高溫微生物鐵還原條件下綠脫石對有機質的保存作用研究*
2019-02-18 07:04:56曾強董海良汪丹
巖石學報 2019年1期
曾強 董海良,2** 汪丹
1.中國地質大學地球科學與資源學院,北京 100083
2.中國地質大學生物地質與環境地質國家重點實驗室,北京 100083
碳循環問題是當今備受科學界關注的熱點問題。與無機碳庫相比,有機碳庫的循環速度更快,其高度動態化的性質對人類活動和氣候變化等問題都具有著深遠的影響(Sombroeketal.,1993)。而有機質的性質和賦存狀態是影響碳循環速率的重要因素。地表環境中絕大多數有機碳都與多種多樣的礦物結合在一起。不同的礦物類型影響著有機質與礦物結合的方式及其穩定性(Keil and Mayer,2013)。因此,研究多種的環境因素影響下這些礦物-有機質復合體的穩定性,對于揭示地表環境復雜的碳循環過程具有十分重要的啟示意義。
粘土礦物是一種由硅氧四面體和金屬(Al,Fe,Mg)氧八面體片構成基本單元的層狀硅酸鹽,這些細小礦物廣泛分布于土壤、沉積物、沉積巖等地表環境,許多粘土礦物結構都含有不同程度的鐵,可以說粘土礦物是鐵的重要載體之一(Stuckietal.,2012)。粘土礦物由于具有巨大的比表面積、陽離子交換能力、以及部分種類具有可膨脹的層間域,使其在環境中能夠結合大量的有機質。粘土礦物結構鐵的氧化還原態變化(即 Fe(II)■Fe(III))對于粘土礦物的理化性質有著深刻的影響(Stucki,2011),而這一變化也可能進一步影響粘土礦物與有機質的結合狀態。其中,粘土礦物結構Fe(III)的微生物還原過程被廣泛地發現于各類環境之中(Dongetal.,2009)。許多功能微生物類群都能利用粘土礦物中的結構Fe(III)作為電子傳遞的受體。之前的研究發現,一些常溫至中溫的具有鐵還原能力的細菌能夠通過結構Fe(III)還原作用釋放少部分插入粘土礦物層間域的有機質,而大部分的有機質能夠在還原-氧化波動的環境中保存下來,說明某些粘土礦物的可膨脹層間域可以有效地保存有機質,能夠抵御外界氧化還原條件波動對有機質礦化的影響(Zhangetal.,2014;Zengetal.,2016)。然而這一過程在中高溫的條件下是否同樣存在,目前尚未有任何報導。本研究的目的就在于探究兩株中高溫的異化鐵還原菌對于插層態的粘土礦物-有機質復合體的還原作用以及該過程對有機質的釋放效果。
1 材料與方法
1.1 綠脫石-有機質復合體的制備
實驗選擇一種產自澳大利亞Uley礦床的綠脫石作為含鐵粘土礦物的代表(稱為NAu-2),主要因為其具有較高的結構鐵含量,其含鐵量約為21.2%,其中98%的結構鐵均為Fe(III)(Keeling,2000)。12-氨基十二酸(12-Aminododecanic Acid,ALA)作為一種模式有機物用于合成有機質-粘土復合體。選用ALA作為模式有機物主要基于兩個原因:一是它含有羧基以及適當碳數目的烷基鏈,這是環境中與粘土礦物結合的有機物的重要特征(Wattel-Koekkoeketal.,2001);二是ALA具有的丙氨基基團(-NH2),在酸性條件下可以通過質子化轉化為氨基基團(-NH3+),從而通過陽離子交換作用插層進入NAu-2的層間域。
ALA插層實驗前,首先將塊狀的NAu-2人工研磨過篩,然后配置成10g/L的懸濁液,充分攪拌24h使其均勻分散。而后,配置濃度約為該懸濁液的NAu-2兩倍陽離子交換量(CEC)的ALA溶液。將ALA加入到粘土溶液前,首先通過用鹽酸調節ALA溶液的pH將其質子化(0.07N HCl、pH=1.14),并加熱至80℃攪拌至溶液澄清為止。隨后,將質子化的ALA溶液和粘土混合,并繼續再80℃的水浴條件下攪拌半小時,此時的溶液pH約為4.5。之后,合成的綠脫石-有機質復合體(簡稱為ALA-NAu-2)通過離心的方法收集,并用80℃的超純水洗滌5次去除任何游離態的以及微弱吸附的ALA分子。
1.2 菌株培養
