗
將采自豚草和三裂葉豚草9個植株部位的種子在室溫下儲存6個月。從每個物種不同植株部位采集的種子樣品中隨機選取30粒,然后均勻地放置于鋪有兩層濕潤濾紙的直徑為9cm培養皿中。模擬豚草和三裂葉豚草集中分布區三月、四月、五月的月平均夜/日溫度(土壤溫濕儀,Watchdog1400全年觀測得出),分別測定種子在(5/15)℃、(10/20)℃和(15/25)℃三個溫度梯度(光/暗=12h/12h,光照強度3000lx)下的萌發率,萌發的種子以胚根突出種皮為準。每個處理重復3次,每日記錄萌發種子數量并且添加少量水分,以保證雙層濾紙濕潤。試驗持續60d,當連續5d沒有種子再萌發時,就視為發芽結束。
發芽率(%)=正常發芽種子粒數/種子總粒數×100
萌發開始時間(d)=從播種到第1粒種子萌發時所需要的時間
萌發持續時間(d)=開始萌發到萌發結束所需時間
1.3 數據分析
利用SPSS軟件統計分析本試驗所得數據,對豚草和三裂葉豚草不同植株部位種子的形態學指標進行單因素方差分析(One-way ANOVA);對不同植株部位、不同溫度下的種子萌發率、初始萌發時間、萌發持續時間進行多因素方差分析(Multi-way ANOVA);不同處理間差異顯著性用Duncan多重比較檢驗(Duncan′s multiple range test);利用origin9.0軟件作圖。
2 結果與分析
2.1 豚草和三裂葉豚草不同植株部位種子的基本形態特征
由表1可知,研究發現三裂葉豚草種子的平均長度為0.67cm,寬度0.4cm;豚草種子的平均長度為0.33cm,寬度0.18cm;三裂葉豚草種子大小是豚草種子的2—3倍。三裂葉豚草種子百粒重1.67g,豚草種子百粒重0.24g;三裂葉豚草種子百粒重是豚草種子的7倍。
由圖2可知,豚草、三裂葉豚草不同植株部位種子的平均長、寬以及百粒重變化不大,在平均值上下波動。

表1 豚草和三裂葉豚草種子形態指標對比
不同小寫字母表示差異顯著(P<0.05)

圖2 豚草和三裂葉豚草不同植株部位種子的形態指標對比Fig.2 The morphological characteristic of seeds produced at different positions of Ambrosia artemisiifolia and A. trifida不同小寫字母表示差異顯著(P<0.05)
2.2 豚草和三裂葉豚草不同植株部位種子數占植株種子總數的比例
由表2可以看出,豚草和三裂葉豚草不同植株部位生產的種子數量具有明顯差異。豚草和三裂葉豚草上頂、中頂部位生產的種子數占單株種子總數的比值最大;其次為上中、中中、下頂,最后為上基、中基、下中、下基。
總體來看,豚草、三裂葉豚草不同植株部位種子數占比,按照植株高度劃分,呈現出上端>中端>下端的趨勢;按照枝條位置劃分,呈現出頂端>枝條中部>枝條底部(近莖端)的趨勢。
