稀土尾礦構樹根系形態及代謝研究
2019-01-14 07:39:56郭圣茂吳南生肖永華彭明生
江西農業大學學報 2018年6期
關鍵詞:差異
丁 菲,郭圣茂,吳南生,肖永華,彭明生,蔣 禮
(1.江西農業大學職師院,江西南昌330045;2.江西省寧都縣林業局,江西寧都342800;3.中儲牧草科技有限公司構樹飼料研究中心,河南蘭考475300)
贛南擁有全國30%以上的中重稀土儲備,有著“稀土王國”之稱,但落后的開采工藝剝離了山體表層植被,造成了災難性的生態環境問題,贛南稀土廢棄礦山的治理迫在眉睫。尾礦土壤營養匱乏、持水性差、含砂量高的極端環境特點使植物很難在此存活,土壤修復極其困難,人工干預才能加快植被恢復進程[1]。構樹(Broussonetiapapyrifera)是桑科構樹屬落葉大喬木,適應性特別強,民間稱其為“惡樹”,近年來逐漸受到眾多研究者的關注。一些學者應用構樹對銻礦、采石礦、鉛鋅礦、菱鎂礦等礦區進行植被修復,認為其是礦區廢棄地生態恢復的適生先鋒樹種[2-3],本課題組前期研究工作也證明構樹對贛南稀土尾礦具有較強的耐受性[4]。文獻表明,關于構樹對礦區的適應性和對旱鹽、低溫、重金屬等脅迫的抗逆性研究主要反映在地上部生長和生理生化響應[5-7],對構樹地下生態系統與抗逆關系的研究極少。根系是地下生態系統中進行物質和能量交換的主要器官,由于直接與土壤接觸,更容易對土壤環境做出反應[8],受生境差異影響,植物根系會在空間分布、形態特征、生理特性上表現出不同的生態適應對策。許多學者研究了根系形態對逆境的響應,Hund等[9]和李文饒等[10]認為干旱脅迫下,根系生長受到抑制,根系生物量、根冠比、根長、根系表面積和體積等發生變化以適應干旱脅迫,王樹鳳等[11]研究發現Cd脅迫抑制了杞柳根的伸長,促進了根的徑向生長,導致根系平均直徑增加。根系適應逆境的生理響應報道也較多[12],王亞男等[13]認為鳶尾根系代謝分析有利于更好地理解植物對石油烴污染土壤的代謝響應并揭示其修復機制。代謝組學是在現代高通量檢測技術平臺不斷發展的基礎上,以生物體內小分子代謝物分析與數據統計分析為基礎,通過生物體內代謝物的種類、數量及其代謝途徑的變化來研究生物體的代謝過程,已經發展成為探索植物對遺傳和環境的改變所做的系統響應的重要工具[14-15]。
