脊尾白蝦2種血藍蛋白大亞基變體的克隆及功能分析
2019-01-07 11:17:36竇全偉李吉濤劉九美孫東方環朋朋
水生生物學報 2019年1期
竇全偉 李吉濤, 劉 萍, 李 健, 劉九美 孫東方 蔡 影 環朋朋
(1. 中國水產科學研究院黃海水產研究所, 農業部海洋漁業可持續發展重點實驗室, 青島 266071;2. 青島海洋科學與技術國家實驗室, 海洋漁業科學與食物產出過程功能實驗室, 青島 266235)
脊尾白蝦(Exopalaemon carinicauda)是分布于中國大陸沿岸、朝鮮半島西岸的淺海低鹽水域的一種熱溫帶海區中小型底棲蝦類, 以黃、渤海產量最高, 是我國重要的經濟蝦類之一[1]。脊尾白蝦具有廣溫性[2]、廣鹽性[3]、繁殖率高[4,5]等特點, 具有較高的經濟價值, 是新興的養殖蝦類, 已成為沿海灘涂地區的主要特色水產養殖品種。
由細菌及病毒等微生物引起的疾病對蝦類養殖造成嚴重的經濟損失[6], 因此對蝦類非特異性免疫防御機制的研究正在廣泛開展。研究表明, 血藍蛋白不但具有載氧功能, 還有儲存能量、維持滲透壓、調節蛻皮及固化表皮的功能[7]。此外, 大量研究證明甲殼動物血藍蛋白也是免疫防御體系中的重要免疫因子, 血藍蛋白及其降解片段還具有凝集活性、抗菌抗病毒活性、酚氧化物酶活性、抗腫瘤活性等多種免疫學功能, 被認為是一種具有重要免疫活性的多功能蛋白[8—11]。目前, 隨著對甲殼動物血藍蛋白功能的深入研究, 其基因cDNA序列已相繼被克隆[12—14]。研究表明, 凡納濱對蝦(Litopenaeus vannamei)有2個血藍蛋白亞基, 且哈維氏弧菌(Vibrio harveyi)感染后其mRNA表達量顯著上調[15]。Tassanakajon等[16]研究發現斑節對蝦(Penaeus monodon)血藍蛋白五種不同亞型在序列及長度上存在差別, 且具有抗菌活性。郭玲玲等[17]以凡納濱對蝦為研究對象, 通過生物信息學方法分析發現凡納濱對蝦血藍蛋白大亞基中存在HcLV1、HcLV2、HcLV3等多種變體, 并采用RACE (Rapid-amplification of cDNA ends)技術獲得HcLV1、HcLV3的cDNA全長, 通過副溶血弧菌(Vibrio Parahemolyticus)、乙型鏈球菌(Streptococcus)及溶藻弧菌(Vibrio alginolyticus)等不同病原菌刺激對蝦后, 血藍蛋白大亞基變體對不同病原刺激發生明顯的響應,且不同病原刺激后可致使不同變體協同作用。Xu等[18]研究發現凡納濱對蝦血藍蛋白大亞基至少存在四種變體且均具有抗病毒活性, 提示血藍蛋白具有分子多樣性, 且在對蝦抗病免疫防御中有重要作用。我們已經獲得了脊尾白蝦血藍蛋白大亞基基因[19],并對其免疫功能進行了初步研究, 但其變體及其功能的研究尚未見報道。
本研究根據本實驗室構建的脊尾白蝦轉錄組文庫, 篩選獲得了2個血藍蛋白變體序列, 采用RACE技術克隆了脊尾白蝦血藍蛋白大亞基變體基因的cDNA全長, 并分析了變體基因在金黃色葡萄球菌、副溶血弧菌及白斑綜合征病毒(White spot syndrome virus, WSSV)感染后基因的相對表達變化, 對豐富脊尾白蝦血藍蛋白的分子多態性與功能多樣性具有重要的意義。
1 材料與方法
1.1 材料
取體長(4.5±0.25) cm、體重(2.19±0.42) g的健康脊尾白蝦暫養3d, 設置金黃色葡萄球菌感染組、副溶血弧菌感染組、WSSV感染組和對照組, 每組100尾, 設置3個平行, 分別進行感染實驗。
