雜草對合成生長素類除草劑的抗性
2019-01-05 05:49:44編譯
世界農藥 2018年6期
葉 萱 編譯
(上海市農藥研究所,上海 200032)
合成生長素類除草劑(SAH,HRAC分類體系的O 組)模擬了天然植物激素吲哚-3-乙酸(IAA)的作用。第1個這種作用機制的除草劑2,4-滴已被廣泛使用70多年。SAH主要用于選擇性防治禾本科作物田中的闊葉雜草,但二氯喹啉酸和氯氟吡啶酯防治一些禾本科和沙草科雜草。SAH被分為7種:⑴苯氧羧酸類;⑵ 苯甲酸類;⑶ 吡啶羧酸類;⑷ 吡啶氧羧酸類;⑸ 喹啉羧酸類;⑹ 嘧啶羧酸類和⑺芳基吡啶甲酸類(arylpicolinate)(表1)。每一種都具有獨特的化學結構(圖1)。自1945年2,4-滴引入后SAH就一直被商業化使用到現在,在2018年氯氟吡啶酯引入(表 1)。2,4-滴引入用于農業給雜草管理帶來了革命,持續性創新發現和開發了數個新穎的SAH。
SAH的全球施用面積僅次于 ALS抑制劑(508×106hm2)和 EPSP 合酶抑制劑(477×106hm2)除草劑,位列第三(366×106hm2)(信息來自陶氏益農,2014年)。除草劑2,4-滴的全球使用面積為161.7×106hm2,是使用最廣泛的SAH,其次依次為麥草畏(50.0×106hm2)和 2-甲基-4-氯苯氧乙酸(MCPA)(31.3×106hm2)(圖 3)。
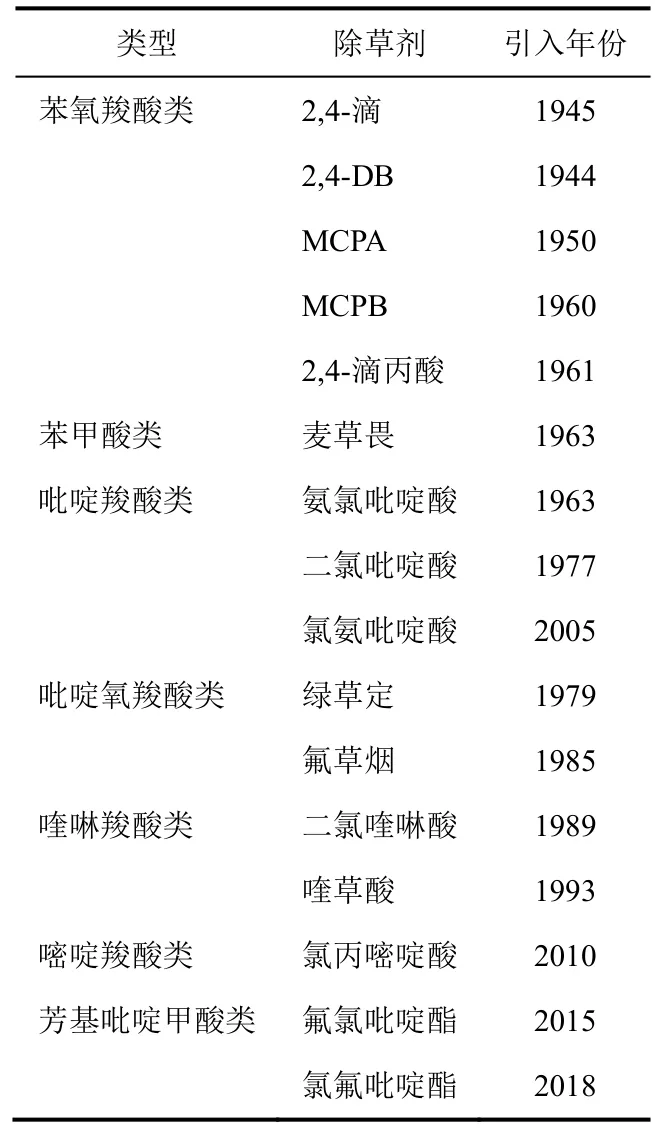
