lysC定點突變及lysC、asdA串聯表達對谷氨酸棒桿菌L-蘇氨酸積累的影響
2019-03-15 05:52:12黃勤勤王慧梅梁玲黃欽耿吳松剛黃建忠
生物技術通報 2019年2期
黃勤勤 王慧梅 梁玲 黃欽耿 吳松剛 黃建忠
(福建師范大學生命科學學院 福建師范大學工業微生物教育部工程研究中心,福州 350117)
L-蘇氨酸(L-Threonine)被認為是動物體內第二或第三限制性氨基酸,是人體和動物的必需氨基酸。已被廣泛應用于食品、飼料及醫藥行業[1-2]。目前,國內外生產L-蘇氨酸的主流方法是微生物直接發酵法[3-4]。其中,大腸桿菌(Escherichia coli)工程菌是目前生產中最為常用的宿主菌株[5]。但是由于其安全性不高,并且隨著代謝的進行,生產過程中會逐漸積累一些毒素,故在醫藥行業中不能使用大腸桿菌生產蘇氨酸。相對來說,無致病性的安全菌株谷氨酸棒狀桿菌同樣作為氨基酸的傳統生產菌株,其優勢就展現出來了。隨著谷氨酸棒桿菌模式菌株ATCC 13032測序的完成,氨基酸合成途徑及代謝調控機理逐漸清晰,以及谷氨酸棒桿菌遺傳操作體系的建立和完善,谷氨酸棒桿菌正成為氨基酸代謝工程改造的基盤菌株[6]。
L-蘇氨酸的生物合成是以TCA循環中間代謝物草酰乙酸為前體。底物葡萄糖代謝經過糖酵解途徑、磷酸戊糖途徑、TCA循環及C3-C4回補途徑等中心代謝途徑,由草酰乙酸經天冬氨酸轉氨酶催化生成L-天冬氨酸,即進入L-蘇氨酸合成途徑。L-天冬氨酸經天冬氨酸激酶(AK,編碼基因lysC)和天冬氨酸半醛脫氫酶(ASD,編碼基因asdA)兩步酶催化反應后,再經過高絲氨酸脫氫酶生成L-高絲氨酸,經高絲氨酸激酶生成L-磷酰-高絲氨酸,最后經蘇氨酸合酶形成L-蘇氨酸[7]。AK由LysC基因編碼,同時受L-蘇氨酸、L-賴氨酸、L-甲硫氨酸的協同反饋抑制。天冬氨酸激酶(AK,編碼基因lysC)是合成L-蘇氨酸中的關鍵酶之一,控制天冬氨酸族合成途徑總碳流量。因此,通過對一些經傳統育種方法選育得到的L-蘇氨酸生產菌進行組學研究分析,發現AK和ASD是該合成途徑上影響L-蘇氨酸產量的關鍵酶之一[8]。
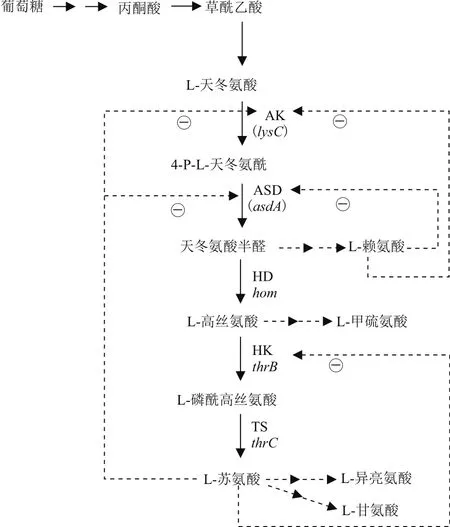
圖1 L-蘇氨酸的生物合成途徑
Dong等[9]將谷棒桿菌的lysC基因第279位突變點A突變為T解除了L-賴氨酸和L-蘇氨酸反饋抑制的能力。其表達lysC基因并且串聯表達hom基因和thrB基因使部分碳流量從L-賴氨酸(50%)轉移至L-蘇氨酸,使谷氨酸棒桿菌L-蘇氨酸產量達到4.85 g/L。徐德雨等[10]發現在高產菌的天冬氨酸激酶AK中存在 G359D 突變,驗證突變后可阻斷賴氨酸引起的別構效應,從而有效解除賴氨酸與蘇氨酸的協同抑制。通過體外天冬氨酸激酶野生型和G359D 突變體酶活檢測,其突變體在10 mmol/L賴氨酸和蘇氨酸同時存在時酶活達到76.94%±1.61%,而野生型酶活僅4.38%±1.28%。由此可見,AK酶定點突變可解除L-蘇氨酸、L-賴氨酸的協同抑制,提高酶活,從而使碳流量更多的流向產L-蘇氨酸方向,達到提高L-蘇氨酸產量目的。
