長江口水生動物食物網營養結構及其變化
2019-01-07 11:17:46王思凱
水生生物學報 2019年1期
徐 超 王思凱 趙 峰 楊 剛 莊 平
(1. 中國水產科學研究院東海水產研究所, 農業部東海與長江口漁業資源環境科學觀測實驗站, 上海 200090;2. 南京農業大學無錫漁業學院, 無錫 214081)
河口位于淡水與海水交匯處, 環境因子復雜多變, 受到河流與近海的雙重影響。徑流和潮汐在此相互消長, 復雜的水動力條件帶來生物生態習性的多樣性, 使得河口水域幾乎包括了所有生態類型的生物群[1]。長江口是我國第一大河口, 受長江干流徑流和海洋潮汐的交互影響, 形成了獨特的生境條件, 為眾多水生動物的生長和繁殖提供了關鍵棲息生境。此外, 長江徑流輸入的大量營養物質, 使其成為近海生產力較高的水域之一, 孕育了豐富的水生生物資源, 支撐著我國近海漁業可持續發展。然而, 長江口也是受人類干擾最為嚴重的區域, 近年來對長江口資源的大力開發和大型水利工程的建設, 使得長江口生態環境質量和生物多樣性受到前所未有的脅迫和影響。劉瑞玉等[2]研究發現, 三峽大壩建成后長江口及其鄰近水域水文、水化學和沉積環境條件的變化, 引起生物群落組成及其分布格局發生改變, 餌料生物下降, 漁業資源減少; 沈新強等[3]研究顯示長江口水域水環境綜合質量已處于嚴重污染水平, 且呈逐年惡化趨勢, 長江口捕撈產量一直趨于下降, 目前除鳳鱭(Coilia mystus)資源尚屬穩定外, 其余均已形不成魚汛。
食物聯系是河口生態系統結構與功能的基本表達形式, 能量通過食物鏈、食物網轉化為各營養層次生物生產力, 形成生態系統生物資源產量, 并對生態系統的服務和產出及其動態產生影響。因此, 食物網及其營養動力學過程是河口生態系統動力學研究的重要內容, 進而為研究河口生物資源優勢種的交替機制和資源補充機制提供理論依據[4]。對河口食物網營養結構的研究以傳統胃含物分析法為基礎、以簡化食物網為核心, 碳氮穩定同位素技術和生態系統模型的應用發展[5]。羅秉征等[6]運用傳統胃含物分析法研究了長江口20世紀90年代魚類食物網與營養結構, 李忠義等[7]運用穩定同位素技術研究了長江口2005年春季拖網漁獲物的營養級, 林群等[8]基于Ecopath模型研究了長江口及毗鄰水域2000年和2006年生態系統結構和能量流動。近年來, 由于傳統拖網作業和高強度的捕撈壓力及水域環境污染嚴重, 長江口生物資源結構特征突出地表現為經濟種類嚴重衰退, 捕撈對象頻繁更替[9], 這些變化可能導致長江口食物網營養結構的改變。因此, 本文運用傳統胃含物分析法研究了長江口2016—2017年度捕獲的43種水生動物的食性類型、營養級及食物網與能量流動, 并與以往研究結果進行對比, 旨在探討長江口水生動物種間食物關系、營養級及能流途徑的變化情況, 從而為長江口水生生物生態學研究、資源評估、保護及合理利用提供基礎資料。