表1 不同種類合成生長素類除草劑
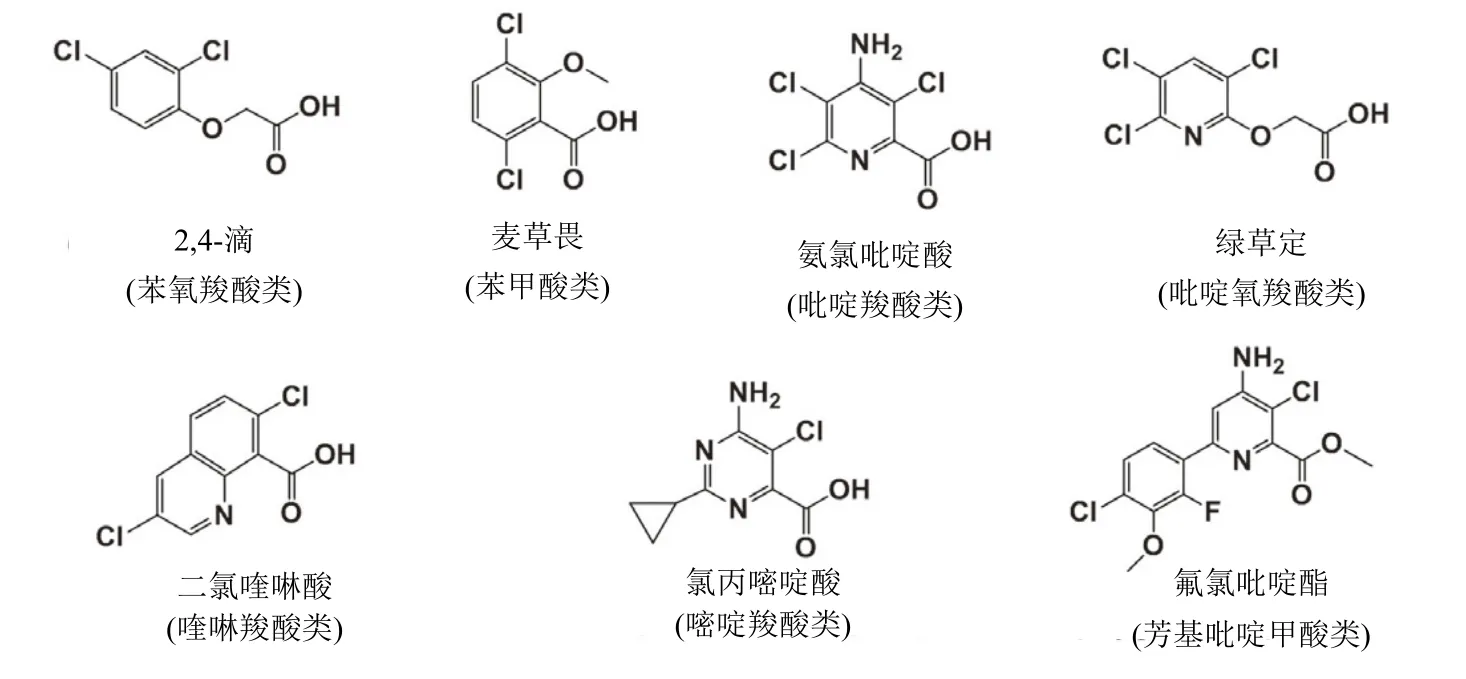
圖1 7種類別合成生長素除草劑代表性產品結構
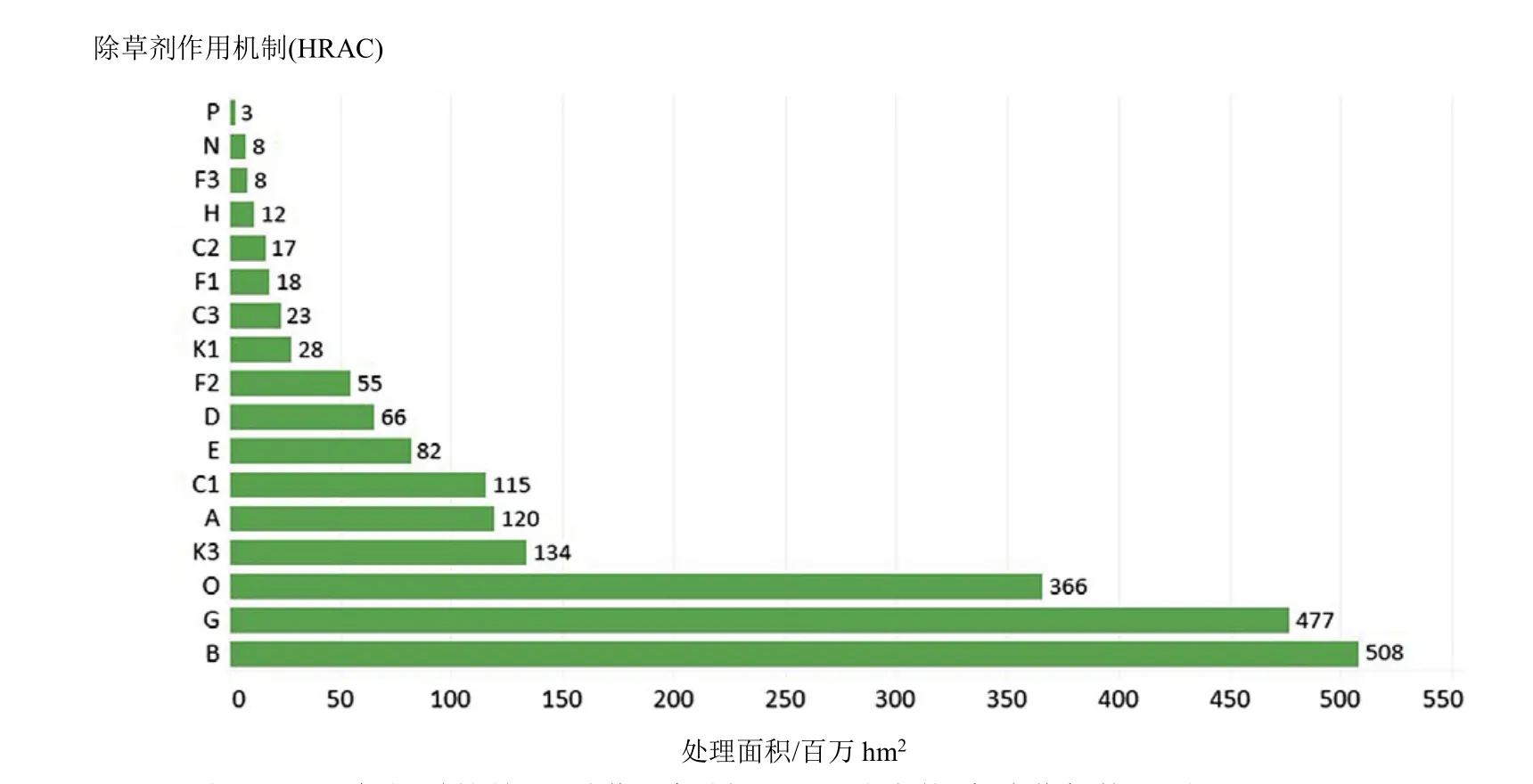
圖2 2014年報道的前17種作用機制(HRAC分類體系)除草劑使用面積(×106 hm2)
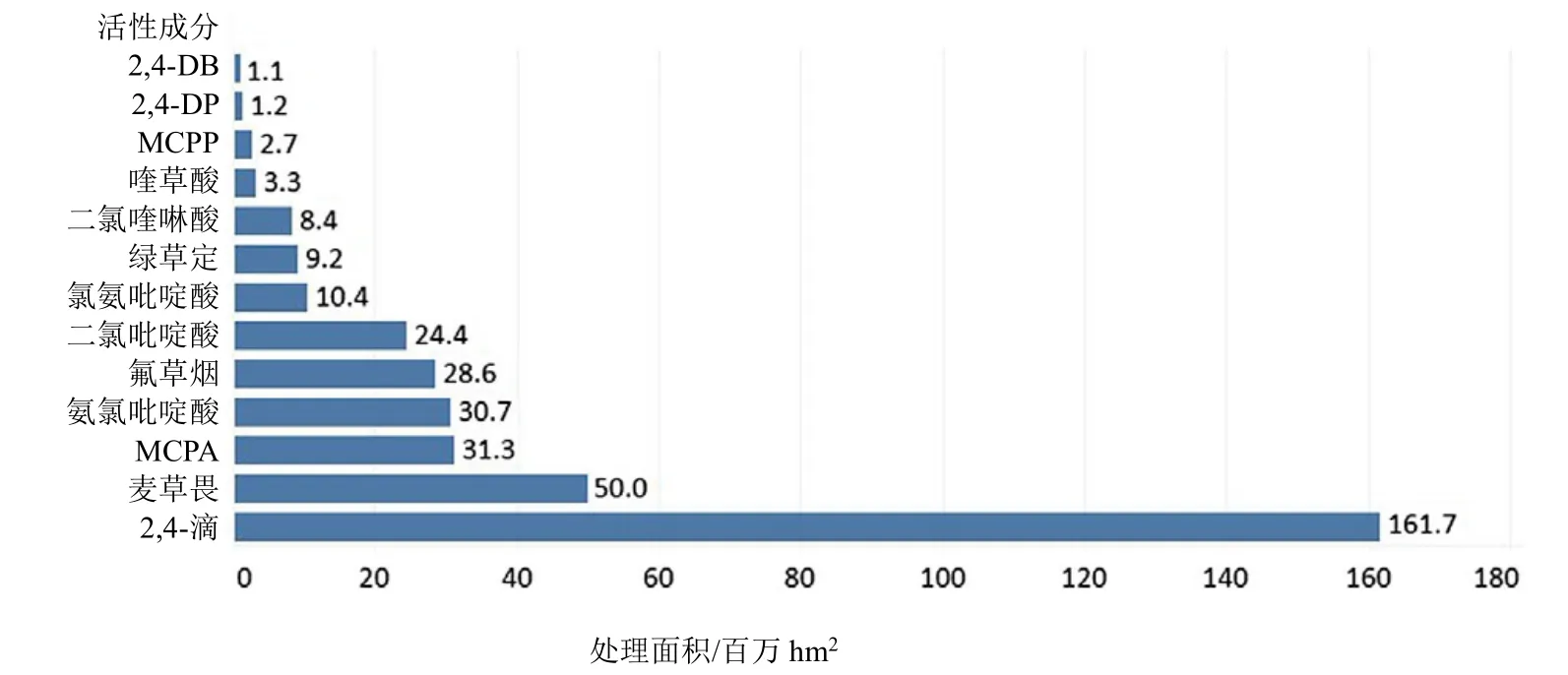
圖3 2014年報道的特定合成生長素類除草劑活性成分施用面積(×106 hm2)
為了持續應用 SAH,掌握雜草對SAH的抗性機制、抗性選擇和進化知識非常重要。在2017年5月在美國科羅拉多州的丹佛舉辦的“全球除草劑抗性挑戰”會議期間,進行了“雜草對SAH抗性”的專題討論會。討論會的主題有雜草品種對SAH抗性的進化;熟知的雜草品種對 SAH抗性進化的機制;SAH的感應、運輸和代謝;SAH發現的最近創新;耐SAH作物的抗性管理。
1 雜草品種對SAH的抗性
在1957年,首例2,4-滴抗性被報道,即在夏威夷節節草(Commelina diffusa)和在加拿大胡羅卜(Daucus carota L.)對2,4-滴產生了抗性。根據“國際抗性雜草調查”,現在有36種抗SAH的雜草品種(30種闊葉雜草、5種禾本科雜草和1種類似禾本科雜草)(圖4)。