綜上,構樹有望成為礦區等困難立地生態重建的先鋒樹種,然而構樹根系在生態恢復中的作用及其地下關鍵生態過程對礦區干旱、貧瘠逆境的響應機制長期被忽視,從地下生態系統角度進一步探討構樹的逆境適應策略具有重要的理論和現實意義。本研究通過分析構樹根系的形態特征,應用代謝組學研究根系代謝產物的變化特點,擬揭示構樹適應稀土尾礦土壤環境的內在機制,發展和完善構樹抗性理論,為尾礦廢棄地的生態恢復提供理論支撐。
1 材料與方法
1.1 研究區概況
試驗地點位于江西省贛南地區寧都縣東山壩鎮,東經116°31′,北緯26°44′,地處中亞熱帶丘陵山區季風濕潤氣候區,四季分明,氣候溫和,平均氣溫在攝氏19.8 ℃度左右,全年無霜期近11個月,年降雨量1 600 mm左右,多集中在4—6月份,約占總降雨量的50%。土壤為稀土礦開采4年后的廢棄殘礦尾砂,開采后的山頭有效土層和風化層基本剝離,水土流失嚴重,使得土壤沙化,沙礫含量高,保水性能非常差,養分含量極低,寸草不生。由于稀土洗礦工藝和水土流失造成試驗地溝蝕嚴重,在造林之前用挖機對廢棄尾礦地進行了充分平整,2015年3月,選擇長勢基本一致的1 a構樹實生苗對稀土尾礦進行造林,平均株高51 cm,平均地徑1.1 cm,造林密度為1.0 m×0.5 m,同時在同一山頭周邊未開采的土壤種植構樹作為對照。
1.2 研究方法
1.2.1 根系形態研究采樣 2017年7月設置試驗樣地(稀土尾礦區,以Y表示)和對照樣地(周邊未開采的坡地,以CK表示)各4塊,樣地面積為10 m×20 m,每塊樣地各選4株標準木共16株,采用挖掘法,先挖開幼苗周邊表層土,再沿著根系分布小心挖掘,同時用水沖洗根部,使其露出土壤,以保護側根不脫落。選取根系完整的植株裝入塑料袋,編號裝入密封的塑料袋,帶回實驗室測量根系形態和生物量[16]。將每株主根上的全部側根剪下,用Epson Expression 00XL掃描儀獲取根系掃描圖像,用Win RHIZO根系分析軟件(加拿大Regent公司)自動統計不同直徑總根長度、總根表面積,掃描后的根系樣品置入烘箱80 ℃烘干24 h至恒質量,再分別稱質量和記錄。比根長(specificrootlength,SRL)為根系總根長與根系生物量之比,比表面積(specificrootarea,SRA)為根表面積與根系生物量之比。
