HOXB7 在特發性肺纖維化中抗纖維化的作用
2019-01-19 02:58:54周淼李風雷張海龍孫俊波
實用醫學雜志 2018年24期
周淼 李風雷 張海龍 孫俊波
1河南中醫藥大學第三附屬醫院肺病科(鄭州450000);2河南中醫藥大學第一臨床醫學院(鄭州450000);3河南中醫藥大學第二附屬醫院中醫內科(鄭州450000)
特發性肺纖維化(idiopathic pulmonary fibrosis,IPF)是一種進行性纖維化的間質性肺炎,發病率高,藥物治療手段有限,預后不良[1]。主要病理特點是Ⅱ型肺泡上皮細胞增生,成纖維細胞積聚和肺實質異常重塑[2]。上皮細胞-間充質轉化(EMT)是器官纖維化局灶成纖維細胞的重要來源,有研究表明Ⅱ型肺泡上皮細胞可通過EMT 轉化為成纖維細胞,從而參與肺纖維化的病理過程[3-4]。
同源盒基因HOXB7 屬于HOX 基因家族B 簇,與腫瘤發生發展關系密切[5]。HOXB7 高表達可促進EMT[6],還沒有相關報道證實其在IPF 中Ⅱ型肺泡上皮細胞EMT 中的作用。因此,本研究檢測HOXB7 對Ⅱ型肺泡上皮細胞EMT 的作用,首次揭示其在IPF 中扮演的角色及可能的調控機制。另外,有研究表明堿性成纖維細胞生長因子(basic fibroblast growth factor,bFGF)對EMT 有調節作用且可被HOXB7 調控[7],因此筆者假設HOXB7 可通過調節bFGF 來調節Ⅱ型肺泡上皮細胞EMT 過程,從而調控IPF 的發生、發展。
1 材料與方法
1.1 材料 A549 細胞購自美國ATCC 公司;胎牛血清(FBS)、青霉素、鏈霉素和DMEM 購自美國HyClone 公司;SYBR Premix Ex Taq II 和TRIZOL購自大連寶生物工程有限公司;HOXB7 鼠單抗、bFGF 鼠單抗、E-cadherin 鼠單抗、α-SMA 鼠單抗、COL1A1 鼠單抗、TGF-β1 鼠單抗、GAPDH 鼠單抗和辣根過氧化物酶標記的羊抗鼠二抗購自美國Cell Signaling Technology 公司;BCA 試劑盒購自美國Pierce 公司;Turbofect 購自賽默飛世爾科技公司。