高溫異化鐵還原菌ThermusscotoductusSA-01由美國邁阿密大學地質微生物實驗室贈送,經甘油冷凍保存于中國地質大學(北京)地質微生物實驗室。其是一株兼性厭氧微生物,最早從南非金礦中分離出來(Kieftetal.,1999)。其最適生長溫度為65℃,采用TYG培養基培養(5g胰蛋白胨、3g酵母提取物、1g葡萄糖/L)。超高溫異化鐵還原菌Geoglobus ahangari購買于德國微生物菌種保藏中心,該菌分離自瓜伊馬斯盆地熱液系統深部2000m處(Kashefietal.,2002),其最適生長溫度為85℃,采用DSMZ 1210培養基進行培養。
1.3 微生物還原綠脫石-有機質復合體
首先將原始的綠脫石原土NAu-2及合成的ALA-NAu-2配置成5g/L的溶液。實驗采用寡營養培養體系,即體系中只有電子供受體為菌體提供能量來源。高溫菌SA-01體系中,乳酸鈉(最終濃度20mM)為唯一的電子供體,粘土礦物結構Fe(III)為唯一的電子受體。超高溫菌G.ahangari體系中,乙酸鈉(最終濃度20mM)為唯一的電子供體,粘土礦物結構Fe(III)為唯一的電子受體。每個還原實驗組都另附一組相同條件下添加電子穿梭體AQDS(蒽醌-2,6-二磺酸,最終濃度0.1mmol/L)的實驗組。電子穿梭體可以極大地提高微生物還原粘土礦物結構鐵的速率。粘土溶液在高溫滅菌之后,將生長至對數期的菌體通過離心收集菌體,然后運用吖淀橙對菌體進行染色在熒光顯微鏡下計數,最后將一定數量的菌體投入反應體系中開始鐵還原反應。
1.4 分析方法
1.4.1 結構 Fe(II)檢測
對于結構Fe(II)的分析采用氫氟酸/硫酸提取法溶解粘土礦物的硅酸鹽結構(Amonette and Templeton,1998)。取0.2mL粘土溶液于裝有0.48mL、3.6N H2SO4的黑色避光離心管中,加入 0.08mL、10%的 1,10-phenanthroline顯色劑,0.04mL、48%的HF溶液,水浴加熱煮沸30min,以促進粘土礦物結構的溶解,然后室溫下冷卻15min,接著加入0.4mL硼酸溶液中和體系中多余的HF,再取0.1mL樣品于1mL 1%的檸檬酸鈉溶液中,通過分光光度計在OD=510nm吸光度下進行檢測。
1.4.2 溶解態 Si、Al、Fe濃度測試
實驗進一步采用電感耦合等離子體發射光譜儀(ICPOES)對反應及對體系中上清液的溶解態Si、Al、Fe分別進行測試。探究經過微生物的鐵還原作用是否會導致粘土礦物的溶解。取粘土礦物懸濁液1mL,離心后將上清液過濾(0.4μm聚醚砜濾膜),將樣品稀釋到一定倍數之后上機測試。
1.4.3 總有機碳測試(TOC)
TOC(Total Organic Carbon)用來表征粘土礦物中有機碳的總量,可以直觀的檢測有機質ALA在粘土礦物中的含量變化,探究微生物的鐵還原作用對有機質的保存是否有影響。首先在厭氧手套箱中取粘土礦物懸濁液5mL,離心后去除上清液,然后用除氧水洗滌三遍,洗去體系中殘留的乳酸鈉和乙酸鈉。待樣品干燥后,取15mg樣品上機測試。實驗使用德國耶拿總C/N 2100s分析儀,工作溫度1000℃。
1.4.4 X射線衍射(XRD)礦物學表征
XRD用于檢測ALA插層綠脫石的效果,以及生物還原后插層和未插層的NAu-2在礦物結構上的變化情況。樣品采用涂片法制備,粘土溶液在厭氧手套
箱中用除氧水洗滌三遍后,均勻的涂抹于巖相學玻璃片上,于厭氧箱中自然晾干后準備上機測試。儀器的工作條件為:儀器功率8000W(管壓40kV,管流200mA),掃描范圍3°~15°(2θ),步長 0.02°,掃描速度 0.01°/s。
1.4.5 傅里葉變換紅外光譜分析(FTIR)
FTIR可以用來表征有機質ALA進入到粘土礦物層間后及微生物作用后引起的有機質化學鍵振動變化,并以此可以分析有機質的插層和脫附情況。在厭氧手套箱中取0.5mL粘土溶液樣品后,離心后去除上清液,并用除氧水洗滌三遍,待樣品在手套箱中自然晾干后,取2mg粉末樣品與200mg KBr迅速充分研磨,壓片后上機測試。掃描范圍為中紅外400~4000cm-1,掃描次數50次。
1.4.6 掃描電鏡觀察(SEM)