1.2 菌懸液及WSSV粗提液的制備
配制LB肉湯和2216E(液體成分: 蛋白胨母膏,磷酸高鐵; 固體成分加瓊脂粉)液體及固體培養基,滅菌備用。將金黃色葡萄球菌(Stphylococcus aureus)和副溶血弧菌(Vibrio parahaemolyticus)菌種分別接種于LB肉湯和2216E固體培養基中, 分別在37℃和28℃培養過夜。選取單菌落接種于液體培養基中, 200 r/min搖床上培養8—10h, 最后將菌懸液離心, 棄上清液, 用生理鹽水對菌體進行反復洗滌, 重懸后, 使菌體的濃度達108CFU/mL。
取感染WSSV的脊尾白蝦病蝦(本實驗室保存)頭尖組織5 g, 加入800 μL 4℃預冷的磷酸鹽緩沖液(1×PBS), 4℃ 20000 r/min離心15min, 將獲得的勻漿液于4℃ 3000 r/min離心15min, 取上清液反復離心3次(4000 r/min 15min、6000 r/min 15min、8000 r/min 15min), 將所得的上清液用0.45 μm濾膜過濾除菌,將除菌的粗提液稀釋100倍(3.7×107copy/mL)進行分裝并保存于-80℃冰箱中備用。
實驗時將菌懸液、WSSV粗提液用微量注射器從脊尾白蝦的第2腹節處注入(20 μL/尾)。對照組注射等量1×PBS緩沖液。在注射后的6h、12h、24h、48h和72h取樣, 每個時間點各取5尾蝦, 取血液、肝胰腺、鰓、肌肉等組織, 用于總RNA提取。
1.3 脊尾白蝦總RNA提取及cDNA合成
使用TransZol Up Plus RNA Kit (北京全式金生物技術有限公司)提取脊尾白蝦組織總RNA, 用核酸定量儀(Thermo, NanoDrop 2000)和1.0%瓊脂糖凝膠電泳檢測總RNA的質量及完整性。
cDNA合成體系(20 μL): 10 μL總RNA, 2 μL Oligo dT (50 μmol/L), 72℃水浴5min, 冰浴2min; 之后再向管中加入1.0 μL dNTP Mixture (each 10 mmol/L)、5×M-MLV Buffer 4.0 μL、0.5 μL RNase Inhibitor(40 μL/mL, TaKaRa)和1.0 μL M-MLV反轉錄酶(TaKaRa), 用DEPC水補足體積。42℃孵育lh; 72℃孵育15min; 4℃孵育20min。合成的cDNA用于后期脊尾白蝦血藍蛋白基因的克隆及Real-time PCR檢測。脊尾白蝦3′和5′RACE模板的合成根據SMARTTMRACE Amplification Kit9 (TaKaRa)說明書進行。
1.4 脊尾白蝦血藍蛋白大亞基變體基因cDNA中間片段克隆
根據本實驗室脊尾白蝦轉錄組庫中獲得的脊尾白蝦血藍蛋白大亞基變體基因EcHcL1和2序列設計引物HcL1-F/HcL1-R、HcL2-F/HcL2-R (上海生工生物工程有限公司合成)(表 1)。PCR反應體系(10 μL): 正反引物各0.4 μL, LA Mix 5 μL, cDNA模板0.2 μL, ddH2O 4 μL。根據引物退火溫度設定反應程序。

表 1 本研究所用引物序列Tab. 1 Sequences of the primers used in this study
1.5 脊尾白蝦血藍蛋白大亞基變體基因cDNA 5′和3′末端的擴增
根據測序得到的中間片段序列設計5′ RACE引物(5HcL1和5HcL2)以及3′ RACE引物(3HcL1和3HcL2)(表 1)擴增脊尾白蝦大亞基血藍蛋白大亞基變體基因cDNA全長序列。按照SMARTTMcDNA Amplification Kit (Clontech)說明書推薦的反應體系及反應條件進行5′ RACE和3′ RACE擴增。
