厚竹高生長期莖稈節部對基本組織細胞壁發育形成的影響
2019-01-14 07:39:50陳阿麗
江西農業大學學報 2018年6期
關鍵詞:生長
夏 敏,陳阿麗,于 芬
(江西農業大學林學院/江西省竹子種質資源與利用重點實驗室,江西南昌330045)
高生長是竹類植物生長發育的關鍵過程,與生物量密切相關。一些竹種在竹筍出土初期每天僅長高幾厘米,以后逐漸加快,每天幾十厘米,在高生長旺盛期一晝夜的高生長量可達1 m[1],并且在45~60 d內即可完成稈型的生長,最大生長高度可達15~20 m[2]。而細胞壁是植物細胞的重要結構,其結構成分和狀態與植物細胞的伸長密切相關,植物細胞的伸長直接影響植物的個體生長。因此探究細胞壁的發育規律對揭示竹稈節間快速高生長的機制有重要意義。

圖1 厚竹莖稈正常節和異常節外部形態Fig.1 External morphological maps of normal and abnormal sections of thick bamboo stems
厚竹(Phyllostachysedulis‘Pachyloen’)是毛竹(Phyllostachysedulis)的一種優良的變異品種,其不僅具有稈壁特厚、上部近實心、生物量大等特點,同時因其優良的竹材沖擊特性和纖維性狀等特征,使其具有重要的種質研究意義和綜合開發利用價值[3-6]。在對厚竹進行研究的過程中發現部分厚竹具有節部異常的現象,異常節和正常節相比,在外部形態上主要表現為節間變短(圖1)。竹稈的迅速高生長主要依賴于節部的居間分生組織。竹稈節部在竹子的生長發育過程中不僅具有機械支撐作用,而且物質的橫向交流也主要發生在節部。竹子的高生長也與竹稈的節部密切相關。節間快速伸長主要是由細胞伸長引起的,細胞伸長與細胞壁結構和物質的變化息息相關。因此猜想,節部異常會不會通過影響細胞伸長過程中細胞壁結構和物質的變化進而影響節間的伸長,從而在外部形態上表現為節間縮短呢?目前,有關厚竹莖稈快速高生長過程中基本組織細胞壁的構建機制及節部對莖稈快速高生長的影響機理還不明確。因此,本文擬通過對比研究節部異常和節部正常的竹子高生長過程中細胞壁的發育對厚竹莖稈快速高生長過程中異常節和正常節節間中部基本組織細胞進行比較發育解剖學研究,了解基本組織細胞壁構建的細胞學基礎,探究節部對高生長的影響,為進一步揭示莖稈快速高生長提供理論依據。
1 材料與方法
1.1 試驗材料
1.1.1 光學材料 實驗材料取自于江西農業大學竹類植物種質園厚竹實驗基地。分別于2016年4月12日、2016年4月16日、2016年4月18日采取不同發育階段節部正常和異常的厚竹筍5株,筍高分別為0.78、1.10、1.30、1.37、1.69 m。要求生長狀態良好、生境相同、無病蟲害。節數從基部開始算起,連續截取各個節間中部和節部的材料。
1.1.2 電鏡材料 實驗材料厚竹于2016年4月18日采自于江西農業大學竹類植物種質園厚竹實驗基地。選取3株高1 m左右的竹筍作為樣株,采取筍尖,然后從基部往上開始的各節間中部取1 mm3左右的小塊。
1.2 方法
1.2.1 光學制片 材料選取后立即投入FAA固定液中固定,成熟材料用70%FAA,幼嫩材料用50%FAA,且幼嫩材料用酒精梯度進行脫水,二甲苯透明,常規石蠟切片法制作并于旋轉切片機上做連續厚約8~10 μm的橫、縱切片,番紅-固綠對染,二甲苯透明,中性樹膠封片,Zeiss AX10熒光顯微鏡下拍照。
1.2.2 透射電鏡制片 材料選取后立即用2.5%的戊二醛和4%的多聚甲醛初固定,1%的鋨酸后固定,磷酸緩沖液沖洗,酒精梯度脫水,spurr樹脂包埋,于LKB-V型超薄切片機切片,H-7650型透射電鏡觀察拍照。
2 結果與分析
根據在光鏡下觀察的組織結構特征,將基本組織細胞的分化與發育過程分為兩個時期:初生壁形成期、次生壁形成期。
2.1 初生壁形成期
該時期主要分為兩個階段,初生壁發育形成初期和初生壁發育形成后期。