SEM用于觀察微生物反應前后NAu-2和ALA-NAu-2礦物形貌和元素成分上的變化。具體制樣與檢測方法如下:將粘土懸濁液樣品滴在干凈的玻璃載玻片上,以濃度為2.5%的戊二醛和2%多聚甲醛為固定液對礦物-微生物混合液進行固定。之后將樣品放入不同濃度的乙醇(25%、50%、75%、95%、100%)之中進行梯度脫水。處理好的樣品放入液態CO2臨界點干燥儀(Quorum K850 Critical Point Dryer,CPD)中進行干燥。干燥后的樣品放置在SEM樣品臺上進行噴Pt處理(Quorum SC7620 Sputter Coater)。準備完畢后用SEM(Zeiss Supra 55 SAPPHIRE)對其進行微觀形貌的觀察,同時利用能譜儀進行礦物化學成分的分析。SEM工作的加速電壓為8~15kV,工作距離為15mm。對于形貌觀察采用及低電流條件(30~40μA)已達到最好的成像效果。對于能譜分析采用較高的電流(50~70μA)以達到更好的信號強度。
2 實驗結果
2.1 綠脫石-有機質復合體的表征
通過掃描電鏡結果顯示,ALA-NAu-2和NAu-2的微觀結構沒有太大的區別,都表現出蒙脫石典型的不規則薄片特征,邊緣卷曲且互相堆疊(圖1a,b),其中ALA-NAu-2碳的含量比NAu-2高,含碳量從不到1%增加至7.455%,間接地說明了ALA與NAu-2相結合(圖1a1,b1)。
XRD的結果顯示,綠脫石NAu-2經過有機質插層后,合成的有機粘土ALA-NAu-2其層間距從12.389?增加到16.747?,表明ALA成功的進入了NAu-2的層間域(圖2)。另一方面,ALA-NAu-2相較于NAu-2,其d(001)峰寬變大,表明有機質進入層間降低了粘土礦物片層結構的有序度。
FTIR的結果表明,ALA-NAu-2在原始礦物NAu-2的基礎上呈現出了許多新的特征峰,其中包含有機質ALA的特征峰和一些合成過程中新形成的特征峰。如3118cm-1處的N-H伸縮振動峰、2925cm-1和2853cm-1處的C-H反對稱和對稱伸縮振動峰、1713cm-1處的 C=O伸縮振動峰、1626cm-1處的O-H變形和N-H彎曲振動峰、1515cm-1處的R-COO-反對稱伸縮振動峰、1466cm-1處的CO-H彎曲振動峰以及1400cm-1處的R-COO-反對稱伸縮振動峰(圖3)。
這些證據共同表明,ALA進入了NAu-2的層間結構,并且ALA的官能團與粘土礦物的片層發生了相互作用。
2.2 微生物還原粘土礦物結構Fe(III)
SA-01與G.ahangari均能夠在添加AQDS的情況下迅速地還原NAu-2和ALA-NAu-2中的結構Fe(III)。但是由于G.ahangari本身不能分泌具有類似電子穿梭體性質的物質,因此在不添加AQDS的情況下不能還原結構Fe(III)(結果未顯示)。兩株菌的還原程度和還原速率都有所不同。SA-01實驗組在反應進行了100天時才停止,而超高溫菌G.ahangari在反應進行7天的時候還原已經達到平衡,但是實驗一直取樣到30天,以保證微生物還原反應徹底停止。
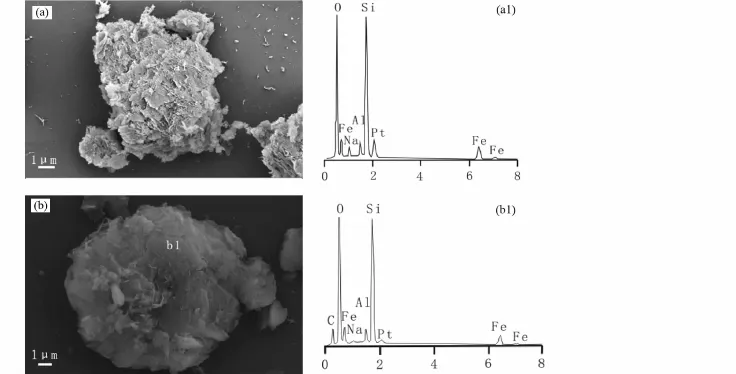
圖1 NAu-2(a)和 ALA-NAu-2(b)掃描電鏡圖像及對應的 EDS(a1、b1)譜圖Fig.1 Scanning electron microscope(SEM)images of NAu-2(a)and ALA-NAu-2(b)and their corresponding energy dispersive spectroscopes(a1,b1)
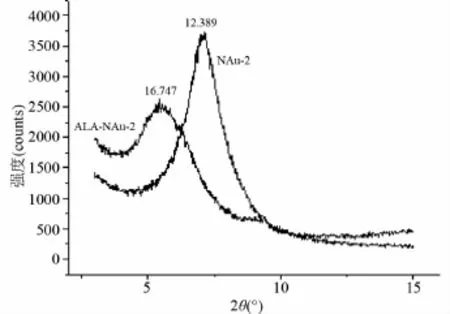
圖2 NAu-2和ALA-NAu-2 X射線衍射圖Fig.2 X-ray diffraction(XRD)patterns of NAu-2 and ALA-NAu-2
SA-01還原綠脫石NAu-2(圖4a)前24h反應速率分別為0.04mM/h(不含 AQDS)和0.099mM/h(含 AQDS),還原ALA-NAu-2(圖4b)前24h反應速率分別為0.033mM/h(不含AQDS)和0.062mM/h(含AQDS)。反應100天結束時,NAu-2實驗組Fe(III)還原程度達到18.48% (不含AQDS)和22.17% (含 AQDS),ALA-NAu-2實驗組分別達到14.42% (不含 AQDS)和18.14% (含 AQDS),比 NAu-2實驗組反應程度要低。