1.2 方法
1.2.1 細胞培養及轉染后形態學觀察 人Ⅱ型肺泡上皮細胞A549 由含10% FBS,2 mmol∕L L-glutamine,100 U∕mL 青霉素和100 μg∕mL 鏈霉素的低糖DMEM 培養基于5%CO2、37 ℃條件下培養,細胞融合度達90%傳代。傳代后細胞生長至70%用無血清培養基培養24 h 使之同步化。隨后進行TGF-β1誘導細胞EMT 過程(將TGF-β1 以5 ng∕mL 的 濃度稀釋于含0.1% BSA 的無血清DMEM 培養基培養48 h)或細胞轉染。細胞分組如下:空白對照(Control組)、TGF-β1 誘導(TGF-β1組)、TGF-β1 誘導48 h 后轉染HOXB7 siRNA(β1-HOXB7 si組)、TGF-β1 誘導48 h 后轉染non-specific siRNA(β1-non-spe組)。最后,使用倒置相差顯微鏡觀察各組細胞形態改變。
1.2.2 組織采集 6例IPF 患者肺組織作為IPF組,其中男5 例,女1 例,年齡45~67 歲,平均58 歲。另外,對照組為因肺部病變行肺葉切除術,肺標本取自遠離病變部位,其中男3 例,女3 例;年齡47~60 歲,平均55 歲。組織立即冷凍。標本均由河南中醫藥大學第三附屬醫院提供,患者簽署了知情同意書。
1.2.3 實時熒光定量PCR(qRT-PCR)檢測[8]TRizol 提取細胞或組織總RNA,利用反轉錄試劑盒以總RNA為模板合成cDNA,而后以cDNA為模板進行qRT-PCR。反應程序:95 ℃4 min;95 ℃10 s,59 ℃30 s,共40 個循環;72 ℃10 min。HOXB7 引物:Sense primer:5′-AAGTTCGGTTTTCGCTCCAGG-3′;Anti-sense primer:5′-ACACCCCGGAGAGGTTCTG-3′。