包括止血馬塘(Digitaria ischaemum)和4種稗屬雜草[稗草(Echinochloa crus-galli)、孔雀稗(Echinochloa crus-pavonis)、西來稗(Echinochloa zelayensis)和芒稗(Echinochloa colona)]的5種禾本科雜草已對二氯喹啉酸產生了抗性。有人提出其為氰化物介導的抗性機制,不同于SAH的。
對SAH產生抗性的經濟重要性雜草有澳大利亞抗2,4-滴和抗MCPA的野蘿卜(Raphanus raphanistrum)、歐洲抗苯氧除草劑的虞美人(Papaver rhoeas),加拿大和美國抗麥草畏的地膚(Kochia scoparia),美國抗 2,4-滴、麥草畏和MCPA的刺萵苣(Lactuca serriola)。此外,內布拉斯加州和伊利諾斯州的長芒莧(Amaranthus tuberculatus)生物型和阿根廷的綠穗莧(Amaranthus hybridus )被確定對一些SAH有抗性。經濟重要性稍差的抗性雜草有加拿大和美國抗2,4-滴的胡羅卜,新西蘭抗 2,4-滴的麝香飛廉(Carduus nutans)和意大利薊(Carduus pycnocephalus),以及在加拿大對SAH多抗的野芥(Sinapis arvensis L.)和抗二氯喹啉酸的豬殃殃(Galium spurium)。
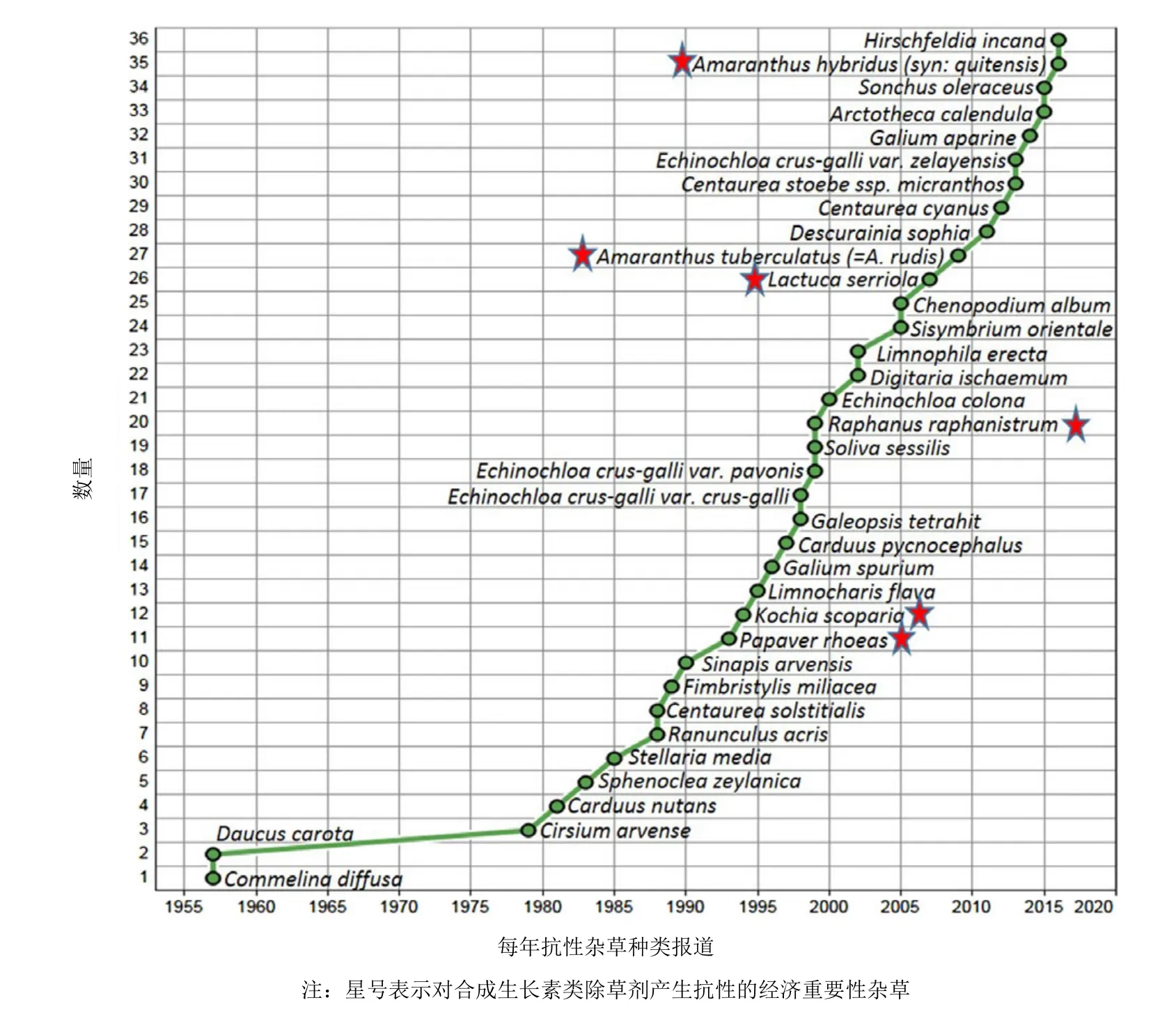
圖4 到2017年已報道對合成生長素類除草劑產生抗性的雜草品種