從顯微結構分析,初期,正常節和異常節基本組織并沒有明顯的差異。該時期主要以細胞分裂為主,體現為細胞數量的增加(圖2),細胞長度變化不明顯(圖3,注:24、15、7是異常節)。從竹筍的縱切面上觀察,頂端分生組織具有原套-原體結構(圖4,A)和節部與節間分化(圖4,B)。細胞排列整齊,細胞核較大且居中,部分細胞正處在有絲分裂狀態(圖4,C)。部分基本組織細胞和纖維細胞中能觀察到雙核仁的現象(圖4,D)。細胞中含有大量的淀粉粒(圖4,E),節部淀粉粒含量較節間多(圖4,F)。
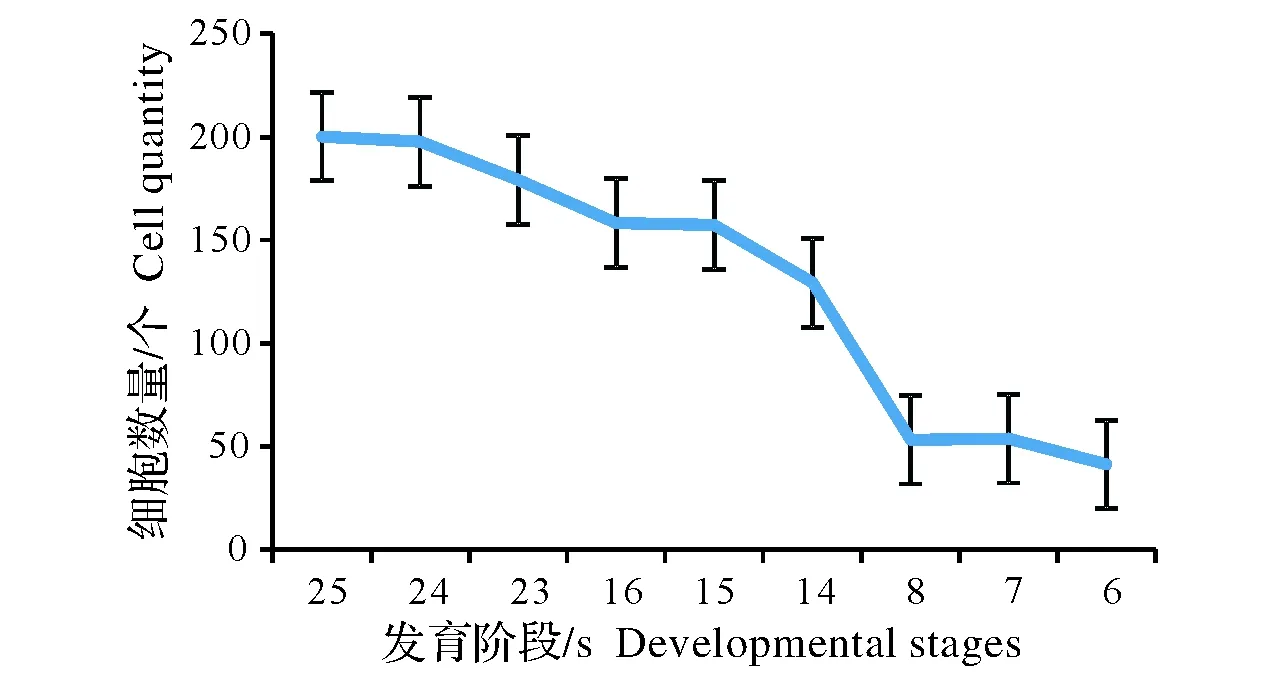
圖2 不同發育階段基本組織細胞數量的變化Fig.2 The varying of ground tissue cell quantity in different developmental stages

圖3 不同發育階段基本組織細胞長度的變化Fig.3 The varying of ground tissue cell length in different developmental stages
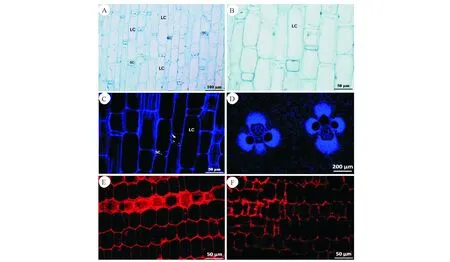
后期,主要以細胞伸長為主,節部異常引起細胞數量的增加和細胞長度的減小(圖2,圖3)。隨著組織的發育,正常節長、短細胞的分化較明顯(圖5,A)。異常節較正常節而言長、短細胞分化不明顯(圖5,B)。兩者細胞壁在熒光顯微鏡下觀察均有增厚的現象(圖5,C、D)。淀粉粒含量節部異常節較正常節多(圖5,E、F)。
從超微結構分析,初期,基本組織細胞壁較薄,胞間連絲豐富,細胞中含有大量細胞器和運輸小泡(圖6,A)。節部異常節較正常節線粒體內嵴狀結構有降解的現象(圖6,B、C)。
后期,隨著組織的發育,基本組織細胞壁加厚明顯,細胞器豐富(圖6,D)。節部正常的節基本組織細胞壁較節部異常節的細胞壁加厚明顯,胞間連絲較豐富,物質運輸小泡仍較多(圖6,D、E、F)。
2.2 次生壁形成期
從顯微結構分析,無論是正常節還是節部異常節,都有長、短細胞的分化且分化明顯,短細胞細胞核邊緣化分布(圖7,A),長細胞的內容物多數已降解 (圖7,B)。長細胞次生加厚明顯(圖7,C、D)。只有極少數細胞中還有零星的淀粉粒分布(圖7,E、F)。
從超微結構分析,長細胞壁次生加厚明顯,能觀察到細胞壁不同的壁層,微管沿質膜分布于細胞壁的內側(圖6,G)。節部異常節較正常節而言,胞間層含有電子密度較低的白色物質(圖6,H),細胞壁上有白色的凸起物質(圖6,I)。
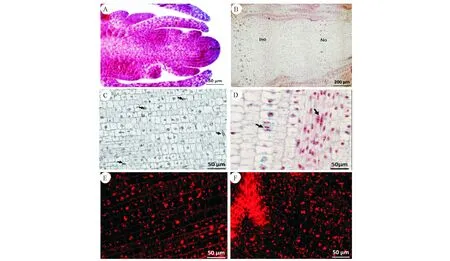
A:頂端分生組織;B:節間與節部基本組織的分化;C:部分基本組織細胞正在進行有絲分裂;D:基本組織和纖維細胞具有雙核仁現象;E:基本組織細胞內含有較多的淀粉粒;F:節部淀粉粒的分布A:Apical meristem;B:Differentiation of ground tissue between internodes and nodes;C:Some groung tissue cells were undergoing mitosis;D:The groung tissue and the fiber cells have the double nucleoli;E:More starch grains were found in the ground tissue cells;F:The distribution of starch grains in the node圖4 初生壁形成初期基本組織解剖結構Fig.4 Anatomical structure of ground tissue in early stage of primary wall formation

A:長、短細胞分化明顯;B:長細胞細胞核邊緣化分布;C:基本組織細胞壁加厚;D:節部異常節基本組織細胞壁加厚;E:基本組織細胞內淀粉粒減少;F:節部異常節基本組織細胞內淀粉粒含量較多.