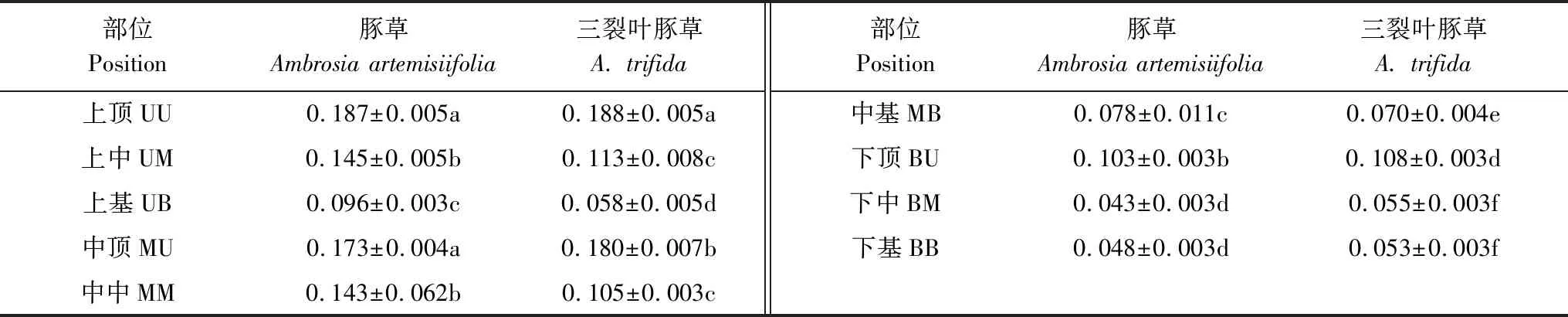
表2 豚草和三裂葉豚草不同植株部位產生種子數占單株種子總數比例/%
不同小寫字母表示差異顯著(P<0.05)
2.3 豚草和三裂葉豚草不同植株部位種子在不同溫度下的萌發特性
由表3可以看出,豚草不同植株部位的種子萌發率存在差異,但三裂葉豚草不同植株部位的種子萌發率沒有差異;溫度對豚草和三裂葉豚草的種子萌發具有顯著影響;而不同位置和不同溫度的交互作用對豚草和三裂葉豚草的種子萌發率沒有影響。

表3 溫度對豚草和三裂葉豚草不同植株部位種子萌發的影響
P<0.05,表示變異來源對萌發率差異顯著
由圖3和圖4可以看出,在供試溫度條件下,豚草和三裂葉豚草9個部位種子隨著溫度的升高,最終萌發率均呈逐漸下降趨勢;且各溫度梯度下,豚草種子的最終萌發率高于三裂葉豚草,在5/15℃溫度下,豚草種子萌發率為70%,三裂葉豚草種子萌發率為60%;在10/20℃溫度下,豚草種子萌發率為63%,三裂葉豚草種子萌發率為57%;在15/25℃溫度下,豚草種子萌發率為58%,三裂葉豚草種子萌發率為54%。

圖3 不同溫度(℃)下豚草不同植株部位種子萌發率/%Fig.3 Germination percentage of seeds produced at different positions of Ambrosia artemisiifolia at various temperatures/%不同小寫字母表示差異顯著(P<0.05)

圖4 不同溫度下(℃)三裂葉豚草不同植株部位種子萌發率/%Fig.4 Germination percentage of seeds produced at different positions of Ambrosia trifida at various temperatures不同小寫字母表示差異顯著(P<0.05)
豚草各部位種子在5/15℃溫度下的萌發率沒有明顯差異,萌發率均在70%左右。但隨著溫度的升高,各部位種子的萌發率逐漸表現出差異,植株的上端(尤其是上頂位置)的種子萌發率明顯高于其他部位,植株下端種子萌發率減小。在10/20℃溫度下,豚草上頂部位種子萌發率最高,達到70%,但中基以及下端種子萌發率只有60%;在15/25℃溫度下,豚草上頂部位種子萌發率為65%,中下及下端種子萌發率只有58%。
三裂葉豚草各植株部位種子在5/15℃、10/20℃溫度下的萌發率均沒有明顯差異,萌發率均在55%—60%之間。但在15/25℃溫度下,各植株部位萌發率表現出差異,且上部的萌發率高于中端和下端,上部萌發率達到60%,中部萌發率為55%,下部萌發率僅50%。