本文以選育獲得的一株甲硫氨酸和異亮氨酸缺陷(Met-/Ile-)及 α-氨基 -β-羥基戊酸抗性(AHVr)的谷氨酸棒狀桿菌T11(Corynebacterium glutamicumT11)為出發菌株,通過基因克隆和序列分析,在已有突變的基礎上,采用PCR介導的方式對lysC基因進行定點突變,構建L-蘇氨酸高效抗反饋抑制的關鍵酶基因lysCr-asdA串聯表達組件轉化出發菌株T11,篩選獲得過量積累L-蘇氨酸工程菌株,進行搖瓶和30 L全自動發酵罐分析工程菌的產酸能力、細胞生長及雜酸的積累情況,為進一步的L-蘇氨酸的代謝工程改造奠定基礎。
1 材料與方法
1.1 材料
1.1.1 菌株和質粒 谷氨酸棒狀桿菌T11(C.glutamicumT11)本實驗室選育并保藏;大腸桿菌JM109感受態細胞,購自于全式金;穿梭表達載體pZ8-1由中國科學院微生物所李寅研究員贈送。
1.1.2 試劑和儀器 限制性內切酶、T4 DNA連接酶、DNA Marker、PrimeStar DNA聚合酶和PCR試劑:TaKaRa寶生物公司產品;引物合成、sanPreP 柱式膠回收DNA試劑盒、質粒小量提取試劑盒:生工生物工程(上海)股份有限公司產品;點突變試劑盒Fast Mutagenesis System:北京全式金生物技術有限公司;酵母粉和胰蛋白胨:Bio Basic Inc公司產品;其余試劑均為國產或進口分析純。
PCR儀:ABI產品;電泳儀:DYY-6C北京六一儀器廠;紫外分光光度計:UV-282PCS,尤尼柯(上海)儀器有限公司;電轉儀:Eppendorf 2510,艾本德中國有限公司;高效液相色譜:島津公司;30L發酵罐:鎮江東方生物工程設備有限公司。
1.1.3 培養基 LB 培養基(g/L):胰蛋白胨10,酵母提取物5,NaCl 10,若為固體培養基,則額外添加2.0%(w/v)的瓊脂粉。
LBG 培養基:LB培養基添加0.5%(w/v)的葡萄糖,若為固體培養基,則額外添加2.0%(w/v)的瓊脂粉。
上述培養基的pH均調至7.0±0.1大腸桿菌用LB培養基;C. glutamicum用LBG培養基,其中用于C.glutamicum和E. coli的篩選標記卡那霉素使用的終濃度分別為 25 μg/mL 和 50 μg/mL。
種子培養基各組成質量分數分別為:葡萄糖2.5%,豆粕提取粉2.0%,尿素0.125%,玉米漿2.0%,KH2PO40.1%,MgSO40.05%,FeSO4·7H2O 0.05%,VB1100 μg/L,VH150 μg/L,20% 的氨水調pH 6.9±0.1。