1.6 序列分析
將測序結果去載體后, 采用ContigExpress Application進行5′和3′端序列拼接, 用NCBI BLAST(http://blast.ncbi.nlm.gov/Blast.cgi)和SMART (http://smart.embl-heidelberg.de/)進行序列比對及結構域預測, 應用SignalP 4.0軟件分析信號肽, 通過DNAMAN軟件進行血藍蛋白的氨基酸序列多序列比對,用MEGA 6.0軟件[20]中的Neighbor-joining法[21]構建系統樹。
1.7 脊尾白蝦血藍蛋白大亞基變體基因組織分布特征分析
根據脊尾白蝦血藍蛋白大亞基變體基因EcH-cL1和2基因設計正反向引物(QHcL1-F/QHcL1-R、QHcL2-F/QHcL2-R)(表 1), 用于Real-time PCR檢測EcHcL1、2基因的組織分布情況。使用SYBR Premix ExTaqⅡ試劑(TaKaRa), Applied Biosystems 7500 Real Time PCR儀上進行, 反應體系為10 μL:5 μL SYBR Premix ExTaqⅡ、1 μL cDNA、0.4 μL正反向引物、0.2 μL ROX Reference dye Ⅱ及3 μL ddH2O。反應程序為95℃ 10min; 95℃ 30s, 95℃ 5s,60℃ 34s, 40個循環; 95℃ 15s; 60℃ 1min; 95℃15s。以β-actin基因作為內參基因, 樣本和內參均設置3個平行, 采用2-ΔΔCt方法[22]計算EcHcL1和2基因的相對表達量, 利用SPSS17.0軟件對結果進行單因素方差分析(One-way, ANOVA), 并對其顯著性進行檢驗。
1.8 病原感染脊尾白蝦后血藍蛋白大亞基變體基因的表達分析
分別提取副溶血弧菌、金黃色葡萄球菌及WSSV感染后不同時間脊尾白蝦的肝胰腺和血細胞組織的RNA, 按照前述方法逆轉錄合成cDNA進行Real-time PCR, 以β-actin作為內參, 樣本和內參均設置3個平行, 檢測細菌感染下的脊尾白蝦血細胞和肝胰腺中EcHcL1和2基因在不同時間點的表達情況。反應體系、反應程序及數據處理同如1.6所述。
2 結果
2.1 脊尾白蝦血藍蛋白大亞基變體基因EcHcL1和2全長cDNA序列的克隆與分析
采用RACE方法和RT-PCR擴增獲得脊尾白蝦血藍蛋白大亞基變體基因EcHcL1和2 cDNA全長(GenBank登錄號: MH069500和MH 069501)。EcH-cL1序列全長2248 bp, 包括2058 bp的開放閱讀框(ORF)、66 bp的5′ UTP和137 bp的3′ UTR; 編碼685個氨基酸, 前21個氨基酸組成信號肽; 預測分子量為79.8 kD 。EcHcL2序列全長2121 bp (ORF:2031 bp), 10 bp的5′ UTP和80 bp的3′ UTR; 編碼676個氨基酸, 前27個氨基酸組成信號肽; 預測分子量為78.3 kD, 理論等電點PI為5.56 。SMART軟件分析表明, EcHcL1-2-3均含有3個結構域: Hemocyanin_N、Hemocyanin_M、Hemocyanin_C, 只是EcHcL1多了一個酪氨酸酶結構域(圖 1)。Hemoeyanin-M結構域中的2個銅離子結合位點含6個保守的組氨酸殘基。三維結構預測軟件(https://swiss-model.expasy.org/)顯示EcHcL1-2三維結構出現了差異(圖 2): 在529—650 aa,EcHcL1以β-折疊為主,EcHcL2以無規則卷曲為主。