G.ahangari在都添加了AQDS的情況下,還原綠脫石NAu-2前24h反應速率為0.28mM/h,還原 ALA-NAu-2前24h反應速率為0.15mM/h。反應結束時,綠脫石NAu-2實驗組Fe(III)還原程度達到28.56%,有機粘土礦物 ALANAu-2實驗組達到23.21%。微生物在還原NAu-2約1.5天后,還原速率急劇下降,還原程度增長幅度減慢直至反應停止,而此時ALA-NAu-2實驗組反應仍在持續進行(圖4c)。
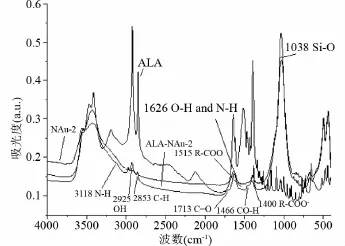
圖3 ALA、NAu-2和ALA-NAu-2復合物傅里葉轉換紅外光譜圖Fig.3 Fourier Transform infrared spectroscopy(XRD)patterns of ALA,NAu-2 and ALA-NAu-2
以上結果說明,微生物對有機粘土礦物ALA-NAu-2的還原程度和還原速率都比粘土礦物NAu-2低,表明有機質進入粘土礦物的層間不利于微生物對結構Fe(III)的還原,這與前人的結果相類似(Zhangetal.,2014;Zengetal.,2016)。
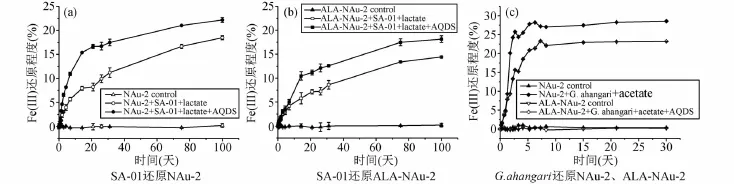
圖4 SA-01對NAu-2的還原曲線(a)、SA-01對ALA-NAu-2的還原曲線(b)以及G.ahangari還原NAu-2和ALA-NAu-2的曲線(c)Fig.4 Reduction curves of NAu-2 by SA-01(a),NAu-2 by SA-01(b),and NAu-2 and ALA-NAu-2 by G.ahangari(c)
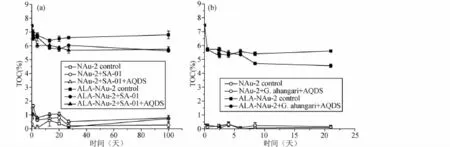
圖5 SA-01(a)和G.ahangari(b)實驗體系中總有機碳隨時間變化曲線Fig.5 Changes of total organic carbon(TOC)in SA-01(a)and G.ahangari(b)reaction system
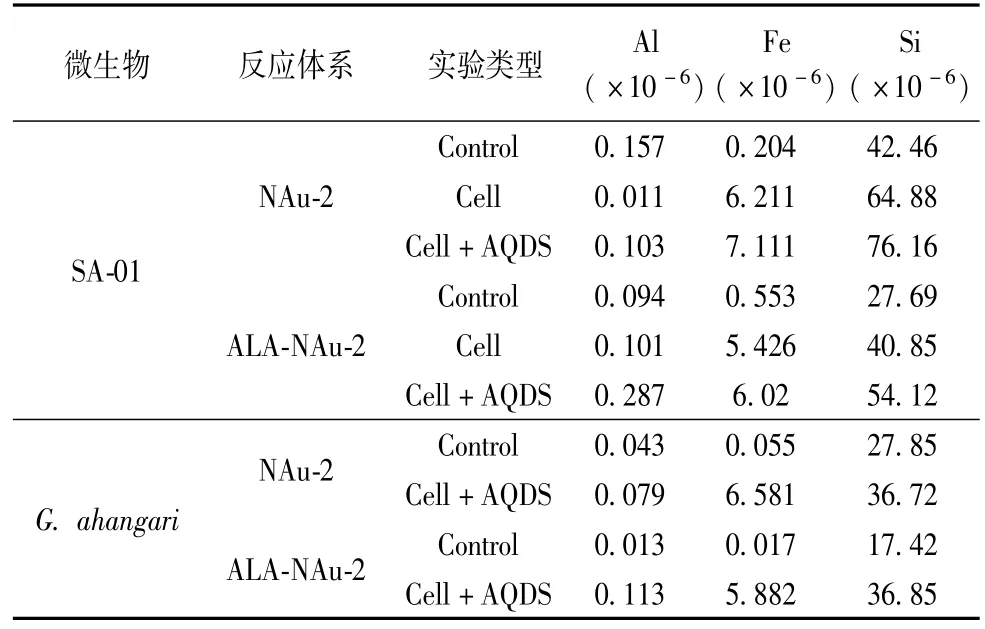
表1 可溶性 Si、Al、Fe離子濃度Table 1 Concentrations of soluble Si,Al and Fe
2.3 溶解態 Si、Al、Fe離子濃度
無菌對照實驗組中可溶性Si、Al、Fe的濃度都很低(表1)。微生物對NAu-2和ALA-NAu-2結構Fe(III)的還原明顯導致了水溶液中溶解態Si、Al、Fe離子濃度的升高。并且可溶性陽離子的濃度隨著還原程度的升高而升高。表明微生物對粘土礦物和有機粘土礦物的還原過程中時礦物發生了部分溶解作用。
2.4 TOC結果
高溫菌SA-01還原體系中(圖5a),NAu-2+細胞和NAu-2+細胞+AQDS實驗組其總有機碳含量比對照組略高,這可能來源于實驗體系中添加的微生物和培養基中的少量有機碳帶入。但是從圖5a中可以看出,微生物和培養基中有機碳含量很低,因此相對于ALA-NAu-2的體系來說,幾乎可以忽略不計。在有機粘土礦物ALA-NAu-2實驗體系中,無菌對照組在65℃下放置100天后,總有機碳含量從7.46%下降到6.80%,表明高溫環境下會有少量有機質從粘土礦物層間釋放出來,約占總有機碳含量的8.6%;未添加AQDS的微生物實驗組,粘土礦物中14.42%的結構Fe(III)被還原,總有機碳含量從7.46%下降到5.64%,表明實驗體系中仍有75.60%(剩余有機碳含量/初始總有機碳含量)的ALA保存在粘土礦物結構中;添加 AQDS的實驗組,粘土礦物中18.14%的結構Fe(III)被還原,總有機碳含量從7.46%下降到5.75%,實驗最終有77.13%的有機質保存在粘土礦物結構中。總體來說,有機質ALA在粘土礦物層間具有很好的穩定性,無菌環境下,即使是在65℃的高溫下也能很好的保存下來。微生物對有機粘土礦物進行改造的過程中,盡管一部分有機質從層間釋放出來,但是大于75%的有機質依舊保存在粘土礦物結構中。