由圖5和圖6可以看出,三裂葉豚草種子初始萌發時間明顯長于豚草。隨著溫度的升高,豚草和三裂葉豚草各部位種子的初始萌發時間提前。且在三個溫度梯度下,兩種植物的初始萌發時間都呈現出下端>中端>上端的趨勢。

圖5 不同溫度下(℃)豚草不同植株部位種子初始萌發時間(d)Fig.5 Initial germination time of seeds produced at different positions of Ambrosia artemisiifolia at various temperatures(d)不同小寫字母表示差異顯著(P<0.05)

圖6 不同溫度下(℃)三裂葉豚草不同植株部位種子的初始萌發時間(d)Fig.6 Initial germination time of seeds produced at different positions of Ambrosia trifida at various temperatures(d)不同小寫字母表示差異顯著(P<0.05)
豚草在5/15℃溫度下,各部位種子初始萌發時間差異明顯,上頂、上中、上基、中頂、中中種子13d左右開始萌發,而中基和下端種子17d左右才開始萌發;在10/20℃溫度下,上頂部位種子萌發最快,其他植株部位的種子7d左右開始萌發;在15/25℃溫度下,上頂部位的種子5d就開始萌發,而且9個部位種子初始萌發時間的差異性逐漸消失。
三裂葉豚草在5/15℃溫度下,上頂部位種子需要45d左右才開始萌發,不同植株部位種子初始萌發時間最長相差6d;在10/20℃溫度下,各個植株部位的種子萌發提前,不同植株部位的種子在35d左右開始萌發;在15/25℃溫度下,各個部位的種子初始萌發時間在30d左右。
由圖7和圖8可以看出,豚草和三裂葉豚草種子隨著溫度的升高,持續萌發時間增加。在各溫度梯度下,豚草種子的持續萌發時間均高于三裂葉豚草。在三個溫度梯度下,二者的持續萌發時間,各部位差異變化明顯,均呈現出上端>中端>下端的趨勢。
豚草在5/15℃溫度下,不同植株部位種子的持續萌發時間為36—41d;在10/20℃溫度下,上中位置種子的持續萌發時間為44d,而下端種子持續萌發時間為38d;在15/25℃溫度下,中中、中基和下部種子萌發持續時間在47—49d左右,上頂、中頂種子萌發持續時間在42d左右。
三裂葉豚草在5/15℃溫度下,不同植株部位種子的持續時間最短僅為6d,最長僅為12d;在10/20℃溫度下,上中、中前部位種子的持續萌發時間為17—19d,而下端位置種子的持續萌發時間為13d;在15/25℃溫度下,上部、中部種子萌發持續時間19—23d,下部種子萌發持續時間在15—17d。