與其他作用機制除草劑特別是乙酰基輔酶A羧化酶(ACCase)和乙酰乳酸合酶(ALS)抑制型除草劑相比,考慮到廣泛使用SAH所形成的選擇壓程度,其抗性發生程度較低(圖5)。雖然SAH使用的時間要長于其他作用機制的除草劑,但相對來說沒有對農業生產造成廣泛的不利影響的抗 SAH雜草案例(圖 4)。
SAH抗性發生低的原因可能有以下幾種:⑴ 這些除草劑具有潛在的多個作用位點;⑵ 一些抗性事例是由隱性基因所致,其擴展比顯性性狀更慢;⑶在除草劑和作物競爭存在的情況下,抗性基因型的適合度降低。雜草交互抗性的發生可能是管理策略開發的1個挑戰。更完全地了解SAH的抗性機制應該能改進抗性管理的措施和更好地應用這些除草劑。
2 一些雜草品種對SAH的抗性機制
2.1 虞美人
虞美人是歐洲冬季谷物中最常見的闊葉雜草。此雜草品種專性異花授粉,種子產量高,種子庫具有高度持久性,適合種子萌發的時間長,故難以防治。隨著除草劑抗性的出現和擴展,虞美人正成為更難防治的雜草,特別是在歐洲的南部。最近10年在西班牙、法國和希臘,報道出現抗苯氧羧酸類(2,4-滴和 MCPA)和抗乙酰乳酸合酶抑制劑(苯磺隆)的虞美人生物型。在西班牙,也可能存在對其他苯氧羧酸類、苯甲酸類或吡啶羧酸類有交互抗性的生物型。很少有人研究虞美人對SAH的抗性機制和基因。
抗性植物中 2,4-滴遷移的缺少可能是抗性產生的原因。此外,用2,4-滴處理的敏感植物產生的乙烯是抗性植物的4~8倍。2,4-滴在抗性植物中可能沒有到達核蛋白受體復合體,壓制了生長素反應性基因,其中一些基因負責乙烯的產生。乙烯的積累可能抑制光合作用,產生H2O2和反應性氧,導致植物死亡。