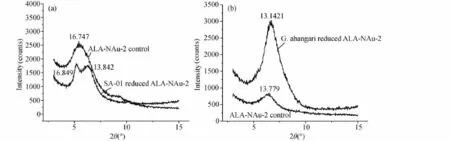
圖6 SA-01(a)和G.ahangari(b)實驗體系中ALA-NAu-2 XRD譜圖Fig.6 XRD patterns of ALA-NAu-2 in SA-01(a)and G.ahangari(b)reaction system
G.ahangari還原體系中(圖5b),有機粘土礦物 ALANAu-2無菌對照組在85℃高溫環境下,第一個取樣點(0.5天)顯示其總有機碳含量直接從7.46%下到5.71%,僅剩76.5%的有機碳保存在粘土礦物層間(剩余有機碳含量/初始總有機碳含量),相比于SA-01的有機粘土礦物無菌對照組反應結束時仍有91.4%保存在礦物結構中,說明更高的溫度會進一步促進ALA的釋放。當ALA-NAu-2無菌對照組在高溫環境下釋放出一部分有機質后,直至微生物反應停止30天時,其所含的有機質含量為5.6%,占原始總有機碳含量的75.1%,相比于第一個點的測量值5.71%,此過程中僅有0.11%的有機碳釋放出來。總體來說,當有機粘土ALANAu-2在放置于85℃環境下,一些表面或是層間吸附不牢固的ALA會立刻從粘土礦物中釋放出來,剩余大于75%的有機質仍然與粘土礦物之間有很強的吸附,二者存在的狀態穩定并且不受高溫的影響。
從G.ahangari還原有機粘土ALA-NAu-2的曲線(圖4c)可以看出,隨著微生物對結構Fe(III)還原程度的升高,有機質ALA不斷從粘土礦物的層間釋放出來。當微生物還原停止時,有機質的含量也不再下降,并且保持穩定。由于微生物的還原能力有限,實驗最終仍有4.54%的有機質被保存下來,占據初始總有機碳含量的60.9%。總之,在溫度穩定之后,微生物的鐵還原作用會進一步導致一部分有機質繼續從粘土礦物層間釋放出來,但是受限于較低的鐵還原程度,最終仍然有大于60%的有機質保存在粘土礦物中。
2.5 XRD結果
SA-01還原有機粘土前后XRD譜圖(圖6a)顯示,經過微生物的鐵還原作用后,有機粘土礦物的d(001)從16.747?的單峰轉變為16.849?和13.842?的雙峰。其中16.849?處的峰比原始ALA-NAu-2主峰16.747?變得更加尖銳。而13.842?的峰與沒有插層的綠脫石d(001)值接近,呈現出NAu-2的特征。表明在微生物還原有機粘土礦物ALA-NAu-2的過程中,一部分的ALA會從層間結構中脫附出來,最終導致層間距減小,并接近于插層前的層間距。
G.ahangari還原有機粘土前后XRD譜圖(圖6b)顯示,置于85℃環境下的對照組ALA-NAu-2相對于原始ALANAu-2,其層間距d(001)從16.849?降至13.779?,層間距減小,表明更高的溫度會促進更多ALA從粘土礦物的層間釋放出來,這一結果與總有機碳測試結果相吻合(圖5b)。加入微生物G.ahangari的實驗組,ALA-NAu-2的 d(001)從16.747?降至13.142?,與沒有插層的綠脫石 d(001)值接近,呈現出NAu-2的特征。另外,相對于無菌對照實驗組,經過微生物還原后的有機粘土礦物峰變的更加尖銳,且峰強增大,說明經過微生物作用后綠脫石的結晶程度有所提高,在反應過程中小顆粒與結晶度較差的綠脫石可能優先發生溶解。
2.6 FTIR結果
FTIR的結果顯示,兩株細菌的鐵還原作用均對ALA與NAu-2的結合產生了一定影響,但是兩株菌之間還有一定差異。依照前人對綠脫石NAu-2的紅外光譜的分區經驗(Neumannetal.,2011),將有機粘土礦物 ALA-NAu-2的紅外光譜分為5個區域:3800~3200 cm-1、3200~2500 cm-1、1800~1500 cm-1、1200~900 cm-1和 1000~700cm-1,分別指示蒙脫石八面體結構中νM-OH(M代表八面體片中的Mg、Al或Fe)伸縮振動帶,C-H伸縮振動帶,O-H變形振動帶,Si-O伸縮振動帶以及八面體中νM-OH彎曲振動帶。
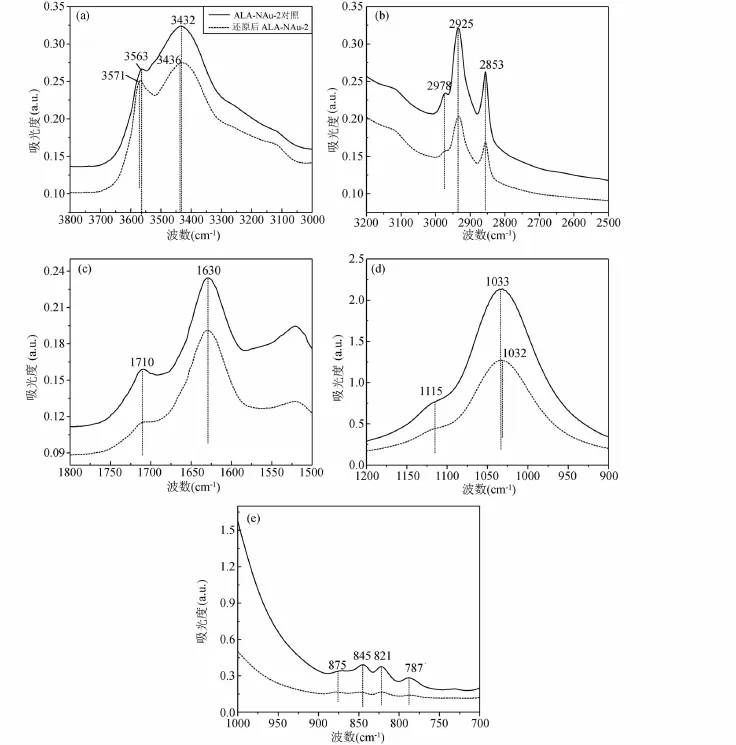