E-cadherin 引物:Sense primer:5′-TTCTTCGGAGGAGAGCGG-3′;Anti-sense primer:5′-CAATTTCATCGGGATTGGC-3′。α-SMA 引物:Sense primer:5′-CTATGCCTCTGGACGCACAAC-3′;Anti-sense primer:5′ - CCCATCAGGCAACTCGTAACTC - 3′ 。COL1A1 引物:Sense primer:5′-AACGAGATCGAG CTCAGAGG -3′;Anti-sense primer:5′-GGGAGG TCTTGGTGGTTTTG-3′。GAPDH引物:Sense primer:5′-CGTCTTCACCACCATGGAGA- 3′ ;Anti-sense primer:5′-CGGCCATCACGCCACAGTTT-3′。反應體系20 μL,包含10 μL SYBR Premix Ex TaqⅡ。GAPDH 為內參,2-ΔΔCt方法計算基因相對表達量。
1.2.4 免疫印記(Western blot) 收集細胞,裂解提取蛋白質,BCA 試劑盒測定濃度且取25 μg 蛋白質進行十二烷基硫酸鈉聚丙烯酰胺凝膠電泳,半干轉法將蛋白質轉移至PVDF 膜。5%脫脂奶粉的Tris Buffered Saline With Tween 溶液室溫封閉膜2.5 h,一抗:HOXB7 鼠單抗(1∶800)、bFGF 鼠單抗(1∶700)、E-cadherin 鼠單抗(1∶500)、α-SMA 鼠單抗(1∶500)、COL1A1 鼠單抗(1∶500)、TGF-β1 鼠單抗(1∶700)和GAPDH 鼠單抗(1∶1 000)4 ℃過夜。辣根過氧化物酶標記的羊抗鼠二抗室溫孵育1 h,暗室曝光觀察結果。GAPDH 為內參對照蛋白。
1.2.5 細胞轉染 將A549(3 × 105)接種于6 孔板,37 ℃,5%CO2條件下培養24 h,參照Turbofect操作說明進行轉染,將HOXB7 siRNA(5′-GCUAUU GUAAGGUCUUUGUTT-3′,5 μg)、non-specific siRNA(5 μg)分別與6 μL Turbofect 混合,分別加入各孔中。5%CO2條件下轉染48 h,后利用qRT-PCR和Western blot 方法檢測轉染效率。