發酵培養基組成質量分數分別為:葡萄糖8%,豆粕提取粉2.0%,(NH4)2SO43.5%,玉米漿2.0%,KH2PO40.1%,MgSO40.1%,FeSO4·7H2O 0.05%,CaCO33.0%,VB1100 mg/L,VH150 mg/L,20%的氨水調pH 6.9±0.1。
1.2 方法
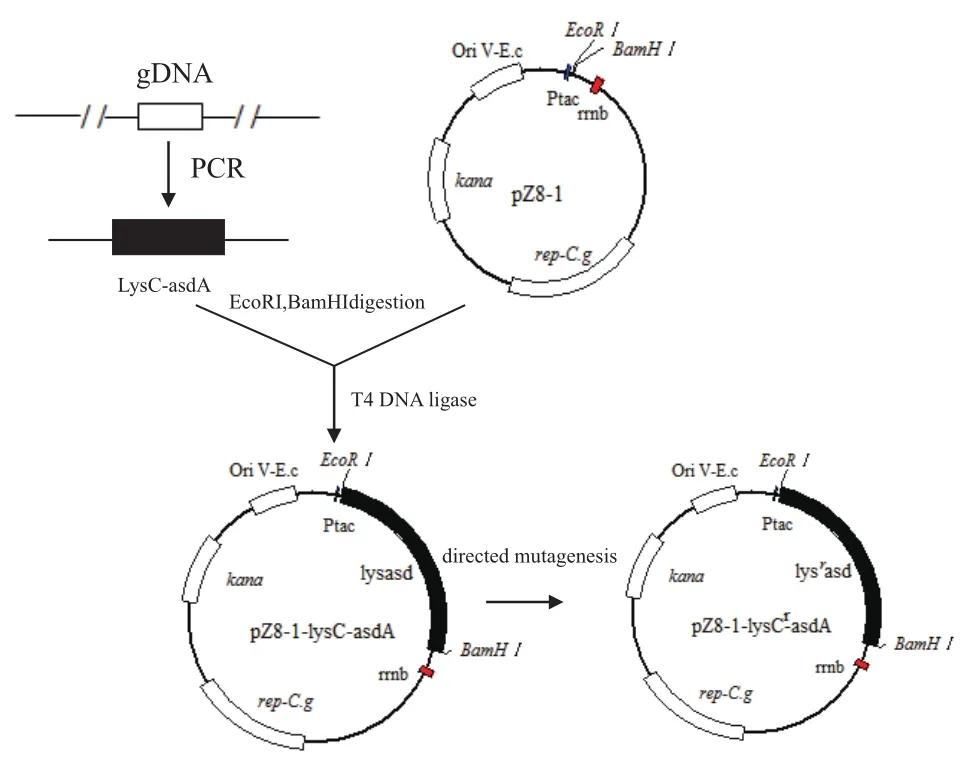
1.2.1lysC-asdA串聯基因簇的克隆 根據C. glutamicumT11菌株的16S rDNA的序列比對結果,以谷氨酸棒狀桿菌ATCC13869基因組序列為模板,設計lysC-asdA串聯基因簇的上下游引物CA-F、CA-R。
CA-F:5'-CGGAATTCCG GTGGCCCTGGTCGTACAGAAATATGGC-3';CA-R :5'-CGGGATCCCG TTACTTAACCAGCAGCTCAGCG-3'。其中引物CA-F帶有EcoRⅠ酶切位點和保護堿基,CA-R帶有BamHⅠ酶切位點和保護堿基(下劃線序列)。
以C. glutamicumT11菌株基因組為模板進行PCR擴增。反應體系為50 μL:10×Ex buffer 5 μL,引物CA-F、CA-R各2 μL,基因組DNA模板0.5 μL,dNTP 2.5 μL,Ex Taq 酶 1 μL,ddH2O 補 足 至50 μL。反應條件為:95℃預變性 5 min;95℃ 40 s,57℃ 40 s,72℃ 2.5 min,35個循環;72℃延伸5 min。PCR產物1%的瓊脂糖凝膠檢測,膠回收試劑盒純化回收,備用。
1.2.2 pZ8-1-lysC-asdA重組載體的構建 采用限制性內切酶EcoRⅠ和BamHⅠ分別對串聯基因片段lysC-asdA和穿梭表達質粒pZ8-1進行雙酶切處理,分別回收目的片段,然后用 T4 DNA ligase進行連接,連接體系參考TaKaRa ligase kit 說明書。連接反應結束后,轉化E. coliJM109感受態細胞,轉化液涂布含50 μg/mL卡那霉素的LB平板,37℃ 倒置培養過夜。從平板上挑選單克隆,采用驗證引物(F:5'-TTCTGGCAAATATTCTGAAATGAGC-3'和R:5'-TTCGCAACGTTCAAATCCGC-3')進行菌落PCR驗證,并挑取陽性克隆進行37℃過夜培養,提取質粒,并對質粒進行酶切和測序驗證。
1.2.3 PCR介導的lysC基因的定點突變 為了將lysC基因的第279位密碼子定點突變,以陽性克隆質粒pZ8-1-lysC-asdA為模板,使用攜帶突變位點的引物Cr-A-F和Cr-A-R,參考Fast Mutagenesis System說明書,進行PCR介導的定點突變,擴增,挑取抗性平板克隆進行酶切驗證,并將陽性克隆送交上海生工進行測序驗證。
lysC基因PCR介導的定點突變引物設計如下:Cr-A-F:5'-CCGATAAGCCAGGCGAGACTGCGAAGG TTTTCCG-3';Cr-A-R:5'-CAGCCTCGCCTGGCTTATC GGAAATACCCAGAACGG-3'