1 材料與方法
1.1 樣品采集
分別于2016年8月(夏季)、11月(秋季), 2017年2月(冬季)、5月(春季)在長江口水域(31°00′—31°80′N、121°00′—122°40′E)設置22個站點進行4次拖網調查(圖 1)。本次調查所使用的調查船功率為45 kW, 單船桁桿拖網為桁桿長6 m的雙囊網, 網具總長15 m,高2 m, 網囊孔徑2 cm, 每個位點拖網時間長0.5h, 平均航速3 Kn, 捕獲的水生動物用自封袋分裝寫好相應的位點標簽, 放入冰箱, 帶回實驗室進行處理。
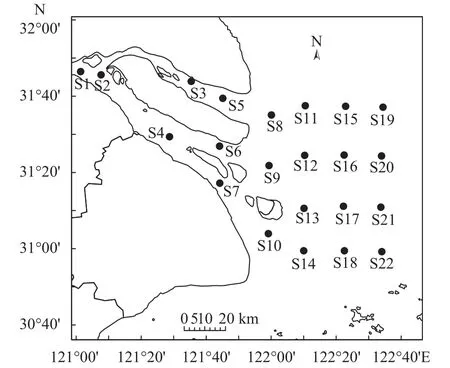
圖 1 長江口采樣地點示意圖Fig. 1 Location of sampling sites in the Yangtze Estuary
1.2 樣品的預處理
野外處理拖網獲得的漁獲物依據不同種類、不同體長、不同采樣位點進行定種分類; 記錄各種漁獲物的名稱并按不同種類進行編號, 測量并記錄各種漁獲物的體長、體重、胃含物重量等, 胃含物用10%的甲醛固定保存; 解剖后的漁獲物分類用自封袋分裝放入-20℃冰箱冷凍保存; 所有樣品帶回實驗室做進一步處理。
實驗室處理胃含物置于OLYMPUS SZX9體式顯微鏡下進行食物種類鑒定, 食料生物的殘體以不易被消化的器官、肢體或外殼作為鑒定的依據, 具體參照表 1[10], 拍照并做好相關記錄。本實驗以胃含物中食物成分的出現頻率、空胃率等為參考依據。
1.3 數據分析
本實驗各種類營養級的相對量度, 參照羅秉征[6]方法以1、2、3、4、5級表示, 1級為自養營養級(浮游植物等); 2—5級為異養營養級, 其中2級為植食性動物; 3級為低級肉食性魚類; 4級為中級肉食性魚類; 5級為高級肉食性魚類。參照羅秉征[6]、張波[11]、竇碩增[12]、張其永等[13]對水生生物食性營養級的研究方法, 用下列公式計算水生生物營養級(TL)、各食物成分的出現頻率(F)及空胃率。