圖7 不同溫度下(℃)豚草不同植株部位種子持續萌發時間(d)Fig.7 Germination duration of seeds produced at different positions of Ambrosia artemisiifolia at various temperatures(d)不同小寫字母表示差異顯著(P<0.05)
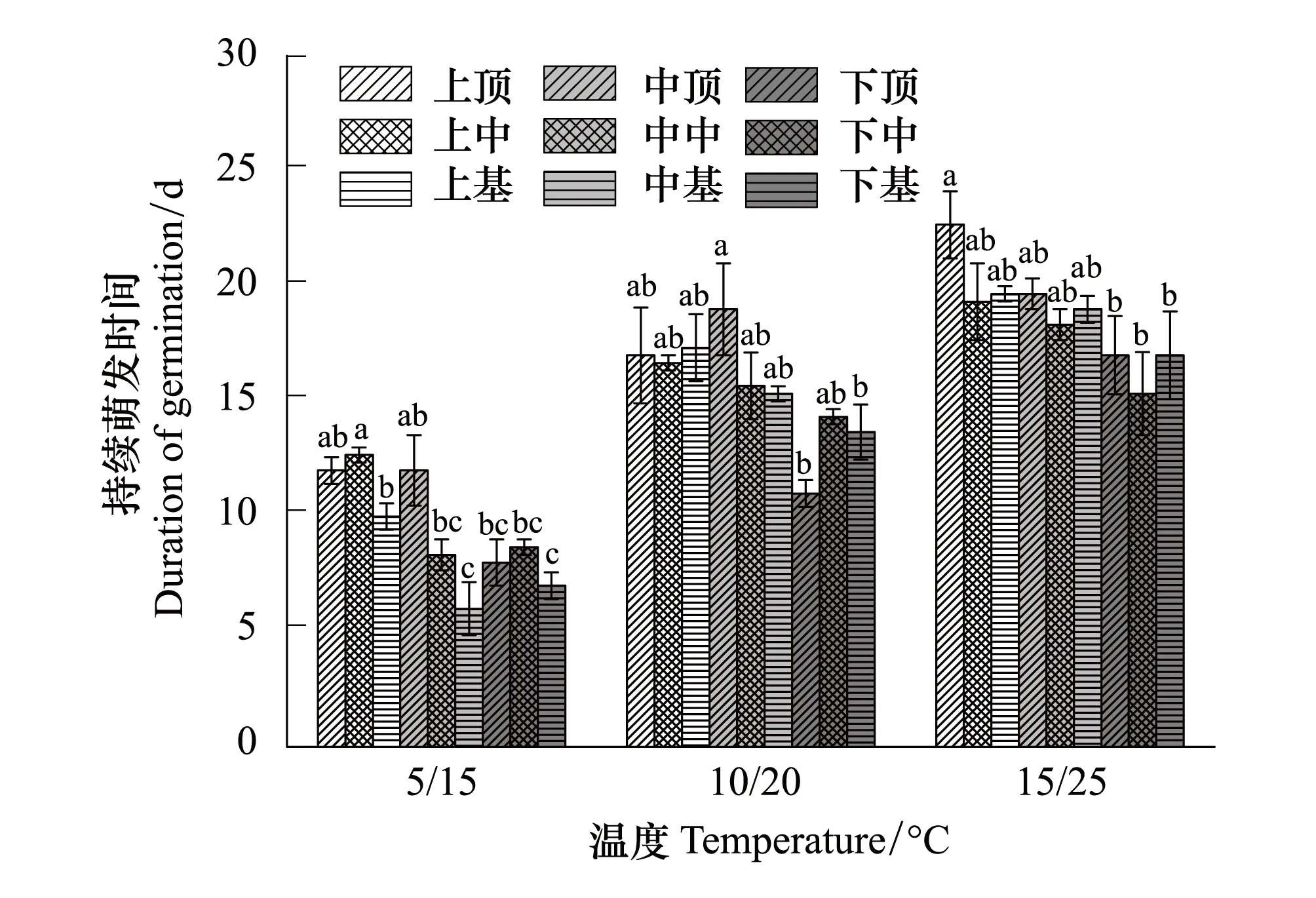
圖8 不同溫度下(℃)三裂葉豚草種子不同植株部位種子的持續萌發時間(d)Fig.8 Germination duration of seeds produced at different positions of Ambrosia trifida at various temperatures(d)不同小寫字母表示差異顯著(P<0.05)
3 討論
3.1 種子大小差異是兩個物種擴散、分布差異的主要原因
豚草和三裂葉豚草自2010年入侵到伊犁河谷,二者均主要分布在路邊、農田邊,草場、居民區等生境,至2016年二者的入侵面積分別是2010年的10150倍和2150倍;2016年三裂葉豚草單獨分布區面積比例僅為3.33%,但豚草單獨分布區面積比例高達79.52%[8]。由于伊犁河谷屬于典型的農牧交錯區,牛羊轉場、農事活動等人為干擾會導致兩個物種在伊犁河谷表現出更快的擴散速度及更大的擴散面積,這也是二者入侵伊犁河谷的主要方式。但豚草的入侵面積明顯高于三裂葉豚草。已有研究表明,小種子比大種子具有明顯的優勢,其擴散能力強,擴散距離遠[24,31-34],并且同樣的風速和高度下,較小的種子傳播得更遠[35]。豚草種子比三裂葉豚草種子小,質量輕,所以,更易于在入侵地擴散。
種子大小和萌發速率之間也存在著顯著的負相關性,即小種子比大種子萌發得快[36-37]。本試驗說明豚草低溫時僅需要13—18d就可以打破休眠而萌發,而三裂葉豚草則需要43—48d才能萌發。
3.2 不同植株部位種子數量比例和萌發特性是兩個物種調節種群密度的重要方式
植物是固著的自養型生物,必須在原地繁衍生息,既要承受來自其他生物的侵害,又要遭受鄰體對水分、礦質營養和光照的競爭。種子大量萌發勢必導致單位面積種群內對有限資源的爭奪,造成個體的生長量下降和死亡率升高,如此“對策”不利于植物的生存和發育,而休眠和擴散能夠允許種子從時間和空間上避免萌發的不利條件[38-41],緩解種群內部植株競爭。豚草和三裂葉豚草不僅植株高密度分布且結實量巨大,二者如何調節種群內部植株密度顯得極為關鍵。
果實或種子在成熟后的散布是決定植物幼苗定植和空間格局的一個重要過程[39]。種子擴散不但受種子大小影響,也與種子在母體植株的相對高度有關。種子位于母體植株突出的位置更容易利用風、動物等媒介進行遠距離傳播[26,42-43],有助于在空間上擴大后代的分布范圍,增加其對環境的適應能力[43-44]。我們的研究表明,豚草和三裂葉豚草9個部位種子雖然沒有大小、質量的明顯差異,但二者植株靠外部的位置(如上頂、上中、中頂、中中、下頂)每年的結實量占結實總量近73%,大批量種子有進行長、遠距離擴散的能力或潛力,以此避免大量種子就近擴散到母株周圍。