其中加粗的堿基為引入的突變堿基。提取質粒并進行測序驗證,構建的表達載體命名為pZ8-1-lysCr-asdA,具體的構建原理如圖2所示。
1.2.4 產L-蘇氨酸工程菌的構建和篩選
1.2.4.1 構建 將獲得的陽性pZ8-1-lysCr-asdA重組質粒電轉導入谷氨酸棒狀桿菌T11感受態細胞中[11]。谷氨酸棒狀桿菌T11感受態細胞按文獻制備[12],抗性平板篩選及菌落PCR驗證,構建含有表達質粒pZ8-1-lysCr-asdA的工程菌株。
1.2.4.2 篩選 從抗性平板中挑取菌落大的陽性克隆,直接接種至每孔含有1 mL發酵培養基的96微孔板初篩,30℃,220 r/min培養48 h,檢測發酵液中L-蘇氨酸含量,并對初篩L-蘇氨酸積累較多的菌株進行搖瓶發酵復篩和遺傳穩定性驗證。遺傳穩定性分析為每24 h斜面傳代一次,傳代5代,每隔一代進行搖瓶發酵,檢測工程菌株的L-蘇氨酸產量。最后選取L-蘇氨酸產量較高且遺傳穩定的工程菌株進行進一步的發酵分析。
1.2.5 L-蘇氨酸工程菌株的搖瓶發酵分析 所獲得的工程菌T11/pZ8-1-lysCr-asdA經搖瓶發酵后分析其L-蘇氨酸含量、菌體生物量以及其他氨基酸的生成情況。搖瓶發酵采用一級發酵,培養條件為:30℃,220 r/min發酵60 h,同時以出發菌株T11為對照菌株。氨基酸的含量采用高效液相色譜系統(HPLC)自動柱前衍生化法測定。色譜柱為Venusil-AA柱。采用流動相組成為水相(1 L):7.6 g無水乙酸鈉,925 mL 高純水,冰醋酸調pH至6.5;有機相(1 L):80% 乙腈。色譜條件:柱溫40℃,流量1.0 mL/min,SPD-M20A檢測器。
1.2.6 L-蘇氨酸工程菌株的30 L罐補料分批發酵 根據搖瓶發酵特性,結合已有的發酵工藝,采用二級發酵的方式對L-蘇氨酸工程菌株T11/pZ8-1-lysCr-asdA進行補料分批發酵,考察高密度發酵情形下的L-蘇氨酸產率、葡萄糖的利用率及細胞生長情況,綜合評價菌株的發酵潛力。
搖瓶種子培養:挑一環新鮮斜面上的菌體接種至裝有 30 mL LBG培養基的250 mL 三角瓶中,30℃,220 r/min 振蕩培養12 h至對數生長中后期,?OD562nm約為 0.2×100。
種子罐培養:按0.33%的接種量,將搖瓶種子液轉接至含有3 L種子培養基的5 L種子罐中,初始攪拌300 r/min,培養溫度30℃,種子罐周期8 h,移種?OD562nm約為0.30×100。
補料發酵培養:30 L發酵罐初始裝液量為15 L,移種量 20%,初始葡萄糖濃度約125 g/L,發酵溶氧維持在25%-35%,初始攪拌轉速 300 r/min,發酵溫度30℃,流加 25%的氨水以控制 pH在6.9-7.0,發酵期間每隔2 h 取樣檢測菌體生物量及殘糖,根據耗糖速率確定補料量;當殘糖降至 5.0 g/L 以下時連續流加700 g/L 的葡萄糖,維持殘糖濃度在5.0-10 g/L,發酵結束前4 h,停止流加葡萄糖,當殘糖基本耗盡時,發酵結束,整個發酵周期約60 h。