將2,4-滴用于遷移性下降的抗性虞美人種群48 h后,在植物的嫩芽和根中檢測到 2,4-滴的代謝物。在處理后的 168 h,沒有檢測到 2,4-滴,只檢測到HPLC保留時間顯示為羥基化的2,3-滴和2,5-滴代謝物。用細胞色素P450抑制劑馬拉硫磷處理,能使基因型從抗性轉為敏感型。這表明細胞色素P450增加了這些抗性種群對 2,4-滴的降解。遷移性降低和除草劑降解的增加可能都是虞美人產生抗性的原因,但哪個是主要機制仍未知。一般在韌皮部的移動性比 2,4-滴更小的極性代謝物能夠降低遷移性。其他人發現在 2,4-滴降解前其遷移性就降低了。分化代謝、乙烯反應和遷移性可能是相互依賴或獨立機制,確切情況還需進一步研究。
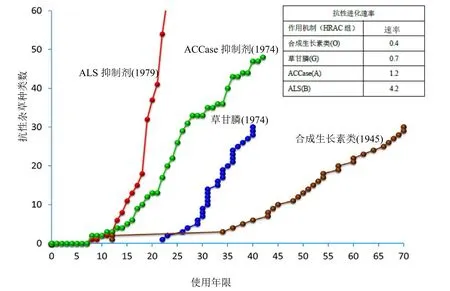
圖5 自引入后到2017年期間ALS抑制劑、ACCase抑制劑、草甘膦和合成生長素類除草劑的抗性雜草品種的數量和雜草抗性相對發展率排序
有能夠降解甲氧咪草煙的對ALS抑制型和苯氧羧酸類除草劑有抗性的虞美人生物型。是否相同的細胞色素P450降解這些作用機制的除草劑,或是否其他酶參與其中,這些仍未知。發現除草劑 2,4-滴能誘導敏感和抗性虞美人表達糖苷水解酶(GH3)和谷胱甘肽S-轉移酶(GST3),因此這些酶表達的增加好像與 2,4-滴的抗性沒有關系。需要確定虞美人抗除草劑的基因和更充分地了解其抗性機制。
2.2 地膚