1.2.2 代謝物分析取樣和檢測 在試驗和對照樣地中分別隨機挖取30株側根根系,5株混合為一個生物學樣品,每組6個重復,立即用液氮速凍用于代謝組分析。取樣品100 mg用液氮研磨,取100 μL樣本,加入300 μL甲醇和20 μL內標,渦旋混勻30 s后,冰水浴中超聲提取5 min,-20 ℃靜置2 h,4 ℃ 13 000 rpm離心15 min,取200 μL上清液于2 mL進樣瓶,進行LC-MS分析。質控樣本(QC)由實驗樣本提取物等量混合制備而成,用于分析樣本在相同的處理方法下的重復性。在儀器分析的過程中,每6~10個檢測分析樣本中插入一個質控樣本,以監測分析過程的重復性。LC-MS分析的儀器平臺由Agilent1290超高效液相色譜串聯Thermo Fisher Scientific 的Q Exactive Orbitrap高分辨質譜儀組成。所用色譜柱為UPLC HSS T3色譜柱(1.7 μm 2.1×100 mm,Waters)。正模式流動相為0.1%甲酸水溶液和乙腈,負模式流動相為5 mM醋酸銨水溶液(用氨水調節pH值至9.00)和乙腈,采用Q Exactive Orbitrap高分辨質譜儀進行一級、二級質譜數據的采集。
1.2.3 數據處理 根系形態所有數據利用Microsoft Excel和SPSS軟件進行處理,同一徑級不同處理之間、同一處理不同徑級之間根長、根表面積、生物量、SRL、SRA的差異采用獨立樣本T檢驗。顯著性水平設定為P=0.05。對質譜分析后得到的原始數據進行基線過濾、峰識別、積分、保留時間校正、峰對齊和歸一化,最終得到一個保留時間、質荷比和峰強度的數據矩陣,利用歸一化后的數據矩陣進行多元統計分析,結合OPLS-DA(正交偏最小二乘法判別分析)的VIP值和單變量統計分析T檢驗P值來篩選顯著差異代謝物并畫熱圖。
2 結果與分析
2.1 構樹修復稀土尾礦的根系形態響應