較早地出苗就意味著擁有較長的生長期和占有較高的優勢層次[45],增強與自身、其他植株種子的競爭能力。研究表明,豚草和三裂葉豚草植株各個部位種子的萌發時間不同,上部種子比中部、下部種子提前幾天開始萌發。而且上部種子位置高,種子擴散距離更遠,更易于拓殖到新生境,避免了同胞間的競爭,在種群更新中占有較為重要的地位。擴散能力弱的種子(如生長在植物較低節點位置的種子)最容易遭受同胞競爭,它們一般表現出較低的萌發率和較慢的萌發速度[46]。所以,不同植株部位種子的數量比例和萌發特性是兩個物種調節種內密度的重要方式。
3.3 不同植株部位種子大小、萌發特性和數量比例是豚草和三裂葉豚草形成入侵能力的必要條件
本研究設置伊犁河谷早春3月、4月、5月三個月份的夜/日均溫變化梯度,試驗結果表明,豚草和三裂葉豚草9個部位種子在三個試驗溫度梯度范圍內均能萌發,表明其能在4月、5月份持續萌發,與野外觀察一致,這也是面對外界環境最大化生存適合度的表現[47]。隨著溫度的升高,具有擴散優勢、生殖比例大的上頂、上中、中頂的種子擁有比其他部位(上基、中中、中基、下頂、下中、下基)更高的萌發率,說明二者的上頂、上中、中頂部位所產生的種子具較強的擴散能力和低休眠性,有利于豚草和三裂葉豚草快速占據新生境并擴大種群,代表“高風險”的適應策略。這兩個物種的上基、中中、中基、下頂、下中、下基部位的種子在母株周圍就近擴散,萌發率低、休眠時間長,形成長久性種子庫。一方面利用長久性種子庫中的種子繁殖后代,避免滅絕的危險[48]。對于有害雜草而言,為了避免被消滅,豚草種子在土壤中埋藏40年后仍能發芽[29],可見豚草適應性如此之強烈。另外,豚草和三裂葉豚草具有極強的種間競爭能力,可形成大片的高密度單一種群,造成“碾壓式”入侵。
此外,豚草和三裂葉豚草在植株外部(上頂、上中、中頂、中中、下頂)種子占據數量優勢,這種生殖策略是否與植株的花序位置效應有關[12,49]?而產生花序位置效應是由于占有優勢位置的花對資源有競爭優勢[50-51]還是由于花粉限制的原因[52]?不同植株部位生產的種子產生不同的萌發特性是由于母體效應[53]還是種子自我抑制[54]?另外,除了不同植株部位種子大小、萌發特性和數量比例之外,種子與土壤微生物的作用、化感作用等均可能影響二者入侵擴散,以上問題有深入研究必要,更有助于深入了解豚草和三裂葉豚草種子與入侵擴散的關系,可為提出高效生態防控建議提供理論基礎。
致謝:新源縣農業局楊春虎、王俊濤幫助野外調查,特此致謝。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