2 結果
2.1 lysC-asdA串聯基因的克隆
以谷氨酸棒狀桿菌生產菌T11基因組為模板,引物CA-F、CA-R為引物對擴增出一段約2 300 bp條帶,并對目的產物進行膠回收(圖3),其與相似基因組ATCC13869的lysC-asdA的理論大小(2 324 bp)基本一致。
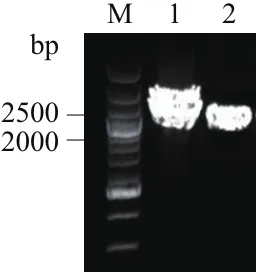
圖3 lysC-asdA基因PCR擴增及回收
2.2 pZ8-1-lysC-asdA重組載體的構建及lysC的定點突變
轉化經酶切的質粒與目的基因,長出單菌落后隨機挑取9個單克隆,用驗證引物直接進行菌落PCR驗證,隨機挑取的8個單克隆均為陽性克隆(圖4-A),篩選菌落PCR陽性的單克隆進行質粒抽提和單切和雙切驗證,確認質粒pZ8-1中插入大小約為2 400 bp的目的片段(圖4-B)。另外,測序結果顯示插入的基因片段確為目標片段,確認pZ8-1-lysC-asdA構建成功。
值得注意的是,克隆的lysC-asdA基因較親緣關系最近(相似性為99%)的ATCC13869存在顯著差異,其lysC基因的ORF區域有多達8個堿基的突變,其中6個為無義突變,但該突變在理論上都與密碼子識別相關,另外兩個堿基的突變都導致了氨基酸的突變;asdA基因也有4個堿基的突變,但都是無義突變,這4個堿基的突變都同樣涉及到密碼子優化的問題。這極有可能和出發菌株T11本身經歷過多輪次的誘變選育有關。pZ8-1-lysC-asdA質粒載體經EcoRⅠ和BamHⅠ酶切后,出現7 000 bp和2 400 bp的兩條帶,與預期結果一致。
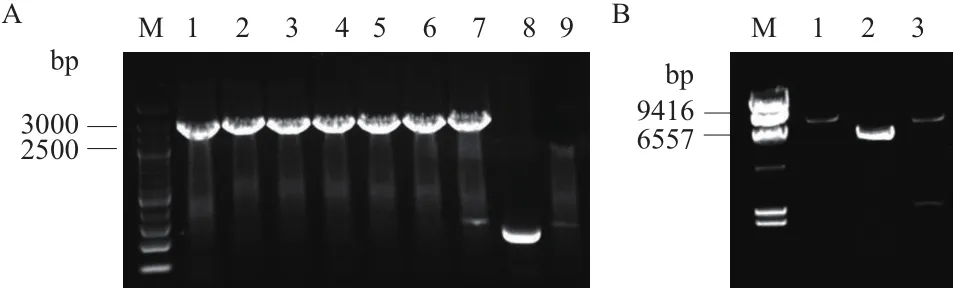
A. 重組表達質粒pZ8-1-lysC-asdA的菌落PCR驗證;B. pZ8-1-lysC-asdA質粒的酶切驗證;(A:M:200 bp Ladder DNA marke;1-7:重組表達質粒pZ8-1-lysC-asdA菌落PCR驗證;8:pZ8-1空質粒對照;9:T11出發菌株對照;B:M:λ-Hind Ⅲ digest DNA marker;1:pZ8-1-lysC-asdA質粒經BamHⅠ單酶切;2:pZ8-1質粒經BamHⅠ單酶切;3:pZ8-1-lysC-asdA質粒經EcoRⅠ和BamHⅠ雙酶切)
為進一步提高lysC基因的抗反饋抑制的能力,采用PCR介導的方式對lysC基因進行另一關鍵位點的定點突變。獲得的陽性克隆中提取質粒并進行測序分析,結果顯示,突變后的lysCr-asdA基因與突變前的基因差異在第279位密碼子由GCT突變成ACT,所編碼的氨基酸由丙氨酸變為蘇氨酸,與定點突變設計目標一致,表明表達載體pZ8-1-lysCrasdA構建成功。