圖 1 脊尾白蝦EcHcL1和2血藍蛋白結構域位置Fig. 1 The locations of three domains in EcHcL1 and 2 hemocyanin A. EcHcL1; B. EcHcL2

圖 2 脊尾白蝦EcHcL1和2血藍蛋白三維結構預測Fig. 2 Three-dimensional structure of EcHcL1 and 2 A. EcHcL1; B. EcHcL2
2.2 脊尾白蝦血藍蛋白EcHcL1和2基因同源性分析
使用NCBI BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi)對脊尾白蝦EcHcL1和2基因編碼的氨基酸序列與其他物種的血藍蛋白氨基酸序列進行同源性比較, EcHcL1和2在氨基酸序列上與日本沼蝦同源性最高達到84.08%, 與凡納濱對蝦、日本對蝦(Marsupenaeus japonicus)以及斑節對蝦(Penaeusmonodon)血藍蛋白序列同源性分別為77.54%、76.66%和76.39%。
利用MEGA 6.0軟件對脊尾白蝦EcHcL1和2基因氨基酸序列進行系統進化分析, 并構建系統進化樹(圖 3)。結果顯示在15個物種中, 脊尾白蝦EcH-cL1和2與脊尾白蝦EcHcL以及日本沼蝦(Macrobrachium nipponense)為一支并與中國對蝦(Fenneropenaeus chinensis)、日本對蝦、凡納濱對蝦、斑節對蝦等動物的血藍蛋白為一個亞群; 與紅鰲鰲蝦(Cherax quadricarinatus)、珍寶蟹(Metacarcinus magister)、中華絨螯蟹(Eriocheir sinensis)、藍蟹(Callinectes sapidus)、擬穴青蟹(Scylla paramamosain)為另一個亞群。EcHcL1和2在進化上與日本沼蝦的親緣關系最近, 與克氏原鰲蝦(Procambarus clarkii)、鯨虱(Cyamus scammoni)、斑點海虱(Eurydice pulchra)、蛀木水虱(Limnoria quadripunctata)等動物的血藍蛋白親緣關系較遠。
2.3 脊尾白蝦血藍蛋白EcHcL1和2基因組織表達分析

圖 3 利用MEGA 4.0構建的基于EcHcL1和2基因氨基酸序列的NJ系統樹Fig. 3 NJ tree about EcHcL1-2 amino acid sequences using MEGA 4.0
為了分析EcHcL1和2的組織表達特異性, 分別設計2種變體的特異性引物。以β-actin作為內參對照, 利用Real-time PCR檢測脊尾白蝦EcHcL1和2在血細胞、肝胰腺、腸、腹神經節、心臟、卵巢、眼柄、胃、鰓和肌肉組織中的表達量。結果如圖 4顯示,EcHcL1和2在脊尾白蝦的10種組織中均有表達。EcHcL1在血細胞中表達最高, 肝胰腺次之, 而在肌肉中的表達量最低;EcHcL2在肝胰腺中表達最高, 而在肌肉和胃中的表達量最低。

圖 4 EcHcL1和2在脊尾白蝦不同組織里的相對表達量Fig. 4 The relative expression of EcHcL1 and 2 in different tissues in E. carinicauda