各餌料類群的營養級大小—苔蘚類、介形類、漣蟲類、等足類、糠蝦類、櫻蝦類以及甲殼類幼體均為1.1級, 珊瑚類、腹足類和瓣鰓類為1.2級, 端足類、掘足類和翼足類為1.3級, 海綿類、海葵類、星蟲類、多毛類、海膽類、海百合類、蛇尾類和海參類為1.4級, 水螅類、水母類、歪尾類和橈足類為1.5級, 口足類和短尾類為1.6級, 長尾類為1.8級, 頭足類為2.5級, 魚類為2—3級[各種食料生物類群的營養級大小, 有一部分系根據Odum和Heald數據(轉引自張其永等[13])]

表 1 食物種類的鑒定標準Tab. 1 Identification standards for different types of food
2 結果
本研究共采集到99種水生動物, 由于部分水生動物的捕獲資源量較少且胃含物不易取得與分析,因此, 選取43種水生動物(其中魚類9目17科33種,無脊椎動物10種, 包括蝦類8種, 蟹類2種)835個樣品進行胃含物分析研究。
2.1 食性類型
長江口水生動物消費者的食物組成廣泛且較復雜, 根據其胃含物中不同食物所屬生態類群的出現頻率(表 2), 將長江口水生動物消費者分為4種食性類型。
(1)浮游生物食性: 該類型大多是水體中上層魚類, 以浮游植物、浮游動物(枝角類、橈足類)、有機碎屑、懸浮顆粒物(POM)及較大型的浮游生物如: 中國毛蝦(Acetes chinensis)、甲殼類幼體、魚卵等為食。包括鱸形目的灰鯧(Pampus cinereus)、孔蝦虎魚(Trypauchen vagina)、拉氏狼牙鰕虎魚(Odontamblyopus lacepedii)、中華刺鰍(Sinobdella sinensis), 鯉形目的貝氏?(Hemiculter bleekeri), 鯔形目的鮻(Chelon haematocheilus)、鯔(Mugil cephalus), 鯡形目的日本鳀(Engraulis japonicus)、鳳鱭(Coilia mystus)[14]共9種。此外, 8種無脊椎動物均為浮游生物食性, 包括日本鼓蝦(Alpheus japonicus)、安氏白蝦(Exopalaemon annandalei)、葛氏長臂蝦(Palaemon gravieri)、巨指長臂蝦(Palaemon macrodactylus)、脊尾白蝦(E. carinicauda)等。
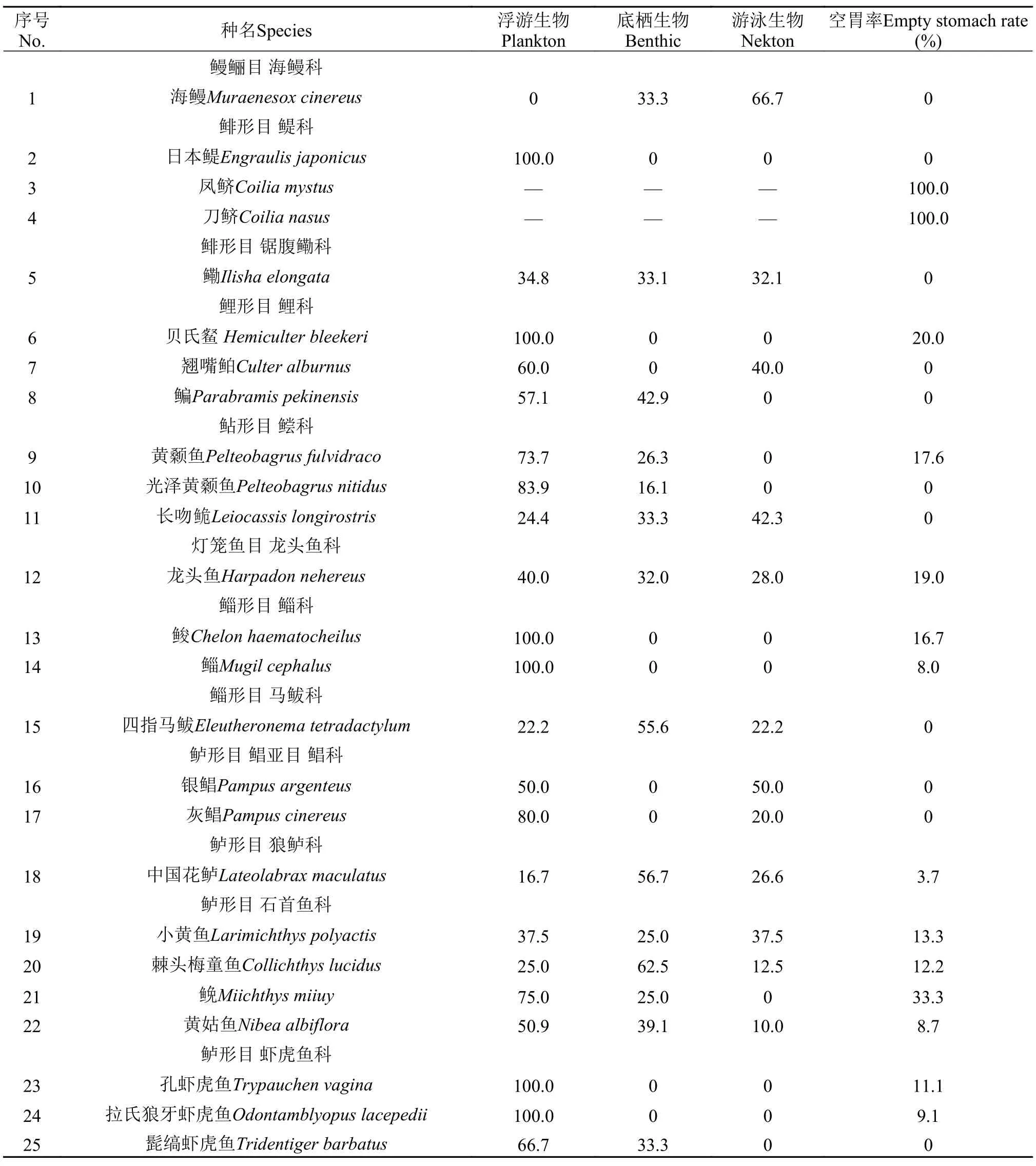
表 2 長江口不同水生動物胃含物中食物所屬生態類群出現頻率百分比Tab. 2 Frequency percentage in the food of different aquatic stomachs in the Yangtze River Estuary

續表 2