圖7 對照組及SA-01還原后ALA-NAu-2紅外譜圖Fig.7 FTIR patterns of control group and reaction group by SA-01
ALA-NAu-2經過SA-01的還原后,峰強都有明顯的減弱,但是峰位沒有太大的偏移。3563cm-1處的νM-OH伸縮振動峰在ALA-NAu-2還原后峰位移到3571cm-1,3432cm-1處為吸附水峰。圖7b中顯示的是有機質ALA特有的C-H伸縮振動峰,經過微生物作用過后峰強略有減弱,表明即使微生物對粘土礦物結構Fe(III)進行還原,仍然有一部分有機質存留在粘土礦物的層間。圖7c中的O-H變形振動峰在還原后也略微減弱,圖7d中1115cm-1處的Si-O峰經微生物作用后基本消失,1033cm-1處的Si-O峰強度減弱,峰形變得寬緩,且略微出現偏移。νM-OH變形振動帶(圖 7e)在875cm-1、845cm-1、821cm-1、787cm-1分別指示 AlFe-OH、AlMg-OH、FeFe-OH以及FeMg-OH振動峰,還原后該區的峰都明顯減弱,875cm-1處的峰幾乎消失。總體來說還原作用對ALA-NAu-2礦物結構內化學鍵振動也產生了一定影響。
與SA-01相比,經G.ahangari作用前后紅外譜圖顯示,還原后的ALA-NAu-2峰的位置也未發生太大偏移,但是峰強的減弱更為明顯,有些甚至消失不見。圖8a中3563cm-1處的νM-OH伸縮振動峰在還原反應后幾乎消失。圖8b中有機質ALA的C-H伸縮振動峰,經過微生物作用過后峰強明顯減弱更加明顯,但是峰形并沒有完全消失,表明微生物還原結束后,仍有部分有機質保存在粘土礦物的層間。圖8c中的O-H變形振動峰在還原后略微減弱,圖8d中1115cm-1處的Si-O峰經微生物作用后基本消失,1038cm-1處的Si-O峰強度減弱,峰形變得寬緩。圖8e中νM-OH變形振動帶還原后該區的峰都明顯減弱,880cm-1處的峰幾乎消失。總體來說與 SA-01相似,ALA-NAu-2經過G.ahangari作用后ALA的特征峰及礦物結構內化學鍵振動也發生了明顯的變化。
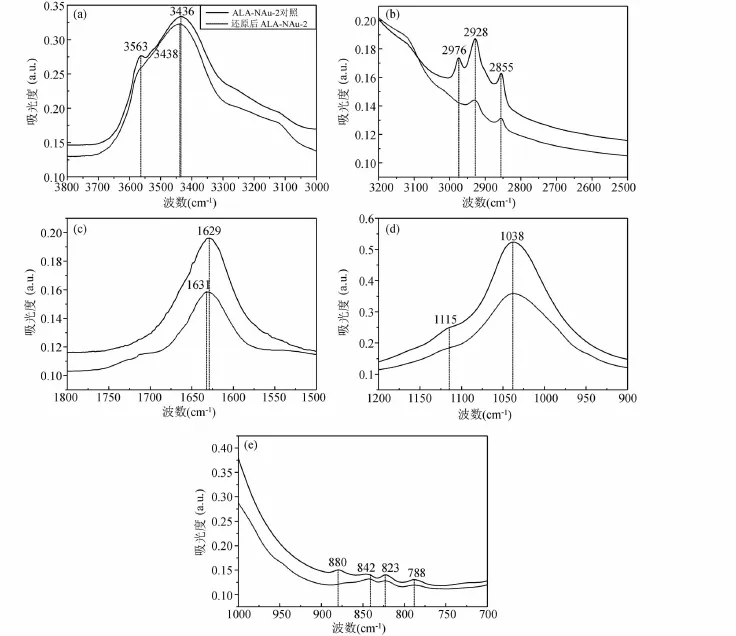
圖8 對照組及G.ahangari還原后ALA-NAu-2紅外譜圖Fig.8 FTIR patterns of control group and reaction group by G.ahangari
2.7 SEM結果
SEM的結果顯示,經過微生物還原的NAu-2,表面轉變成疏松多孔的絮狀物,并呈現出凌亂的網狀結構(圖9a)。EDS(圖9b)顯示,經過微生物作用后的礦物,其Fe含量有所降低,有機粘土礦物經微生物作用后,有機碳的含量也有所降低,這一結果與TOC、XRD的結果都有很好的對應關系。
SA-01對ALA-NAu-2還原后,由于粘土礦物結構被破壞,礦物發生還原溶解和再沉淀反應,在掃描電鏡下觀察到一系列粘土礦物溶解所產生的納米SiO2聚合物(圖9c)。
3 討論
3.1 有機質對微生物還原粘土礦物的影響
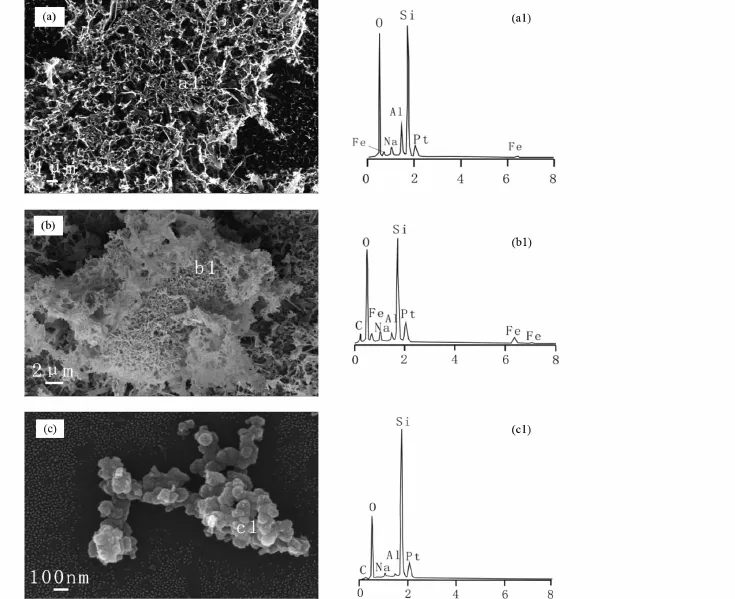
圖9 還原后NAu-2(a)、ALA-NAu-2(b)和新礦物納米二氧化硅(c)掃描電鏡圖像及其對應的EDS譜圖(a1、b1、c1)Fig.9 SEM images and corresponding EDSof reduced NAu-2(a),ALA-NAu-2(b)and newly-formed nanosized silica and their corresponding energy dispersive spectroscopes(a1,b1,c1)