注:B、D、F 節部異常節A:The differentiation between long and short parenchyma cells was obvious;B:Marginal distribution of long cell nucleus;C:Thickening of ground tissue Cell walls;D:The cell wall was thickened in the ground tissue cell walls of abnormal node;E:Starch grains decreased in the ground tissue cells;F:The starch grains in the ground tissue cells of abnormal node were higher.Note:B、D、F abnormal node圖5 初生壁形成后期基本組織解剖結構Fig.5 Anatomical structure of ground tissue in the late stage of primary wall formation

A:初生壁形成初期基本組織細胞壁較薄;B:胞間連絲豐富,細胞器較多;C:節部異常節線粒體結構;D:正常節胞間連絲豐富;E:初生壁形成后期基本組織細胞壁加厚明顯;F:節部異常節運輸小泡較少;G:基本組織細胞壁次生加厚;H:基本組織細胞壁上電子密度較低的白色凸起物質(如箭頭所示);I:基本組織細胞壁胞間層電子密度較低的白色物質(如箭頭所示).C、F、I 節部異常節;A、B、C 初生壁形成初期;D、E、F 初生壁形成后期;G、H、I 次生壁形成期A:The cell walls were thin in the early stage of primary wall formation;B:Rich plasmodesmatas and organelles;C:The mitochondrial structure of the abnormal node;D:The plasmodesmatas in normal node were rich;E:The cell walls were thickened obviously in the late stage of primary wall formation;F:The transport vesicles in abnormal node decreased;G:The long parenchyma cell walls were secondary thickening;H.White protruding material with low electron density on the cell walls of ground tissue (as shown in the arrow);I:A white substance with low electron density in the cell wall of the cell wall (as indicated by the arrow).C,F,I Abnormal node;A,B,C In the early of primary wall formation;D,E,F In the late stage of primary wall formation;G,H,I The secondary wall formation圖6 基本組織細胞壁的超微結構Fig.6 Ultrastructure of the cell walls of the ground tissue
A:短細胞細胞核邊緣化分布;B:長細胞的內容物多數已降解;C:基本組織長細胞壁次生加厚;D:示維管束橫切面;E:基本組織細胞仍有零星淀粉粒分布;F:示節部異常節淀粉粒的分布.注:F 節部異常節A:Short cell nucleus marginalization distribution;B:Most of the contents of long parenchyma cells has been degraded;C:The long parenchyma cell walls were secondary thickening;D:Showing the transverse section of the vascular bundle;E:Ground tissue cells still have sporadic distribution of starch grains;F:The starch grains distribution of abnormal node.Note:F abnormal node圖7 次生壁形成期基本組織解剖結構Fig.7 Anatomical structure of ground tissue in secondary wall formation