(2)底棲生物食性: 該類群魚類性情較溫和, 游泳速度較慢, 多屬于底棲或近底棲魚類, 以底棲藻類、底棲貝類、螺類、底棲端足類、糠蝦類等為食。包括鲇形目的黃顙魚(Pelteobagrus fulvidraco)、光澤黃顙魚(Pelteobagrus nitidus), 鱸形目的香斜棘?(Repomucenus olidus)、斑尾刺蝦虎魚(Acanthogobius ommaturus)、矛尾蝦虎魚(Chaeturichthys stigmatias)、髭縞蝦虎魚(Tridentiger barbatus)、鮸(Miichthys miiuy)、金錢魚(Scatophagus argus), 鯉形目的鳊(Parabramis pekinensis), 鰈形目的短吻紅舌鰨(Cynoglossus joyneri), 鯡形目的刀鱭(Coilia nasus)[15]共11種。此外, 2種無脊椎動物三疣梭子蟹(Portunus trituberculatus)、日本鱘(Charybdis japonica)為底棲生物食性。
(3)游泳生物食性: 該類型既有底層魚類, 又有中上層魚類, 性兇猛貪食, 游泳活動能力強, 以游泳動物(魚類、頭足類)鯔、鮻、龍頭魚、矛尾鰕虎魚、水母等為食。包括鱸形目的中國花鱸(Lateolabrax maculatus)、銀鯧(Pampus argenteus), 鲇形目的長吻 (Leiocassis longirostris), 燈籠魚目的龍頭魚(Harpadon nehereus)以及鰻鱺目的海鰻(Muraenesox cinereus)共5種。
(4)混合(底棲生物和游泳生物)食性: 該類型魚類食譜很廣, 在長江口魚類中種類較多, 其食物類群既有底埋生物、底面層生物又有底層生物和游泳生物。包括鯡形目的鰳(Ilisha elongata), 鱸形目的黃姑魚(Nibea albiflora)、小黃魚(Larimichthys polyactis)、棘頭梅童魚(Collichthys lucidus)、睛尾蝌蚪蝦虎魚(Lophiogobius ocellicauda), 鲀形目的黃鰭東方鲀(Takifugu xanthopterus), 鯔形目的四指馬鲅(Eleutheronema tetradactylum), 鯉形目的翹嘴鲌(Culter alburnus)共8種。
2.2 營養級
本研究分析了43種水生動物的營養級(表 3),結果表明, 這43種水生動物營養級主要集中在2.0—4.0級, 其中植食性魚類和無脊椎動物類位于最低的2.0—3.0級, 低級肉食性魚類在3.0—3.5級,中級肉食性魚類位于最高的3.5—4.0級。在所有消費者中, 植食性消費者所占比例最高為76.75%, 低級肉食性魚類為18.60%, 中級肉食性魚類最低為4.65%。在所有魚類樣品中, 海鰻營養級最高為3.68; 而在無脊椎動物樣品中, 三疣梭子蟹營養級最高為2.53。