從高溫菌SA-01和超高溫菌G.ahangari還原粘土礦物、有機粘土礦物的結果來看,二者均能對NAu-2和ALANAu-2中的結構Fe(III)進行還原,由于兩種粘土礦物的性狀不同,致使ALA-NAu-2的還原程度和還原速率均比NAu-2要低。這可能有以下幾個原因:首先質子化的ALA可以通過置換綠脫石NAu-2的陽離子和層間水,從而進入粘土礦物的層間。目前的研究指出,一些具有鐵還原功能的微生物類群在還原粘土礦物結構鐵的過程中,其電子傳遞的方式主要通過平行于粘土礦物d(001)結構層的方向進行(Zhaoetal.,2015;Shietal.,2016)。即微生物首先會還原粘土礦物片層邊部八面體片中的結構Fe(III),而后電子不斷以平行于d(001)面的方向向粘土礦物內部傳輸。一些研究也通過模型計算方法證明了,相比于通過四面體片層的垂直d(001)方向的電子傳遞途徑,這種電子傳遞方式更容易實現(Alexandrovetal.,2013)。但是,還是有部分研究表明,在添加了電子穿梭體AQDS的情況下,微生物還原過程中電子也可能通過垂直 d(001)片層的方向傳輸(Bishopetal.,2014)。這種垂直d(001)片層的電子傳遞方式在一些鐵含量較低的蒙脫石礦物中也被發現(Lattaetal.,2017)。ALA作為一種烷基類有機質,其導電能力要遠弱于金屬離子。因此當ALA通過取代了粘土礦物層間的陽離子占據了層間域后,會更加不利于電子在垂直于d(001)片層的方向傳輸。并且ALA插層后粘土礦物的層間距明顯增大,顯然也不利于電子通過層間域在這個方向的傳輸。
另一方面,質子化的ALA還可能取代粘土表面的陽離子而吸附于其表面,還原過程中脫附的ALA也可能通過該方式附著于粘土表面,因此同樣也不利于電子在平行于d(001)的方向上傳播。考慮到質子化的ALA帶正電,而細菌細胞膜帶負電,因此脫附的ALA也可能覆蓋在微生物的表面,從而抑制了微生物的代謝活動,降低其鐵還原的速率。綜合來看,粘土礦物表面和層間吸附的有機質對結構Fe(III)還原程度和還原速率的降低都起著很重要的作用。因此微生物對ALA-NAu-2結構Fe(III)的還原程度和速率都要比NAu-2要低。
3.2 微生物作用下有機粘土礦物的轉變
微生物可以通過破壞礦物結構獲取自身新陳代謝和生長所需的能量和營養物質。因此,礦物在微生物的調控下會發生溶解、沉淀與轉化,在一些條件下甚至會產生一些新的礦物,如白云石、無定形二氧化硅、菱鐵礦甚至一些自然元素等(Konhauser,2007;Dongetal.,2009)。本文實驗結果顯示,高溫菌SA-01在對ALA-NAu-2結構Fe(III)進行還原的過程中,隨著還原程度的升高,有機粘土礦物結構發生溶解,同時生成了一些納米二氧化硅顆粒,而ALA的存在對于次生礦物的產生并沒有明顯影響。
之前有研究表明,粘土礦物層間插層的有機質的種類從某種程度上決定了微生物的還原程度(Zhangetal.,2007)。如果插層的有機質可以作為營養物質被微生物利用或是作為電子穿梭體存在于實驗體系中,將會提升微生物對Fe(III)的還原程度,并且可以促進粘土礦物的轉變。如果插層的有機質對微生物具有毒害作用,比如甲苯,微生物的還原作用將會受到抑制,而粘土礦物也不會發生轉變。本文中所用的有機質ALA既不能被微生物作為營養物質利用,也不能作為電子穿梭體而存在,對微生物也沒有毒害作用,因此微生物鐵還原作用所導致的礦物轉變幾率不大。
3.3 粘土礦物中有機質的釋放機制
綜合分析高溫菌SA-01和超高溫菌G.ahangari實驗組TOC隨時間變化曲線可以看出,有機質的釋放主要受兩個因素的影響,即結構Fe(III)的還原程度和溫度(圖4、圖5)。
溶解態Si、Fe濃度測試結果和還原后 ALA-NAu-2的SEM-EDS譜圖證實微生物在對粘土礦物進行還原時,粘土礦物結構發生還原溶解(表1、圖1b、圖9b),這是一部分有機質從粘土礦物層間釋放出來的主要原因。隨著微生物對有機粘土礦物ALA-NAu-2結構Fe(III)還原程度的升高,少量有機質陸續從粘土礦物的層間釋放出來,當微生物還原停止時,有機質的含量也不再發生變化,二者呈正相關關系。由于微生物還原能力有限(還原程度<30%),總體來說有機質仍會在粘土礦物層間得到很好的保存。并且由于微生物優先還原粘土結構邊部的Fe(III),并且采取從邊緣向中心蔓延的方式進行還原,因此,當微生物還原停止時,位于ALANAu-2中心位置的有機質仍能很穩定的存在于粘土礦物結構中,而邊緣位置由于發生還原溶解,或是還原后產生離子半徑更大的Fe(II)擠壓粘土片層,造成結構的不穩定,該部位的有機質很有可能被釋放出來,層間距減小,恢復到插層前NAu-2的狀態,這就解釋了ALA-NAu-2經SA-01還原后XRD譜圖出現雙峰的原因(圖6a),這與我們之前的研究結果也相似(Zengetal.,2016)。