地膚為侵染北美大平原作物地和非作物地的一年生夏季雜草。它出現于早春。雌蕊先熟生物學確保了高水平的遠交,導致高的遺傳變異性。成熟的種子沒有硬的外殼,很少休眠。地膚對莠去津、麥草畏、草甘膦和數種ALS抑制型除草劑產生了抗性。
抗麥草畏地膚被首次發現于20世紀90年代的科羅拉多州和內布拉斯加州。到2016年,抗除草劑生物型已普遍,特別是在科羅拉多州和堪薩斯州的休耕小麥田中。到目前為止,在科羅拉多州還沒有發現對氟草煙產生交互抗性,雖然報道蒙大拿州有。在25年中使用先進的選擇技術,培育了抗性地膚品系 9425,與敏感品系 7710相比其對麥草畏的抗性增加了30倍。性狀為顯性或半顯性。與敏感品系相比 9425品種對麥草畏的遷移性降低了。RNA-序列表明品系 9425體內影響生長素運輸的一組獨特基因被下調了,這可能是導致抗性植物中麥草畏遷移性下降的原因。
已發現少量的抗氟草煙地膚種群。雖然氟草煙是有效的可靠的防治各種環境中地膚的除草劑,但由于其雜草防治譜窄,不被看作是“可獨立使用”的產品。由于地膚對其他除草劑的抗性廣泛,而用氟草煙防治地膚日益增加,管理地膚對氟草煙抗性的進化將非常重要。
2.3 刺萵苣
刺萵苣是菊科一年生、冬性一年生或二年生雜草,是美國西北部的一大問題雜草。此植物具有深的主根,能在干旱條件下存活。刺萵苣除了與作物競爭水分、空間和營養外,其產生的黏白乳膠能堵塞收獲設備,增加谷物濕度。在收獲過程中與谷物混合的花蕾難于去除,降低了谷物的品質。