羅秉征等[6]曾經分析了長江口20世紀90年代60種常見魚類的營養級大小。本研究選取了所采集物種中與之相同的12種魚類進行比較分析, 結果表明這12種魚類在2016—2017年的營養級比20世紀90年代普遍偏低(圖 2)。
2.3 食物網及能流途徑
根據各消費者的胃含物種類, 本研究確定了長江口水生食物網3種主要的生產者類型, 即底棲藻類、浮游植物和有機碎屑及POM。結合各消費者的取食關系和營養級, 繪制了長江口水生生物食物網3種能流途徑(圖 3)。
(1)底棲藻類→中下層植食性魚類(鮻等)→低級肉食性魚類(龍頭魚等)→中級肉食性魚類(海鰻)。
(2)浮游植物→植食性無脊椎動物(葛氏長臂蝦等)→低級肉食性魚類(小黃魚等)→中級肉食性魚類(海鰻)。
(3)有機碎屑、POM→中上層植食性魚類(鳀等)→低級肉食性魚類(四指馬鲅等)→中級肉食性魚類(海鰻)。
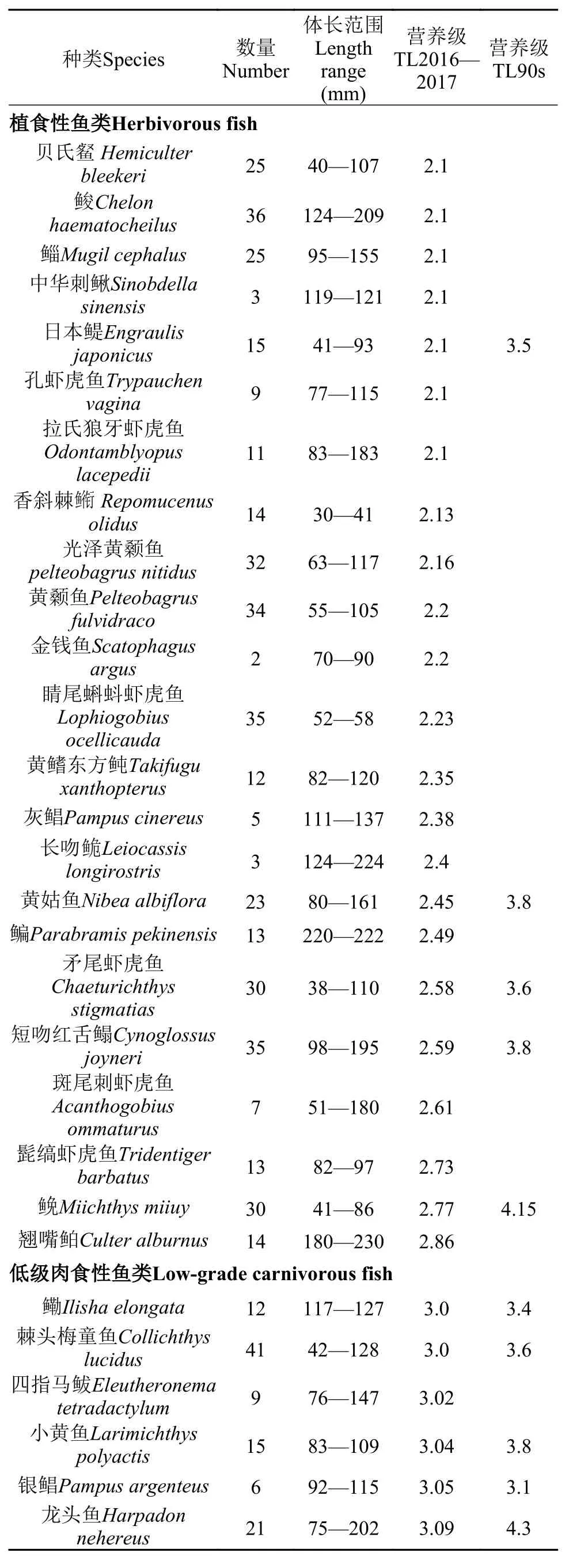
表 3 長江口水生動物2016—2017年及20世紀90年代營養級Tab. 3 Trophic levels of aquatic animals in the Yangtze Estuary during 2016 to 2017 and in the 1990s
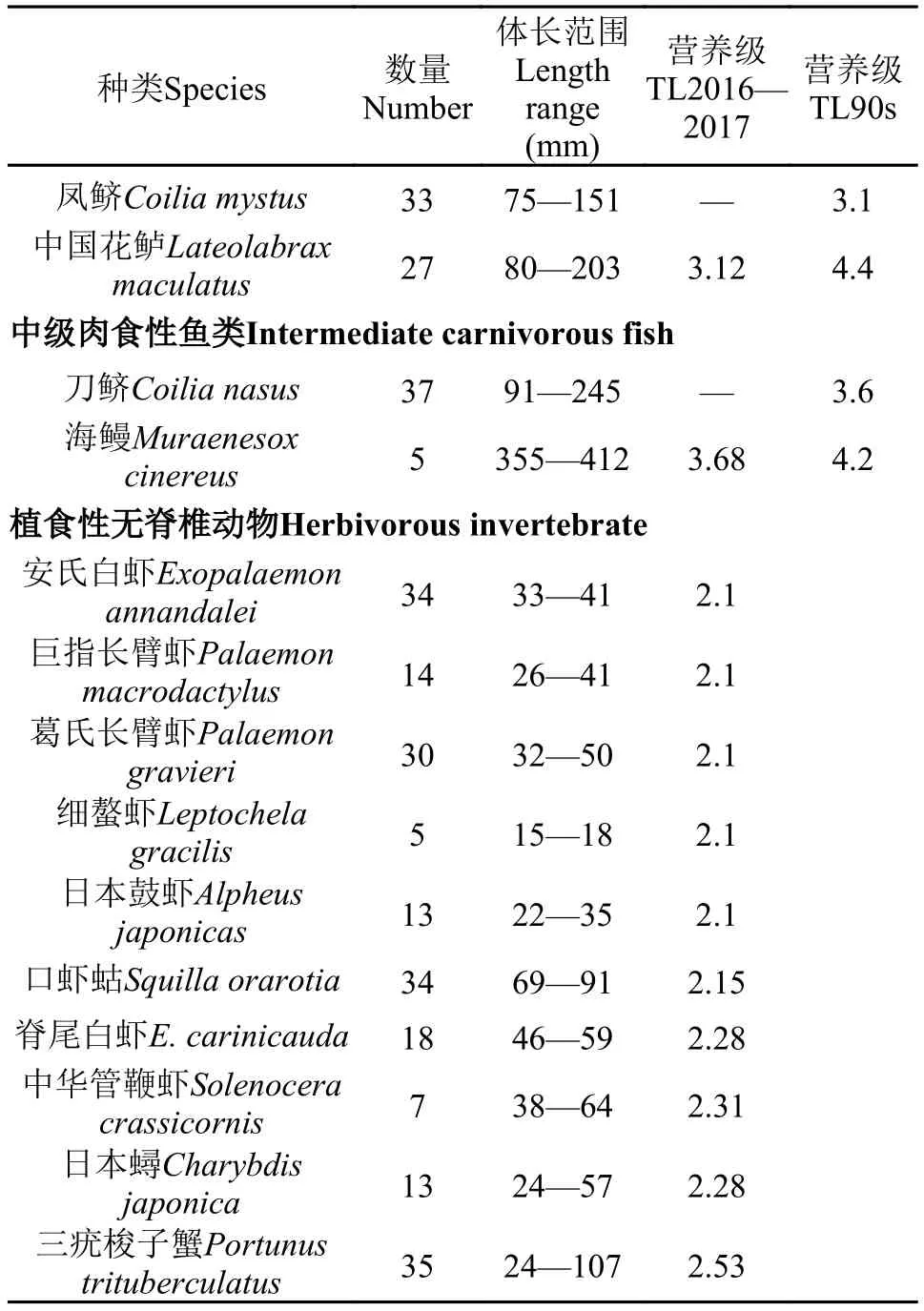
續表 3
一些植食性魚類如鮻、鯔、貝氏?以浮游植物(綠藻、硅藻等)、浮游動物(橈足類、枝角類、糠蝦類、磷蝦類等)為主要食物, 同時又被大型兇猛魚類如中國花鱸、刀鱭、龍頭魚、海鰻等捕食。底棲蝦、蟹類如脊尾白蝦、安氏白蝦、葛氏長臂蝦、三疣梭子蟹既捕食小型底棲餌料生物, 同時又被其他魚類如四指馬鲅、棘頭梅童魚等捕食。各種河口食物鏈緊密連接在一起, 構成了長江口錯綜復雜的食物網(圖 4)。
3 討論
研究發現長江口水生動物的食性類型分為4種,營養級介于2.0—4.0級。長江口水生生物的食物網結構較為復雜, 主要由牧食食物鏈和碎屑食物鏈構成的復雜食物網。此外, 長江口水生生物資源棲息環境日益遭到破壞, 生物資源量處于退化狀態, 平均營養級與歷史相比有所降低[16,17]。
3.1 資源種類