2.1.1 細根根長、表面積和生物量 從圖1可以看出,與周邊未開采礦區構樹相比,稀土尾礦區構樹細根總長下降了32.39%、總表面積下降了21.69%,根系生物量下降了52.6%。除>2 mm徑級外,同一徑級根系長度、表面積和生物量均呈顯著減少。進一步分析表明,尾礦區構樹<1 mm徑級的根長占總根長的84.01%,比對照區同徑級根長占比增加10.05%,而>2 mm細根長度所占整個根系的百分比例呈減少的趨勢。尾礦區構樹根系<1 mm徑級的表面積占比也出現類似情況。
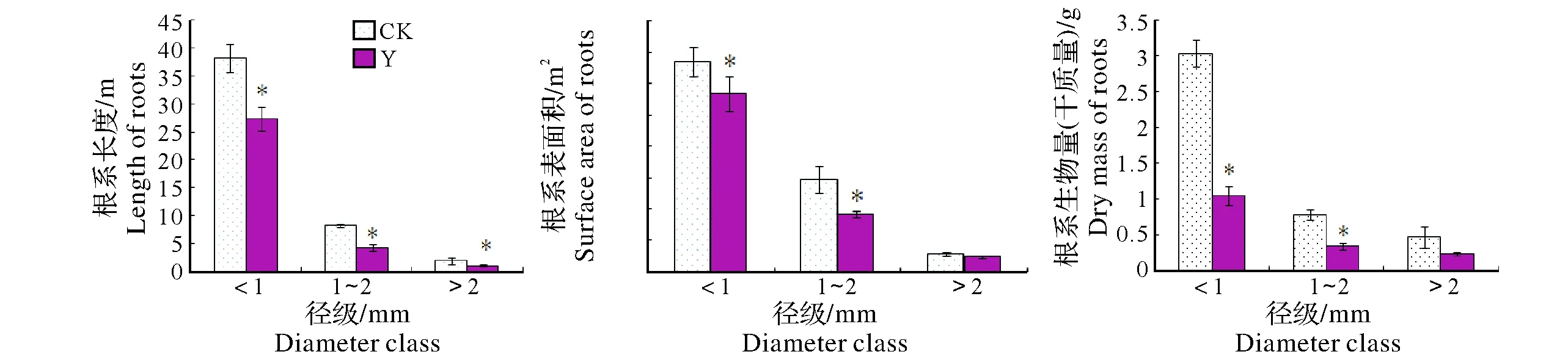
Y,尾礦區Rare earth tailing area;CK,周邊未采礦區unmined area around earth tailing柱形圖上方*的差異表示在5%水平上差異顯著Values in each column followed by the * are significantly different at 0.05 level according to LSD圖1 構樹不同徑級根系長度、表面積和生物量Fig.1 Root length,surface area and dry mass of different diameter class in B. papyrifera
2.1.2 比根長、比表面積 由圖2可知,尾礦構樹總的比根長表現出增加的變化趨勢,其中?<1 mm的比根長比對照增加109.58%,1 mm
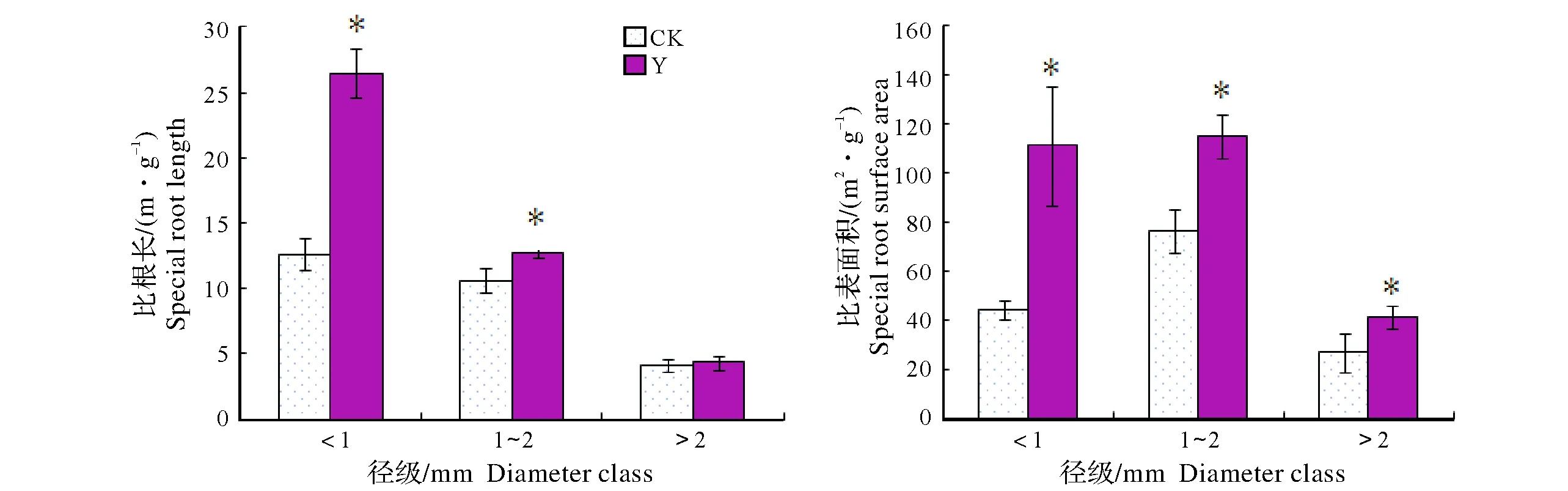
圖2 構樹不同徑級根系比根長、比表面積Fig.2 Special root length and Specific surface area of different diameter class in B. papyrifera
2.2 構樹修復稀土尾礦的根系代謝分析
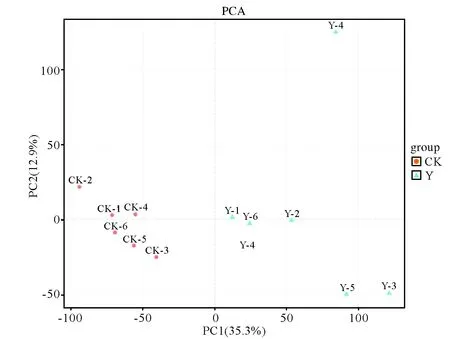
圖3 構樹處理(Y)與對照樣品(CK)的PCA得分圖Fig.3 PCA scores of treated(Y) and control (CK) in B. papyrifera