2.4 脊尾白蝦血藍蛋白EcHcL1和2基因在肝胰腺中的表達
如圖 5、圖 6所示, 脊尾白蝦感染金黃色葡萄球菌、副溶血弧菌、WSSV后, 脊尾白蝦肝胰腺中EcHcL1和2基因表現出一定的時間依賴性, 總體趨勢表現為先升高后降低。EcHcL1表達量在金黃色葡萄球菌和WSSV感染后均在12h達到峰值, 與對照組有極顯著差異(P<0.01), 到72h下降到與對照組無顯著性差異(P>0.05);EcHcL1表達量在副溶血弧菌感染后在24h達到峰值, 與對照組差異極顯著(P<0.01),24h后表達量持續下降,到72h下降到與對照組無顯著性差異(P>0.05)。EcHcL2表達量在金黃色葡萄球菌感染后趨勢不明顯, 在48h達到峰值,與對照組有顯著性差異(P<0.05), 到72h下降到與對照組無顯著性差異(P>0.05); 在副溶血弧菌和WSSV感染后均在24h達到峰值, 與對照組差異極顯著(P<0.01), 24h后表達量持續下降, 到72h下降到與對照組無顯著性差異(P>0.05)。
2.5 脊尾白蝦血藍蛋白EcHcL1和2基因在血細胞中的表達分析

圖 5 EcHcL1基因在脊尾白蝦肝胰腺組織中的表達變化Fig. 5 Expression level of EcHcL1 in hepatopancreas of E. carinicauda
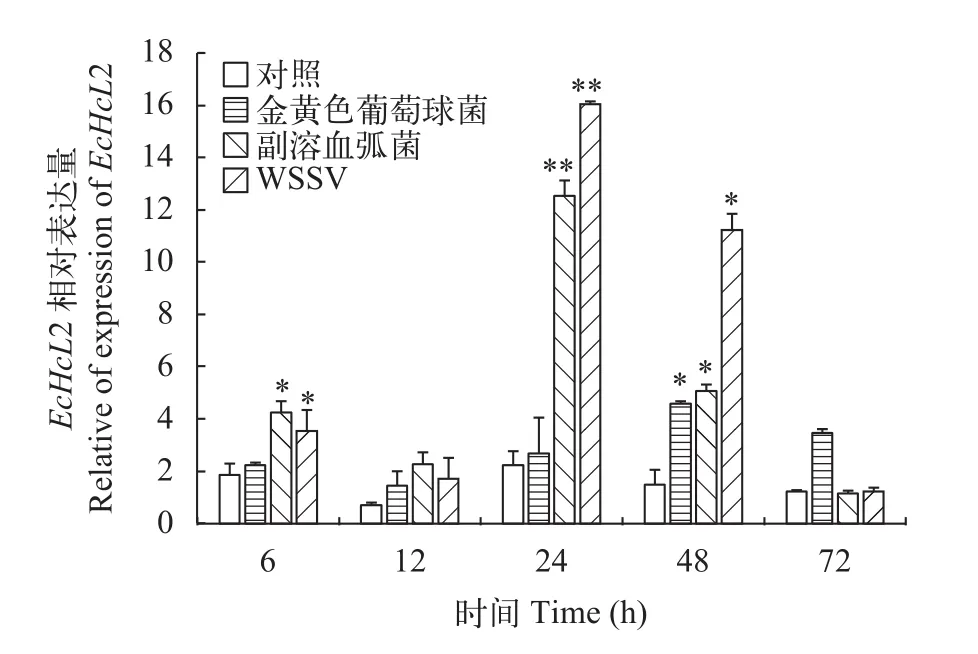
圖 6 EcHcL2基因在脊尾白蝦肝胰腺組織中的表達變化Fig. 6 Expression level of EcHcL2 in hepatopancreas of E. carinicauda

圖 7 EcHcL1基因在脊尾白蝦血細胞中的表達變化Fig. 7 Expression level of EcHcL1 in haemocytes of E. carinicauda

圖 8 EcHcL2基因在脊尾白蝦血細胞中的表達變化Fig. 8 Expression level of EcHcL2 in haemocytes of E. carinicauda
如圖 7、圖 8所示, 脊尾白蝦感染金黃色葡萄球菌、副溶血弧菌、WSSV后, 脊尾白蝦血細胞中EcHcL1和2基因表現出一定的時間依賴性, 總體趨勢表現為先升高后降低。EcHcL1表達量在金黃色葡萄球菌感染后均在12h達到峰值且與對照組差異顯著(P<0.05), 到72h下降到與對照組無顯著性差異(P>0.05); 在副溶血弧菌和WSSV感染后均在48h達到峰值, 與對照組差異極顯著(P<0.01), 到72h下降到與對照組無顯著性差異(P>0.05)。EcHcL2表達量在金黃色葡萄球菌感染后趨勢不明顯; 在副溶血弧菌和WSSV感染后均在24h達到峰值, 且WSSV感染后表達量上升趨勢明顯大于副溶血弧菌感染后的表達量。
3 討論
血藍蛋白是甲殼動物血淋巴中的含銅呼吸蛋白, 它的基本生物學功能是載氧, 另外它還具有多種免疫活性, 例如酚氧化酶活性、抗病毒活性、凝集活性和抗菌活性等多種免疫功能[23—25]。為了深入研究脊尾白蝦大亞基的分子特征, 我們采用RACE和生物信息學技術, 對脊尾白蝦血藍蛋白大亞基2種變體(EcHcL1和2)進行全長cDNA擴增和序列分析。我們發現EcHcL1和2與脊尾白蝦血藍蛋白大亞基EcHcL相似, 其分別編碼680個左右的氨基酸。兩者均具有血藍蛋白的典型結構域, 包括銅離子結合區, 6個組氨酸位點和Ig-like區等, 其中EcHcL1多了一個酪氨酸酶結構域, 推測可能與色素代謝及酚氧化酶活性有關[23]。