3 討論與結論
3.1 高生長期基本組織細胞壁的發育形成
細胞壁是植物所特有的結構,其結構、成分和狀態與植物細胞的伸長密切相關。
基本組織細胞壁在發育形成的過程中,細胞內含有大量的線粒體、內質網和高爾基體等細胞器。在細胞壁的邊緣分布大量的運輸小泡。內質網是分泌蛋白等物質合成、儲存、加工和運輸的場所,在新壁形成過程中,細胞壁物質經內質網和高爾基體等細胞器的合成加工,分泌運輸小泡,再通過運輸小泡和質膜融合通過胞吐的方式將物質釋放出,參與細胞壁的形成[7]。
植物在生長發育的過程中,當可溶性糖的含量超過自身消耗所需時便會以淀粉的形式儲存。在細胞活躍的時期,便會逐漸的降解為可溶性糖,為植物生長發育提供物質能源。厚竹初生壁發育形成后期,異常節較正常節淀粉粒含量較多,說明該時期異常節細胞代謝緩慢。在次生壁發育形成過程中,部分細胞中還含有極少數的淀粉粒,該時期淀粉粒通過降解為可溶性糖的形式為細胞壁的合成提供物質能量來源。在莖稈生長發育過程中,基本組織細胞始終存在豐富的胞間連絲,胞間隙也逐漸的變大且部分胞間隙能觀察到有降解物的存在,有研究表明,胞間隙可作為質外體運輸的有效途徑[8]。說明基本組織細胞降解的產物通過胞間連絲和胞間隙兩種途徑來實現物質的胞間轉移。賀新強和崔克明[9]通過對植物次生壁形成的研究認為,次生壁是細胞停止生長后在初生壁和質膜之間形成的具有木質素或其他次生代謝產物沉積的具有多層結構的細胞壁。基本組織長、短細胞的細胞壁存在很大的差異,長細胞次生壁逐漸加厚,能觀察到細胞壁不同的壁層,且顏色加深明顯。短細胞僅在與長細胞相鄰的地方次生加厚,這和Liese等[10-13]的研究結果一致。以上結果表明:高生長期,基本組織細胞壁形成過程中,豐富的細胞器參與細胞壁物質的合成,如細胞壁物質經內質網和高爾基體等細胞器的合成,加工分泌運輸小泡,運輸小泡和質膜融合以胞吐的方式將物質釋放出,參與細胞壁的形成,微管系統控制細胞壁物質的沉積排列方向。這個過程中,線粒體為其提供能量來源,淀粉粒的降解為其提供主要物質能源。物質的運輸主要有兩種方式,即共質體運輸和質外體運輸。
本實驗通過對不同發育階段相同部位細胞長度和細胞數量統計數據分析表明,節間伸長是由細胞分裂和細胞伸長共同決定的,細胞分裂是基礎,細胞伸長是導致節間快速伸長的主要原因。本實驗和前人的研究結果相符[14-15]。
3.2 節部在厚竹莖稈快速高生長中的作用
竹稈的迅速高生長主要依賴于節部的居間分生組織。竹稈節間維管束被基本組織細胞完全分開而相互游離,物質的橫向運輸和環流主要在節部進行[16-18]。許婷婷[19]對厚竹異常節和相鄰正常節節部進行解剖結構分析發現,與正常節相比,異常節的節部韌皮結數量較多,而韌皮結是節部物質橫向運輸的重要部位,起到分流和緩沖的作用,故會影響細胞的生長發育。由于節隔的缺失,竹子莖稈節間和節部物質橫向運輸的渠道減少。通過解剖結構分析發現,在初生壁形成的后期,節部異常節較正常節而言,淀粉粒的含量較多,運輸小泡數量較少,細胞壁加厚不顯著,長、短細胞分化不明顯。節間快速伸長主要是由細胞伸長引起的,而細胞的伸長需要物質和能量的供給。綜上,節隔缺失通過引起節間和節部與物質運輸有關結構的變化,使節間細胞伸長所需的物質和能量不能及時的供給,進而影響著節間的伸長,在外部形態上表現為節間縮短。
在次生壁發育形成期,節部異常節基本組織細胞壁與正常節相異之處為:胞間層具有電子密度較低的白色物質;細胞壁內側含有電子密度較低的白色凸起物質。目前,還不能判斷這兩種物質為何物,將在前期研究基礎上,研究兩種物質的成分和功能,進一步揭示莖稈迅速伸長生長的細胞學基礎。
致謝:在整個實驗過程中,許婷婷、胡添翼、苗森浩及竹子資源與利用重點實驗室的老師和同學們對研究給予了幫助,謹致謝意!
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