1.3 統計學方法 采用SPSS 22.0 進行統計學分析,數值均采用均數±標準差表示。兩組均數間比較采用t檢驗,多組之間采用單因素方差分析。以P<0.05 為差異有統計學意義。
2 結果
2.1 纖維化組織中HOXB7 表達量升高 為研究纖維化對HOXB7 表達量的影響,利用qRT-PCR 檢測纖維化組織HOXB7 的mRNA 表達量。結果表明,纖維化組織HOXB7 mRNA(圖1)的表達量是正常組織的2 倍,差異具有統計學意義。
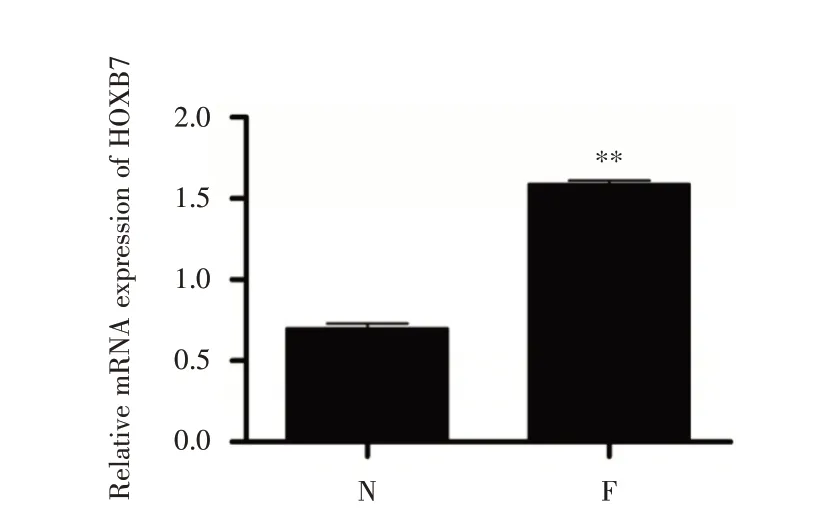
圖1 特發性肺纖維化對HOXB7 表達量的影響Fig.1 The effect of IPF on HOXB7 expression
2.2 抑制HOXB7 表達量 為探究HOXB7 對細胞纖維化的作用,首先通過細胞轉染HOXB7 siRNA抑制HOXB7,qRT-PCR 和Western blot 分別檢測細胞轉染后A549 中HOXB7 mRNA 和蛋白表達量的變化。結果表明,β1-HOXB7 si組HOXB7 mRNA(圖2A)和蛋白表達量(圖2B)較β1-non-spe組顯著下調。
2.3 下調HOXB7 抑制細胞纖維化 在HOXB7 抑制的情況下,通過倒置相差顯微鏡觀察HOXB7 對A549 細胞纖維化的影響。結果表明,TGF-β1組細胞形態與control組相比,細胞由鵝卵石形或多角形轉變為梭形、細胞間連接變松散,呈間充質細胞形態。而β1-HOXB7 si組細胞形態又轉變為鵝卵石形或多角形與control組相同(圖3)。
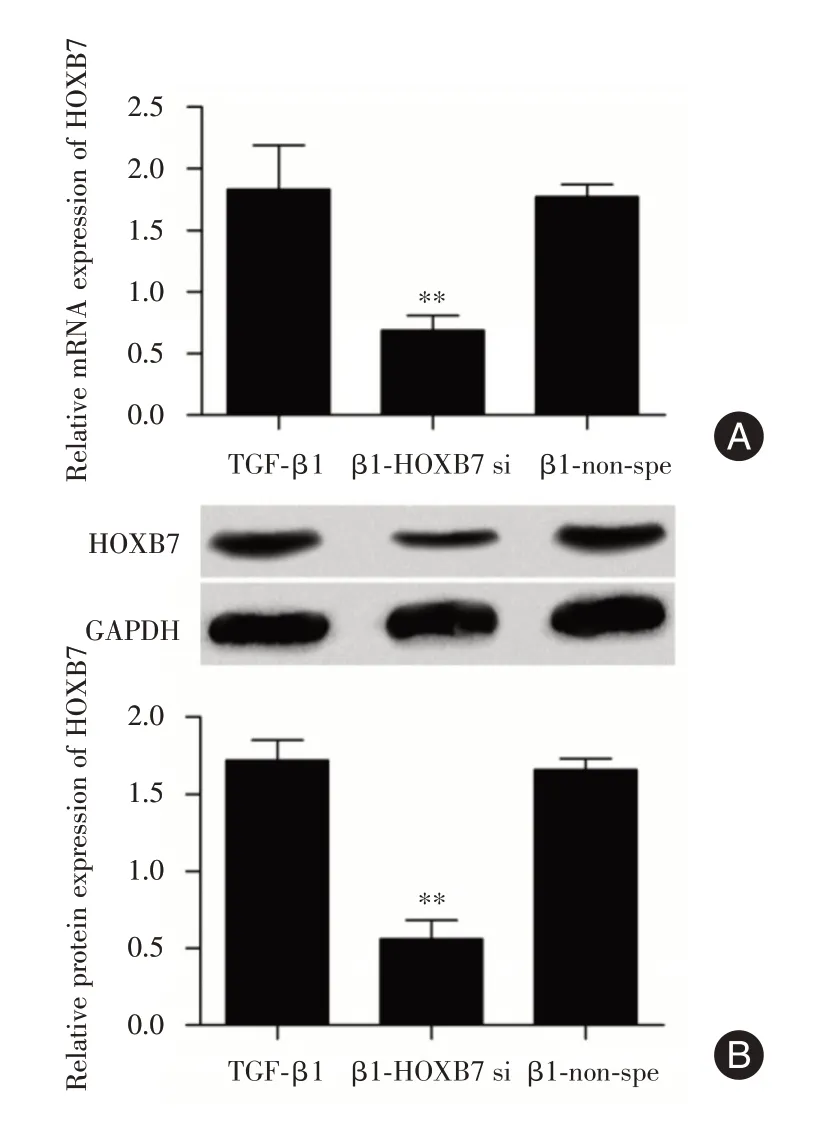
圖2 轉染后A549 中HOXB7 的表達量變化Fig.2 The change of HOXB7 expression in A549 aftertransfection