2.2.1 構樹修復稀土尾礦的根系PCA和離子流分析 從圖3的PCA得分圖可以看出,兩組樣品6個重復的數據點檢測結果都分別集中在一起,說明整個分析測試過程中系統誤差(實驗操作、儀器穩定性)較小,數據可重復性好。另外,兩組樣品的數據點在空間上可以明顯區分,說明它們的代謝產物在數量、種類和濃度等方面存在差異,這種差異在總離子流圖(圖4)中也得到充分體現:如圖4,檢測到2.8~3.5 min,3.6~4.2 min,6.7~8.2 min幾個階段稀土尾礦構樹根系中大量小分子物質顯著上調,而在8.2~9.2 min,10.5~11.2 min階段許多代謝物質與對照相比呈顯著下調趨勢。經過LC-MS分析,在處理組(Y)和對照組(CK)樣品中共檢測出1 968種代謝物存在顯著差異(VIP≥1且T-testP<0.05),其中上調物質1 812種,下調物質156種。
2.2.2 稀土尾礦構樹根系差異代謝產物分析 如圖5所示,與周邊未采礦坡地構樹相比,稀土尾礦構樹根系代謝物中,亞油酸、蘋果酸、木二糖、葡萄糖、葉黃素、松三糖、半乳糖醇、色氨酸、酪氨酸、D-泛酸、葫蘆巴堿、乳糖、麥芽酚、白藜蘆醇等38種物質含量顯著增加,而減少了絲氨酸、十六烷酰胺、硬脂酰胺等8種物質的含量,大部分的代謝物是在初生代謝途徑中產生的,包括有機酸、氨基酸、糖類及其它組分。
為了進一步分析這些差異物質在稀土尾礦構樹根系代謝中的貢獻,對VIP值進行計算,如圖6所示,發現葫蘆巴堿、棕櫚酸、谷胱甘肽醇VIP>10,其次為海藻糖、3-羥基-L-谷氨酸、亞油酸、麥芽糖、色氨酸、1,7-二甲基黃嘌呤、3-脫氫莽草酸酯等,它們在應對稀土尾礦環境中起到了實質性的作用。
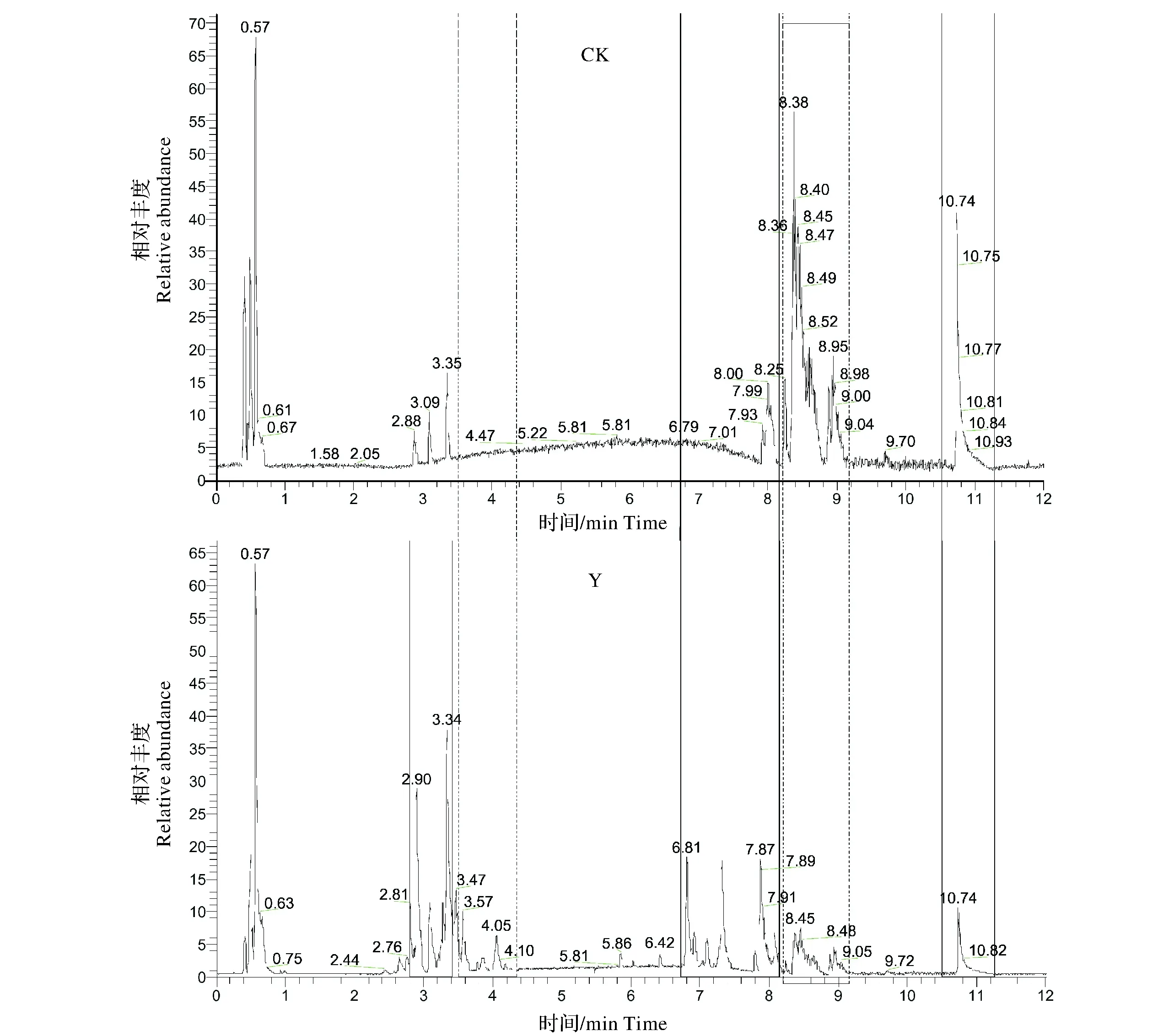
圖4 構樹處理(Y)與對照樣品(CK)的總離子流圖Fig.4 Total ion chromatography of treated(Y) and control(CK) in B. papyrifera
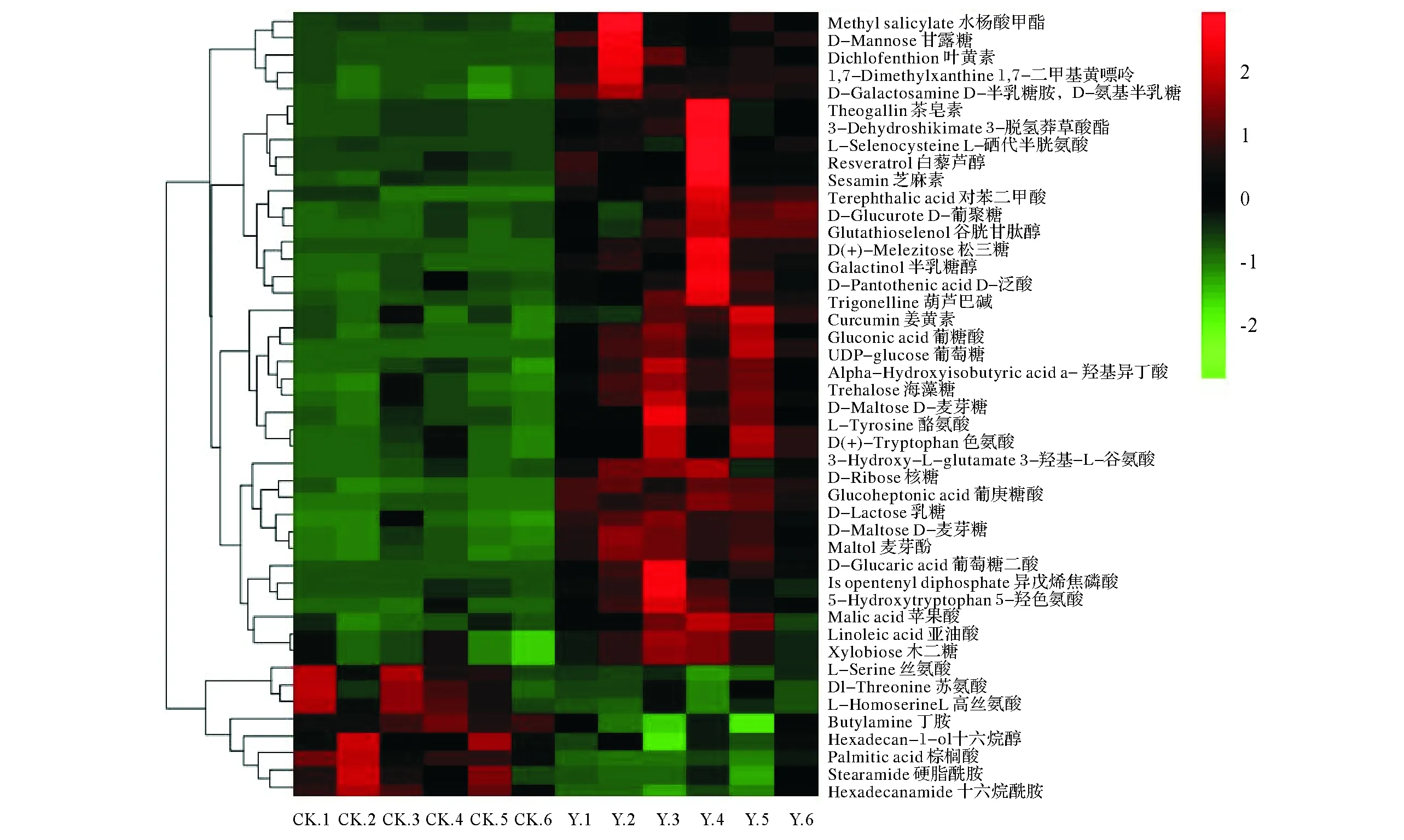
圖5 構樹處理(Y)與對照樣品(CK)的差異代謝物熱圖Fig.5 Differential metabolite Heatmap of treated (Y) and control (CK) in B. papyrifera