圖 2 長江口12種魚類20世紀90年代與2016—2017年營養級大小比較Fig. 2 Comparing the trophic level of 12 fish species in the Yangtze Estuary in the 1990s and during 2016 to 2017
長江口近海漁業資源在我國海洋漁業上占有舉足輕重的地位[18]。長江口漁業生物主要包括魚類、甲殼類、頭足類共3個生態類群[19]。從各生態類群的漁獲重量占總漁獲量不同季節平均百分比可見: 每個季節都是以魚類為主要優勢群, 所占比例在70.65%—88.10%; 頭足類和甲殼類則為次要組成部分, 在各個季節所占的重量百分比都比較低。其中, 甲殼類所占總漁獲量百分比平均值為15.78%;頭足類所占總漁獲量百分比最高為4.80%, 最低僅為0.10%。從各生態類群種類數的季節及年間變化可知, 群落生物量、多樣性以夏秋季節最大, 春冬季節最小。其中, 魚類種類秋季最多, 春季最少; 甲殼類種類秋季最少, 其余三季種類差異不大; 頭足類種類四季變化差異均不明顯。這與李建生等[20]研究的長江口2000—2002年漁場漁業生物群落結構的季節變化結果相吻合。此外, 本次研究中長江口2016—2017年水生生物季節間總種類數的變化為從春季經夏季到秋季呈現上升的趨勢, 從冬季到來年春季呈下降的趨勢。種類數和資源密度的最低值并不出現在同一季節, 造成這種差異的原因可能與漁業生物的群體構成有關, 因為夏季主要是產卵群體, 個體相對較大, 所以雖然種類數較少但是資源密度值卻不是最低的[20]。

圖 3 長江口水生食物網能流途徑簡化模式Fig. 3 Simplified model of energy flow path for Yangtze Estuary Aquatic animals' food web
2017年度捕獲的水生生物種類共65種, 相同取樣位點2016與2015年度捕獲的水生生物種類則分別為73種和84種。這表明長江口水生生物的種類逐步減少, 資源量逐漸降低。長江口經濟魚類包括: 銀鯧、鰳、黃姑魚、鳳鱭、刀鱭、棘頭梅童魚、龍頭魚等[21]。目前, 主要經濟種群都已處于過渡利用的狀態。由于不遵循自然規律, 捕撈壓力過大, 結果反而使單位產量下降, 水生生物中主要傳統經濟種類日益減少, 而一些非經濟種類和不可食用幼體的比重增加, 漁獲質量下降。生物資源作為一種可更新的資源, 必須進行繁殖保護, 加強科學管理與合理開發利用, 才能達到可持續利用長江口水生生物資源[21]。
3.2 食性
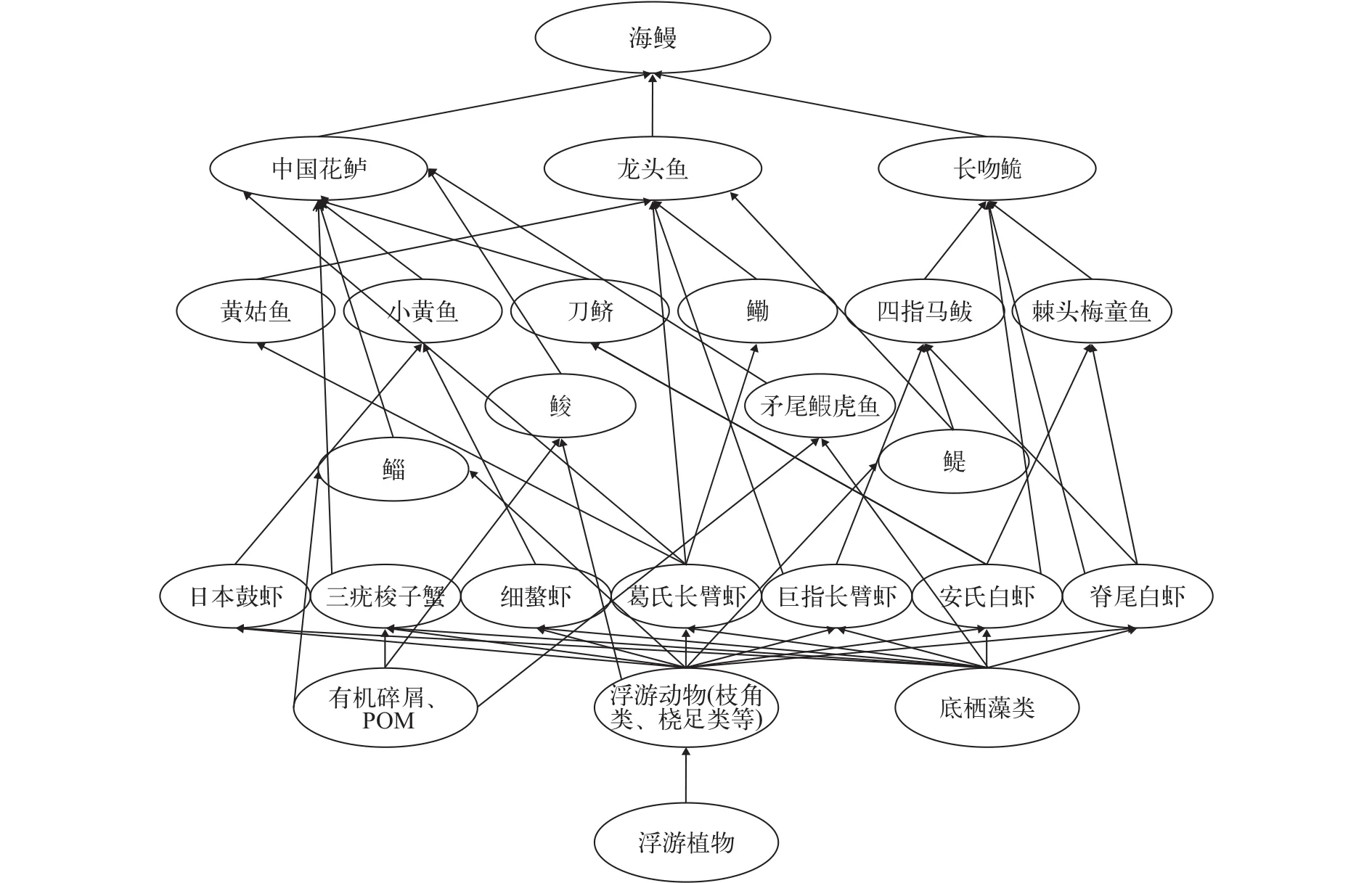