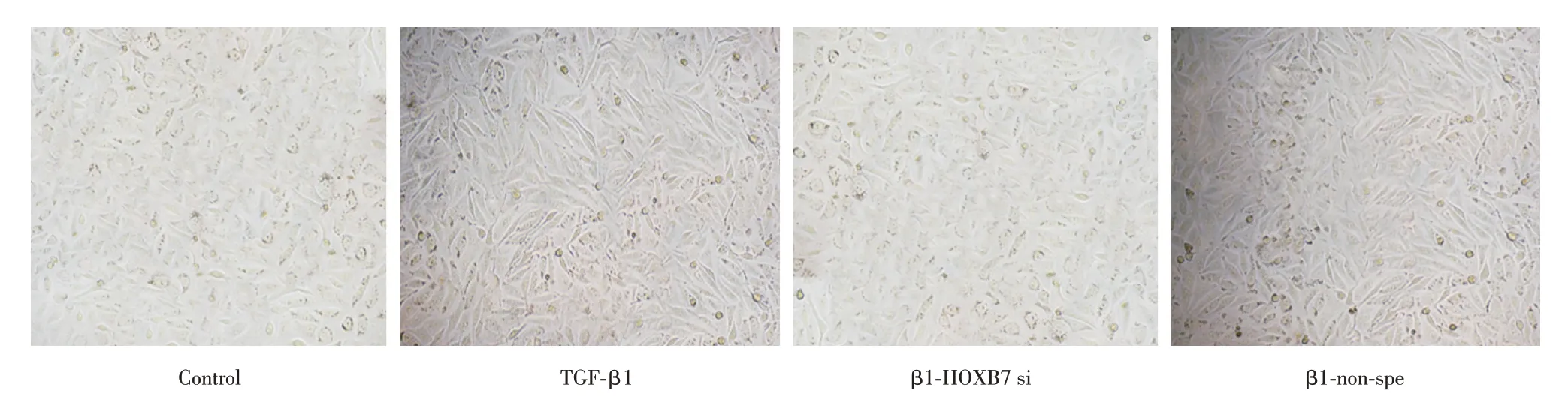
圖3 轉染后A549 形態學變化(×200)Fig.3 Morphological change of A549 after transfection
2.4 下調HOXB7 抑制細胞EMT 過程 為進一步證實HOXB7 對A549 細胞纖維化的作用,檢測了抑制HOXB7 對A549 EMT 的標志物分子E-cad、α-SMA 和纖維化標志物COL1A1 表達量的影響。結果顯示,與β1-non-spe組相比,β1-HOXB7 si組中,E-cadherin的表達量顯著升高,α-SMA 和COL1A1的表達顯著降低(圖4)。
2.5 HOXB7 通過調節bFGF 來調控細胞EMT為探究HOXB7 對細胞纖維化調控的可能機制,利用Western blot 檢測bFGF 蛋白表達量。結果表明,β1-HOXB7 si組中bFGF 的蛋白表達量較β1-non-spe組顯著降低(圖5A)。為了探究HOXB7 對A549 細胞纖維化的調控作用是否可以通過調控bFGF 來實現,用bFGF(9 ng∕mL)孵育β1-HOXB7 si組細胞5 min,發現E-cadherin 蛋白表達量明顯降低,α-SMA 和COL1A1 的蛋白表達量均顯著升高(圖5B)。
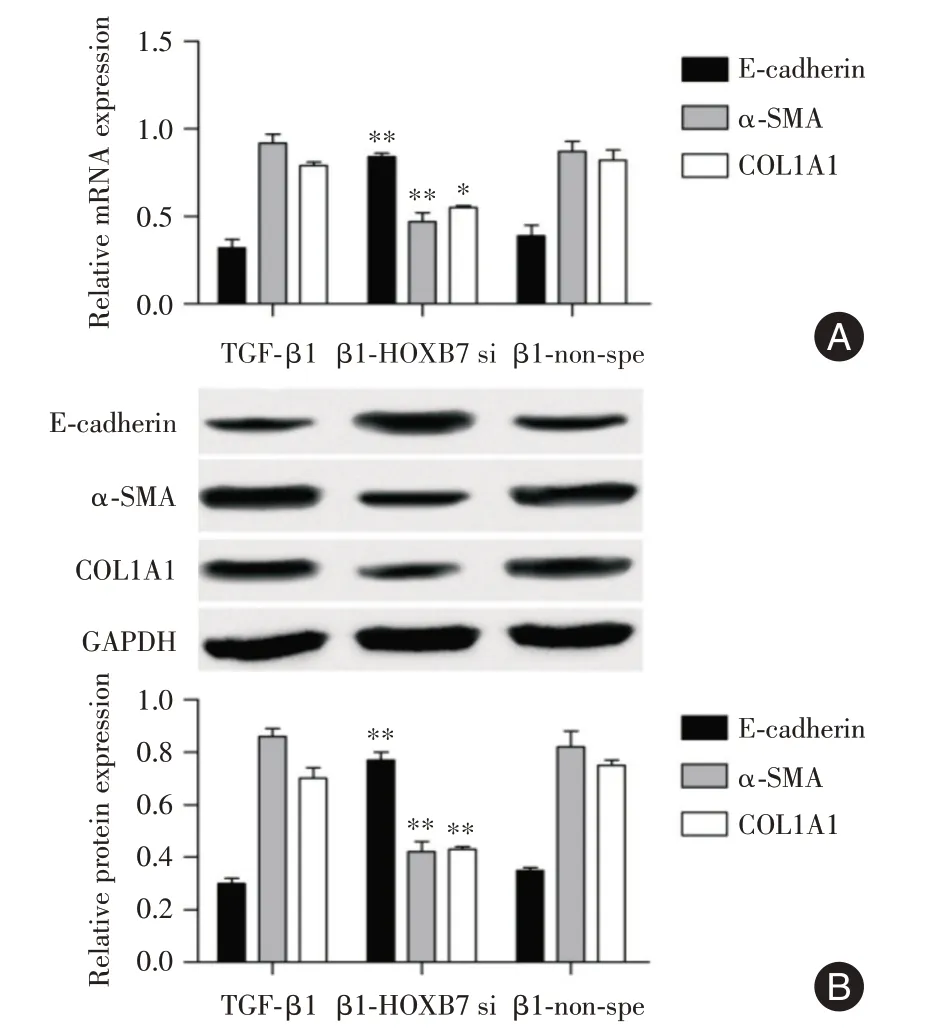
圖4 轉染后A549 中EMT 標志物和COL1A1 的表達量變化Fig.4 Changes in expression levels of EMT markers and COL1A1 in A549 after transfection
3 討論