圖6 構樹根系差異代謝物的VIP值Fig.6 VIP of different metabolites in B. papyrifera
3 討論與結論
3.1 稀土尾礦構樹根系形態適應特征
根系生長關系到植物的水分和養分吸收,遭受脅迫時可以通過形態、生理方面的變化來適應環境[17-18],表征根系形態特征的參數主要有根系總長度、表面積、生物量、平均直徑、比根長、比根面積等。本試驗地土壤沙化干旱、營養匱乏,是稀土尾礦植被恢復的主要障礙因子[19]。研究發現,與周邊未開采礦區構樹相比,稀土尾礦土壤環境使構樹根系的形態特征發生了變化,但各形態指標變化不一致。
一方面,側根長度、表面積和生物量明顯減少,可能主要是由于尾礦土壤貧瘠、含水量低,限制了根系的生長,這與Pregitzer等[20]研究結論一致。但是低序級側根根長和表面積占比提高。研究表明,水和無機礦物質的吸收通過較細的根進行[21-22],直徑越小的根吸收養分和水分的能力越強[23]。吳敏等[24]發現,栓皮櫟幼苗的小細根可以幫助根系在土壤中占據更多的空間位置,增強根系對周圍水分的吸收能力。因此,構樹在尾礦干旱貧瘠的環境下根系形態的這種變化可以擴大橫向吸收水分和養分的面積,是根系對資源匱乏生境的生態適應。
另一方面,尾礦構樹總比根長、總比表面積比對照顯著增加,小徑級根系比根長和比表面積增加幅度較大且差異顯著。比根長主要表征植物根系收益和投入的關系,比根長增加,單位質量的根系具有更大的吸收表面積,土壤資源的獲得效率更高。武高林等[25]對4種鳳毛菊屬(Saussurea)植物研究后發現,植物在惡劣條件下能夠通過增加比根長來更有效地吸收土壤中的養分。比表面積反映植物根系對土壤資源獲取的能力[26],Bauhus等[27]認為,SRL和SRA決定了根系吸收水分和養分的能力,其值越大,表明投入相同的成本有較高的養分利用效率。本研究SRL和SRA的變化表明構樹根系能夠以較少的投入獲得更多的收益,水分和養分利用效率較高,說明構樹不是被動忍受而是主動調節其根系生長以提高吸收效率減緩逆境傷害。