圖 4 長江口主要水生生物食物網Fig. 4 Food web of main aquatic in the Yangtze Estuary
本研究胃含物分析結果表明長江口水生動物的食性分為浮游生物食性、底棲生物食性、游泳生物食性、混合食性4種類型。羅秉征等[6]研究了長江口20世紀90年代魚類食性類型, 同樣分為4種食性類型, 其中浮游生物食性魚類以橈足類, 如哲水蚤、劍水蚤及猛水蚤等浮游動物為主要食物; 本實驗中浮游生物食性魚類則主要以浮游植物、有機碎屑、POM等為主要食物, 且浮游植物及有機碎屑在食物中所占比例較大。造成這一差異的原因主要與采樣位點有關, 本實驗采樣點集中在長江口門及口內水域, 而羅秉征等[6]的研究則集中在長江口外及外海水域; 此外, 隨著長江徑流源源不斷的入海和營養物質的大量輸入, 使得長江口有來自陸地淡水或由海水帶來的大量碎屑、細菌和其他異養性微生物, 河口可以將它們分解成溶解的顆粒有機物[22], 從而為長江口浮游生物食性魚類提供了豐富的食源。
此外, 在本實驗中浮游生物食性種類最多, 所占比例最大, 為39.53%; 游泳生物食性種類最少, 所占比例最小, 僅為11.63%。由于目前長江口捕撈壓力的增大, 長江口水生生物的資源量銳減, 經濟價值低、個體小、年齡結構簡單和營養層次低的類群在長江口及其鄰近海域占據優勢地位[23]。因而長江口浮游生物食性水生生物所占比例越來越高,游泳生物食性水生生物所占比例逐漸降低。
3.3 營養級
本研究結果表明長江口水生動物營養級集中在2.0—4.0級, 普遍小于羅秉征等[6]、張波等[11]在長江口的研究結果。一方面同一物種的營養級大小降低, 例如12種優勢魚類營養級大小均小于20世紀90年代的研究值, 平均營養級從3.80下降到2.87, 中國花鱸與前期研究值相差甚至達到1.28級; 另一方面高營養級物種種類和數量降低, 如低級肉食性魚類所占比例由20.00%[6]下降到18.60%。
隨著捕撈強度的增大, 長江口水生生物資源量處于退化狀態。李建生等[24]研究發現長江口漁場漁業生物資源2003年各調查航次代表季節的漁業生物種類數與2000—2002年各年份同期相比, 種類數有不同程度的降低。以底拖網為主的捕撈漁業使底層的大型經濟魚類迅速減少, 漁業生物捕食關系發生改變, 水生生物逐漸由肉食性、長壽的底層魚類向浮游生物食性、短壽的上層魚類轉變[23]; 此外, 長江口及其鄰近海域由于長期受到環境污染和漁場海洋環境條件的變化, 如三峽大壩的成功截流、洋山港深水碼頭以及長江口深水航道治理工程的建設等因素, 從而導致水生生物群落結構發生顯著變化[25]。