2.3 工程菌株獲得及其篩選
將獲得的含有抗反饋抑制突變的串聯表達質粒 pZ8-1-lysCr-asdA電轉化至L-蘇氨酸菌株C.glutamicumT11感受態細胞,采用含卡那霉素抗性平板進行篩選,隨機挑取單菌落,驗證引物F和R組成的引物對其進行菌落PCR鑒定,結果顯示在約2 800 bp的特異條帶,與理論預測大小一致,說明外源質粒導入宿主菌株,成功獲得含有表達載體的工程菌。
挑選了300個單克隆進行96孔板初篩,并從中獲得10株生長速度較快且L-蘇氨酸含量較高的菌株進行搖瓶復篩和遺傳穩定性驗證。結果顯示,選取的10株相對高產的菌株中,幾株菌株傳至第二代遺傳穩定性逐漸降低,在斜面傳至第三代時,產酸明顯下降,再繼續傳代,L-蘇氨酸產量下降最高達到14.4%。相對而言,其中一株菌株L-蘇氨酸產量最高,而且穩定性維持在較高水平,具備進行進一步發酵驗證和分析的潛力,將該工程菌株菌株命名為T11/pZ8-1-lysCr-asdA。
2.4 工程菌株T11/pZ8-1-lysCr-asdA搖瓶發酵產L-蘇氨酸能力分析
為了分析工程菌株T11/pZ8-1-lysCr-asdA發酵合成L-蘇氨酸的能力,將其與菌株T11/pZ8-1和T11/pZ8-1-lysC-asdA及出發菌株T11在相同條件下進行搖瓶發酵培養,考察L-蘇氨酸濃度、主要雜酸情況及菌株生物量(OD562nm),結果見表1。
從L-蘇氨酸的生物合成和生物量來說,菌株T11/pZ8-1-lysCr-asdA的L-蘇氨酸的濃度和細胞密度都最高,T11/pZ8-1-lysC-asdA次之,這說明串聯基因簇的表達對于L-蘇氨酸的合成與積累作用顯著,而且通過調控其他氨基酸的生物合成,一定程度上對細胞的生長代謝帶來了益處;轉入空載體pZ8-1/T11菌株無論是細胞密度還是L-蘇氨酸濃度都是最低的,這可能與空載體在宿主細胞內消耗了一定的能量,從而影響細胞繁殖和氨基酸的代謝合成有關。

表1 T11/ pZ8-1-lysCr-asdA發酵性能比較
另外,從主要雜酸的生物合成情況來看,出發菌T11與對照菌株T11/pZ8-1的賴氨酸、丙氨酸、和甘氨酸含量接近,而含有串聯基因簇的工程菌株,其丙氨酸含量顯著降低,但賴氨酸和甘氨酸含量有所提高,說明lysC-asdA的串聯表達加速了前體物丙酮酸的轉化,有效地將碳流量引入了合成蘇氨酸的合成途徑,顯著提高天冬氨酸族合成代謝流,但這其中碳流向不僅包括蘇氨酸的生物合成,還有一部分流向了賴氨酸的生物合成,而甘氨酸作為蘇氨酸的降解途徑之一,其生物合成的增加意味著更多的蘇氨酸在細胞內被降解。
相比較T11/pZ8-1-lysC-asdA與T11/pZ8-1-lysCrasdA菌株,lysC基因的突變對于L-蘇氨酸的生物合成有積極意義,其進一步提高了lysC基因編碼產物抗反饋抑制的能力,增強了L-蘇氨酸生物合成代謝流。而對于提高 L-蘇氨酸產量,lysCr-asdA的定點突變和串聯表達較lysC-asdA串聯表達具有更顯著的作用。