綜合前述,應該是由于稀土尾礦嚴重缺水缺肥的惡劣土壤環境條件誘導構樹產生了更多數量、更小直徑的細根,而構建和維持單位長度細根消耗的碳水化合物較少,有利于行使細根的吸收功能[28],這可能是構樹應對稀土尾礦環境的根系調整策略。
3.2 稀土尾礦構樹根系代謝響應
植物對非生物脅迫的耐受性與敏感性是一個非常復雜的生命過程,大量物質含量改變,積累滲透保護劑、調整啟動抗氧化酶保護系統、激活相關保護性的代謝途徑以重構代謝平衡[29]。本研究中檢測到大量差異代謝物質,主要包括氨基酸、糖類、有機酸等。
氨基酸在植物應對逆境生理中扮演著重要的角色,大量研究證明,非生物脅迫下植物體內游離氨基酸呈積累趨勢[30],它們應對脅迫的重要職能是參與滲透調節[31-32],其中某些特定氨基酸的積累則可能預示著植物耐脅迫能力得到增強的積極信號[33]。本研究中分析表明稀土尾礦構樹根系色氨酸、酪氨酸顯著上調,分別富集到多種氨基酸代謝通路,參與了應對環境壓力的調節,與前人的研究結論類似。
糖為植物生長發育和代謝過程提供碳和能量,增強植物抗逆性,在進化早期即獲得抵抗脅迫重要調節功能。本研究發現,構樹在稀土尾礦環境壓力下,松三糖、核糖、麥芽糖、海藻糖、木二糖、葡萄糖、甘露糖含量顯著增加,它們可能參與滲透調節,增強了構樹適應稀土尾礦環境的能力。其中海藻糖VIP值達7.8,在所有糖類差異代謝物中表現最為突出。海藻糖不同于其它糖類,它對生物體具有神奇的保護作用,是應激代謝的重要產物。它能夠保護生物細胞和生物活性物質在脫水、干旱、高溫、冷凍、高滲透壓及有毒試劑等不良環境條件下活性免遭破壞。許多對外界惡劣環境表現出非凡抗逆耐受力的物種,都與它們體內存在大量的海藻糖有直接的關系[34]。海藻糖與構樹抗逆性的關系值得進一步深入研究。
在構樹根系差異代謝物中,葫蘆巴堿( trigonel-line)的VIP值最大。葫蘆巴堿是一種嘧啶甜菜堿,是細胞質中的親和性溶質,是一種較好的滲透調節物質,對細胞膜的穩定性起到重要的保護作用[35]。Cho等[36]對17個大豆品種進行鹽脅迫處理,其中抗鹽品種葉片中葫蘆巴堿含量增加1.5倍以上。除此之外,葫蘆巴堿還被認為是一種天然的植物激素而引起生物學者的關注,因為其能使植物根尖組織細胞周期的G2期停止或減慢G2期,Mazzuca等[37]以萵苣(LactucasativaL.)