IPF 是彌漫性間質性肺病中的特殊類型,無特效治療方法,嚴重威脅人類健康。IPF 共同特點是大量成纖維細胞∕肌成纖維細胞聚集和細胞外基質積聚[9]。成纖維細胞和肌成纖維細胞聚集是肺纖維化重要環節,肌成纖維細胞激活可分泌Ⅰ和Ⅱ膠原蛋白,同時促進α-SMA 的表達[10-11]。Ⅱ型肺泡上皮細胞可通過EMT 轉化成肺成纖維細胞∕肌成纖維細胞[12]。在EMT 過程中,上皮細胞通過特定程序轉化為具有間質表型細胞并獲得遷移能力,上皮細胞標記物E-cadherin 等表達下調,間葉標記物α-SMA 等表達上調[13]。因此,本研究選擇E-cadherin 作為上皮細胞標記物,α-SMA 為間葉標記物,可以有效評價EMT 的轉化。Ⅰ型膠原蛋白(COL1A1)的表達也是纖維化的標志[14]。本研究通過檢測Ⅱ型肺泡上皮細胞A549 的EMT 轉化,來評定HOXB7 對肺纖維化的作用。
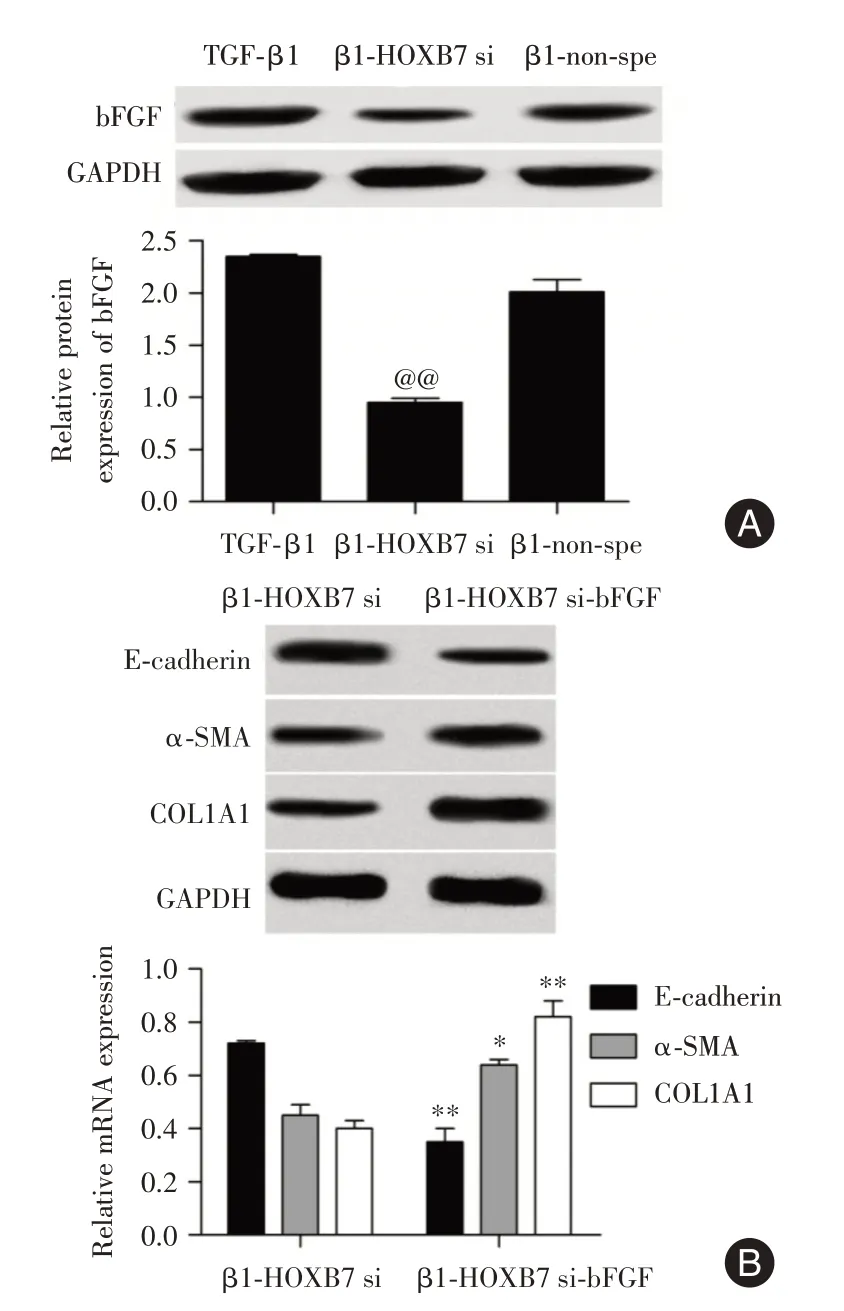
圖5 bFGF 對A549 中EMT 的影響Fig.5 Effect of bFGF on EMT in A549
HOXB7 是胚胎發育時期調控上皮包括肺上皮細胞增殖、分化、遷移的主控基因,而在成體時期,HOXB7 高表達可促進上皮細胞惡性轉化[15]。研究表明,HOXB7 的高表達促進結腸癌、口腔癌、黑色素和肺腺癌等多種癌癥細胞的增殖和EMT[16]。HUAN 等[17]發現,HOXB7 通過誘導細胞EMT 轉化促進乳腺癌細胞遷移。YANG 等[6]發現,HOXB7促進骨肉瘤細胞EMT,從而促進骨肉瘤細胞的增殖和轉移。STIEGELBAUER 等[5]報道HOXB7 被miR-196b-5p 抑制后,結腸癌細胞EMT 同樣被抑制,從而降低結腸癌細胞轉移。然而,對于HOXB7是否對肺纖維化過程中EMT 有影響,目前為止還沒有相關報道。SHIMBORI 等[18]表明,TGF-β1 可顯著誘導A549 細胞纖維化。本研究發現,IPF組織HOXB7 的表達量顯著升高。降低HOXB7 表達量有效抑制TGF-β1 誘導的A549 形態上纖維化轉變且抑制EMT 和COL1A1 的表達,說明A549 纖維化被抑制。因此,本研究首次發現HOXB7 在IPF組織中表達量上調,且干擾HOXB7 表達可抑制TGF-β1 誘導的體外培養A549 細胞的纖維化。為IPF 的預防和治療提供了一定的理論基礎,但本研究側重于對體外培養細胞的研究,所以在后續的實驗中可以側重于體內研究。