Ryder等[26]指出, 過度捕撈和環境退化使生物群落的生態系統失去恢復力和完整性, 生態系統的穩定性變差, 使依賴生態系統產出的漁業產量在質和量2個方面具有不可預見的變化。因而長江口水生生物的營養級降低很大程度上應該是環境因素和捕撈因素的共同作用結果[27]。此外, 本實驗調查采樣主要以底拖網為主, 拖網生產的漁獲物組成主要以底層及中下層魚類為主, 中上層魚類如鰳、刀鱭等也占有一定比例, 因而計算得出的營養級整體較低。
部分魚類如鮸的營養級2.77級與張波等[11]的4.15級相差較大, 這與捕獲鮸個體的大小有關, 鮸平均體長421.6 mm, 為肉食性魚類, 食量大, 以魚蝦為主要餌料[28], 而捕獲到的鮸平均體長僅63.5 mm,因此營養級相差較大; 又如長吻 為肉食性魚類,因為主要是冬、春季捕獲, 攝食量少且捕獲的體長較短, 平均體長僅為182.3 mm, 因此營養級較低, 僅為2.4級。此外這兩種魚類營養級的降低還與餌料種類的變化及捕撈壓力等有關。
3.4 食物網
從構建的簡化食物網分析, 長江口漁業資源群落食物網的頂級捕食者從高營養級階層生物逐漸轉變為中低營養級階層生物, 食物鏈越來越短。通過食物網和能流途徑可以發現: 浮游植物、底棲藻類、有機碎屑等在食物網中作為初級生產者為初級消費者浮游動物提供食物; 而初級消費者浮游動物則被次級消費者底棲無脊椎動物及植食性魚類所攝食; 次級消費者又被低、中級肉食性魚類所捕, 能量逐級傳遞, 從而構成長江口復雜的食物網。其中葛氏長臂蝦、安氏白蝦、脊尾白蝦、巨指長臂蝦、三疣梭子蟹等在食物網中作為重要的食物餌料起著關鍵作用, 由于它們大多數是長江口及鄰近水域的優勢生物種, 在該地區分布廣、數量較多、棲息時間較長、所以為該地區經濟魚類的繁衍和持續提供了可靠的食物保證[6]。同時, 由于它們本身也是重要的可開發經濟漁獲物, 所以合理利用及保護這些漁業資源對保護該水域漁業生產力及其生物多樣性具有重要意義。
總之, 長江口水生生物一方面通過攝食空間及攝食方式的不同產生隔離, 在一定程度上避免了時空上的攝食競爭; 另一方面通過營養生境的重疊及捕食者與被捕食者的相互捕食關系把長江口水生生物通過復雜的食物鏈、食物網連接起來, 從而保證了該水域水生生物持續、穩定的發展[6]。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
今日農業(2021年11期)2021-08-13 08:53:34
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