系統進化分析結果顯示EcHcL與日本沼蝦血藍蛋白親緣關系最近,疣酋婦蟹、中華絨螯蟹、斑點海虱等甲殼動物的血藍蛋白親緣關系次之。這說明脊尾白蝦血藍蛋白大亞基變體與大亞基具有相似的分子特征, 提示其變體可能與血藍蛋白大亞基一樣具有多種免疫學功能。組織表達分布結果顯示EcHcL1和2在10種組織中均有表達, 且均在肝胰腺和血細胞中表達較高, 與凡納濱對蝦[26]的研究結果一致, 說明2種變體組織表達分布廣泛且在均在與免疫發生的組織或器官中表達量較高, 推測2種變體可能在免疫應答中發揮了重要作用。
研究表明圓尾鱟(Carcinoscorpius rotundicauda)凝集素CL5可產生不同的亞型來識別細菌和真菌: 與細菌(金黃色葡萄球菌、大腸桿菌)相結合的CL5的電泳圖譜大致相似, 而與真菌相結合的CL5的電泳圖譜與細菌圖譜比較出現較大的差異[27]。Destoumieux等[28]研究指出, 斑節對蝦可產生5種不同的抗菌肽, 其中抗菌肽3不僅具有抗真菌活性, 還可以抑制大腸桿菌E. coli363和細菌滕黃微球菌(Micrococcus luteus)的生長。Woramongkolchai等[29]研究表明了斑節對蝦感染WSSV后抗菌肽5的mRNA水平在24h后明顯上調, 提示其可能具有抗病毒活性。
由上可知, 存在于甲殼動物中的多態性免疫分子, 其不同變體可能具有不同的免疫活性。由此推測脊尾白蝦血藍蛋白大亞基的不同變體也可能具有不同的免疫學功能。為此, 我們對脊尾白蝦進行了病原脅迫實驗, 進一步運用Real-time PCR策略對2種變體的mRNA表達情況進行了探討。結果由圖 5—8可知, 在3種病原菌脅迫下,EcHcL1在肝胰腺中表達量趨勢為先上升達到峰值后下降到與對照組無顯著性差異。EcHcL2表達量趨勢為先上升再下降再上升達到峰值, 最后72h下降到正常水平。與Yang等[30]報道的鰻弧菌感染中國明對蝦后不同時間點FcToll的表達變化情況相同。這樣的表達變化模式可能是在病原脅迫的初期, 機體對于外界刺激, 做出最初的應激反應, 出現較弱的表達上升(如6h的表達上升), 此種變化在12h恢復至原始水平, 但是, 最初較弱的表達上升, 不足以抵御病原的刺激, 在24h出現第二次表達上調, 來加強對病原菌的免疫防御。當脊尾白蝦受到金黃色葡萄球菌脅迫后, 在肝胰腺和血細胞中EcHcL1均在12h出現表達量最高的情況(約為對照組的2—3倍), 說明相對其他兩種菌, EcHcL1對球菌做出較早的免疫反應(副溶血弧菌脅迫后,EcHcL1在肝胰腺和血細胞中分別在24h和48h達到峰值; 在WSSV感染后,EcH-cL1在肝胰腺和血細胞中分別在12h和48h達到峰值)。當副溶血弧菌脅迫和WSSV后, 在肝胰腺和血細胞中EcHcL2均在24h出現峰值(約為對照組的2—9倍), 相對于球菌,EcHcL2對弧菌和WSSV做出更明顯的免疫反應(球菌脅迫后,EcHcL2在肝胰腺中48h達到峰值, 在血細胞中表達量沒有明顯上調趨勢)。綜上表明脊尾白蝦血藍蛋白大亞基變體與血藍蛋白大亞基一樣, 其mRNA表達變化與病原脅迫密切相關, 可能具有抗菌抗病活性, 且不同變體對不同病原的響應程度存在差異性, 提示可能不同的血藍蛋白變體在對蝦免疫防御中具有不同的最佳免疫學功能。
綜上, 血藍蛋白大亞基不同變體序列共同組成脊尾白蝦特有血藍蛋白超家族, 且這些變體具有相應的最佳免疫防御功能, 共同參與脊尾白蝦的免疫防御, 對豐富脊尾白蝦血藍蛋白的分子多態性與功能多樣性具有重要的意義。