有研究[19]表明在口腔癌和乳腺癌細胞中,HOXB7調控bFGF,進而影響癌癥發生、發展。bFGF具有促進EMT 的作用,如在晶狀體上皮細胞、肝細胞癌細胞、前列腺癌細胞中等[7,20]。因此,筆者假設HOXB7 通過調控bFGF 影響A549 細胞纖維化。本研究,結果表明在TGF-β1 誘導細胞的情況下,降低HOXB7 顯著抑制bFGF 表達量,且bFGF 孵育上述抑制HOXB7 的A549 后,顯著消除了抑制HOXB7 對細胞EMT 的作用。因此,減少HOXB7 表達量對TGF-β1 誘導的A549 細胞纖維化的抑制作用,可通過調控bFGF 來實現。
綜上,本研究發現IPF組織中HOXB7 表達量顯著上升,降低HOXB7 顯著抑制TGF-β1 誘導的A549 細胞形態學上纖維化的變化,且抑制EMT 和COL1A1 表達。另外。降低HOXB7 可抑制bFGF的表達,且通過調控bFGF 來抑制TGF-β1 誘導的A549 細胞纖維化,為IPF 的防治提供新的潛在靶標和理論基礎。
猜你喜歡
中老年保健(2022年2期)2022-11-25 23:46:31
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
昆明醫科大學學報(2022年4期)2022-05-23 13:04:50
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
昆明醫科大學學報(2021年4期)2021-07-23 01:21:34
科技傳播(2019年22期)2020-01-14 03:06:54
國際呼吸雜志(2019年21期)2019-11-25 09:52:20
國際呼吸雜志(2019年20期)2019-11-23 08:46:14
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