另一方面,雖然高溫無菌環境也會導致少量ALA自發地從粘土礦物層間釋放出來,例如65℃下時,有機質釋放了0.66%,占總有機碳含量的8.6%;85℃下時,有機質釋放出了1.75%,占總有機碳含量的23.5%。這說明溫度高可能會由于熱力學作用造成一些弱結合態的ALA的脫附。但是在溫度穩定之后,微生物的鐵還原反應對粘土礦物造成的還原性溶解或是結構的畸變仍是ALA釋放的主要原因。因此,針對一些結構鐵含量不高的膨脹性粘土礦物如蒙脫石,微生物鐵還原作用機制弱更弱,那么層間域對有機質保持作用應該更強。
3.4 粘土礦物保存有機質的環境及地球化學意義
盡管眾多的前人研究發現不同類型的礦物對有機質的吸附會有效地降低有機質的礦化速率,從而對有機質起到一定的保存作用(Lützowetal.,2006;Keil and Mayer,2013),但是對于粘土礦物的可膨脹層間域作為有機質保存的場所的相關研究卻甚少。這可能與現今有限的檢測技術手段有所關系。因為對于檢測和表征礦物表面吸附態的有機質較為容易,而對于粘土礦物層間域中的填充有機質的定性定量研究相對較難。例如粘土礦物的d(001)層間距可以根據含水量,層間陽離子類型的不同從幾埃到十幾埃之間發生變化,因此單純依靠XRD確認天然樣品中的層間有機質填充十分困難。并且由于粘土礦物顆粒細小和非均一性較大,一些常規的譜學方法也不容易區分表面、邊部吸附的有機質和填充在層間域中的有機質。雖然高分辨的透射電子顯微鏡已經能夠較為清晰地觀測到粘土的片層結構(Zhaoetal.,2017),但是在電鏡的相關配件上(如X射線能譜、電子能量損失譜)還達不到對有機質進行精細鑒定的程度。因此在天然樣品中粘土礦物層間有機質的定性和定量研究上還存在一定的技術空白。
但是我們通過實驗室的模擬研究發現,一些具有特定官能團的有機質能夠進入粘土礦物的層間域,并且能夠在較高的溫度下和微生物的鐵還原作用中保存下來。這填補了我們在常溫至中溫條件下發現類似現象的空白。對于自然界中是否存在有機質填充在層間域的粘土礦物,我們從理論層面將其擴大到了更廣的溫度范圍,但是未來還需要更多的工作去對天然樣品進行定性定量的表征。
另外粘土礦物的層間域還具有大量的酸性位點,因此被認為是催化有機質生烴的重要場所(Yuanetal.,2013)。而有機質的催化生烴往往需要較高的溫度,我們的結果也證明了即使在高溫條件下有機質也能夠相對有效地保存在粘土礦物的層間域,這為粘土礦物與油氣生成和埋藏之間的關聯進一步提供了理論依據。
4 結論
通過對一種插層態的有機質-粘土礦物復合體的微生物鐵還原結果顯示,溫度和鐵還原作用是控制有機質從粘土礦物層間脫附的主要因素,隨著溫度和鐵還原程度的升高,會有少量有機質從粘土礦物層間脫附出來,但是大部分的有機質仍保存在層間。說明即使在高溫條件下粘土礦物的可膨脹層間域也能夠有效地保存一些具有特殊官能團的有機質。這對極端環境下粘土礦物對有機質的保存作用提供了更多理論依據。
致謝 感謝中國科學院廣州地球化學研究所袁鵬研究員對有機質-粘土礦物復合體合成方面的支持。
謹以此文恭賀葉大年院士八十華誕。
猜你喜歡
科學大眾(2023年17期)2023-10-26 07:39:14
艦船科學技術(2022年8期)2022-06-05 07:36:28
哲學評論(2021年2期)2021-08-22 01:53:34
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:05:56
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中國公路(2017年18期)2018-01-23 03:00:38
數學物理學報(2017年6期)2018-01-22 02:26:40
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現代企業(2015年9期)2015-02-28 18:56:50