2.5 工程菌株T11/pZ8-1-lysCr-asdA 30L罐的補料分批發酵
為進一步分析T11/pZ8-1-lysCr-asdA工程菌株的代謝情況,以出發菌株T11為對照,在30 L 全自動發酵罐中進行補料分批發酵,考察菌株的發酵生產性能。發酵過程代謝曲線見圖5,工程菌株T11/pZ8-1-lysCr-asdAL-蘇氨酸發酵產量達65.5 g/L,糖酸轉化率達 39.5%,較出發菌株分別提高29.5%和33.9%。
發酵0-16 h,菌株處于適應期,細胞密度均增長較慢,但細胞的耗糖速率有差別,工程菌株T11/pZ8-1-lysCr-asdA的耗糖速率明顯要大,說明其細胞更為活躍,代謝更為旺盛;發酵16 h之后,細胞均進入對數生長期,菌株T11/pZ8-1-lysCr-asdA在發酵24 h開始流加補糖,36 h時達到最大菌濃,其最大菌濃時OD562nm為112.6,補糖時間和達到最大菌濃時間均較對照菌株提前6 h,最大菌濃也較對照菌株T11高16.8%;發酵36 h后,L-蘇氨酸的積累速率明顯加快,兩者均在發酵56 h時達到最大的產酸率,隨著發酵的進行,L-蘇氨酸的積累和生物量在56 h和60 h之間降低,因此,發酵在60 h終止。
3 討論
AK是整個天冬氨酸族氨基酸合成途徑上的第一個關鍵酶,由lysC基因編碼,控制合成天冬氨酸族氨基酸的碳流量。ASD是繼AK后另一個關鍵限速酶,這兩個基因的轉錄皆受L-賴氨酸、L-蘇氨酸反饋阻遏調控。故對于構建天冬氨酸族氨基酸如L-賴氨酸、L-甲硫氨酸、L-蘇氨酸、L-異亮氨酸高產菌株具有重要意義。Kalinowski等[13-14]發現lysC基因與asdA基因是同一基因簇的相鄰基因,其中lysC基因編碼蛋白由兩個亞基α和β相互重疊組成,而當β亞基中的一個堿基發生改變時,整個AK蛋白表現抗反饋特性。本研究以一株Met-、Ile-和蘇氨酸結構類似物抗性的蘇氨酸產生菌為出發菌株,在獲得已鑒定突變的基礎上,進一步對lysC基因的第279位密碼子進行定點突變,核苷酸由GCT突變為ACT,所編碼的氨基酸由丙氨酸突變蘇氨酸,氨基酸由疏水性變為親水性,這可能進一步改變AK的分子構象,可能Ala279位點具有穩定配體Lys與AK結合的作用,從而加強了Lys對AK的反饋抑制。故將該位點改變打破了其與Lys間的非共價鍵作用,從而解除賴氨酸對天冬氨酸激酶的反饋抑制,削弱L-蘇氨酸和L-賴氨酸對AK酶抑制能力,同時提升了磷酸-天冬氨酸對AK的親和能力,從而增強L-蘇氨酸代謝流。這一點,與工程菌株的蘇氨酸生物合成和積累的情況一致,lysC的突變有效的提高了蘇氨酸合成的碳流量,使L-蘇氨酸產量提高。