的根尖組織作為試驗材料,用3mmol/L葫蘆巴堿處理根尖組織,減小了原生根系的伸長速率。根據前述根系形態的變化,推測葫蘆巴堿可能影響了稀土尾礦構樹根系的生長和形態特征。所以,類似于前人的研究成果,葫蘆巴堿作為構樹根系代謝的非常重要的差異代謝物對稀土尾礦土壤環境做出響應,可能一方面作為滲透調節物質保護著細胞膜,另一方面調控著根系的生長。
當植物處于逆境條件時,體內的活性氧產生和清除的動態平衡遭到破壞,近年來人們對于植物活性氧氧化傷害機制和其防御體系做了大量研究,發現抗壞血酸、類胡蘿卜素等許多抗氧化物質可以通過多條途徑直接或間接地猝滅活性氧[38]。本研究表明,稀土尾礦構樹根系中3-脫氫莽草酸酯、谷胱甘肽、泛酸、白藜蘆醇、芝麻素、胡蘿卜素等差異代謝物顯著上調,研究表明,3-脫氫莽草酸酯是一種十分有效的抗氧化劑,其活性優于沒食子酸、丙基沒食子酸[39];泛酸在體內主要以CoA形式參與糖、脂、蛋白質代謝,具有抗脂質過氧化作用,保護細胞質膜不受損害;GSH不僅可以作為GR的底物通過Halliwell-Asada途徑清除H2O2,也可以作為抗氧化劑直接清除活性氧,是一類重要的抗氧化物質;白藜蘆醇、胡蘿卜素也是強抗氧化劑[40],可見,這些差異代謝物都是重要的活性氧清除劑和抗氧化劑。
本研究分析了稀土尾礦構樹的根系形態和根系代謝響應,結果表明?<1 mm的細根比例和細根表面積占比顯著提高,總比根長、總比表面積顯著增加且小徑級根系SRL和SRA增加幅度較大,這種形態上的主動調節可以減少生長消耗,提高水分和養分吸收效率;根系代謝組學分析表明滲透調節作用和抗氧化作用是稀土尾礦構樹適應稀土尾礦環境的主要途徑,根系色氨酸、酪氨酸、蘇氨酸、絲氨酸等氨基酸物質,松三糖、核糖、麥芽糖、海藻糖、木二糖、葡萄糖、甘露糖等糖類物質及葫蘆巴堿等差異代謝物是重要的滲透調節物質,3-脫氫莽草酸酯、谷胱甘肽、泛酸、白藜蘆醇、芝麻素、胡蘿卜素等差異代謝物顯著上調,作為抗氧化物質,增強了構樹應對環境壓力的能力。根系形態上的調整和根系代謝的差異是構樹適應稀土尾礦環境的重要機制。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