加速限速反應作為菌株選育的常用策略,除了解除終產物的反饋抑制手段之外,關鍵酶基因的高效表達則更能從關鍵酶的表達量上實現限速反應的加速進程。Ishida等[15]通過構建重組質粒pDR34,在乳糖發酵短桿菌同時串聯表達了hom、thrB和thrC基因,發酵72 h使得L-蘇氨酸產量從17.9 g/L達到24.8 g/L。Dong等[16]將谷氨酸棒桿菌ATCC 13869 敲除ddh基因和lysE基因使蘇氨酸產量提高28%,賴氨酸的量降低95%,同時串聯表達lysC、hom和thrB基因進行72 h搖瓶發酵驗證蘇氨酸含量達到7.27 g/L。本研究以強啟動子tac作為起始元件,在已有菌株特性的基礎上,串聯表達抗反饋抑制的關鍵酶基因,極大的提高了蘇氨酸產量。但在甲硫氨酸和異亮氨酸合成受阻的基礎上,蘇氨酸的另一支路——賴氨酸的合成同樣得到了增強,不僅如此,作為蘇氨酸的分解代謝產物之一的甘氨酸的含量也有所增加,其實這些都和關鍵酶基因表達之后的代謝流增強有關。這也提示我們在關注蘇氨酸積累的同時,需要進一步系統的考慮其他的支路代謝和氨基酸轉運體系,比如阻斷蘇氨酸競爭代謝支路,即抑制或敲除賴氨酸的表達;降低蘇氨酸胞外降解途徑,即敲除合成甘氨酸相關酶基因,運用代謝流分析的原理與方法對蘇氨酸代謝途徑進行全面分析,為我們進一步的改造明確方向。
與搖瓶發酵不同,30 L罐通過流加補料的方式保持細胞的生長和代謝性能,其目標產物的產量往往會得到大幅度的增長。值得注意的是,本研究中工程菌株T11/pZ8-1-lysCr-asdA相較出發菌株T11來說,其較出發菌株糖耗時間提前,菌株生長速度加快,最終體現為菌株發酵產酸時間提前,展現了更好的高密度發酵的適應性,較出發菌株其生長速率更快,原因可能一是改造后菌株的前體物丙酮酸轉化成生長抑制物如乳酸、乙酸量降低,從而減少了細胞生長的傷害,相對的加速其轉化成草酰乙酸方向,從而使碳流量更多的合成蘇氨酸的方向。二是lysCr-asdA基因與含強啟動子的pZ8-1載體串聯表達不僅使菌株的AK和ASD酶高效表達,也使碳流量方向途徑中其他的酶活得到促進,從而代謝速率加快,而且在流加補糖的過程中保持較高的L-蘇氨酸的比合成速率,這說明外源基因的表達不僅直接促進了蘇氨酸的合成,對于細胞生長和其他所需氨基酸的代謝調控也要一定的影響。
4 結論
本研究以一株Met-、Ile-和AHV抗性的蘇氨酸產生菌T11為出發菌株,在獲得已鑒定突變的基礎上,進一步對lysC基因的第279位密碼子進行定點突變同時串聯基因簇lysCr-asdA。成功構建pZ8-1-lysCr-asdA表達組件,串聯表達抗反饋抑制的關鍵酶基因,轉入出發菌株T11并通過96孔板高通量初篩和搖瓶復篩得到一株高產L-蘇氨酸且遺傳穩定工程菌T11/pZ8-1-lysCr-asdA。發酵結果顯示其 L-蘇氨酸產量顯著提高,L-賴氨酸、L-甘氨酸含量均比出發菌株提高,而丙氨酸的含量下降,表明lysC的定點突變及lysC、asdA基因的串聯表達對谷氨酸棒桿菌L-蘇氨酸的代謝流有明顯影響。