美國西北部種植的約80%小麥在生長過程中至少被施用1種SAH 1次。在2004年夏天在美國華盛頓普爾曼附近,應用840 g/hm2草甘膦和540 g/hm22,4-滴 2次后刺萵苣還能夠存活。此外調查發現對2,4-滴的差異反應可遺傳,防治失敗不是草甘膦抗性所致。在被處理后3周抗性植物從花冠再次長出的部分被用 2,4-滴處理后 2~3周受到傷害。抗性水平是敏感生物型的9~10倍。此2,4-滴抗性生物型似乎對苯氧羧酸類和苯甲酸類包括MCPA和麥草畏在內的其他SAH已發展交互抗性。
刺萵苣對 2,4-滴抗性的遺傳性受單獨共顯性基因控制。進行了環標記的14C-2,4-滴生物動力學研究來了解2,4-滴抗性的機制。在應用后96 h,抗性生物型吸收了較少的 2,4-滴,與敏感生物型相比,處理葉中有較多的放射活性。發現在處理的抗性和敏感生物型植物葉或花冠中2,4-滴的代謝率沒有差異。
在處理后前4 d,敏感生物型的植物生長受到了刺激,抗性生物型沒有。4 d后,敏感生物型的生長幾乎停止,而抗性生物型繼續生長。抗性生物型持續生長可能是生長素信號傳導反應發生改變所致的降解下降引起。也許,抗性生物型體內生長素信號傳導的改變和正常生長的維持可能已降低了抗性刺萵苣生物型對 2,4-滴的吸收和遷移。還沒有測定基因精確的作用。轉錄物組學和靶標基因表達研究可能有助于確定引起抗 2,4-滴刺萵苣生長素信號傳導反應改變的潛在基因。
2.4 野蘿卜
野蘿卜是澳大利亞南部種植系統中最成問題的雙子葉雜草,每年給種植者造成的作物產量損失至少達5700萬澳元,增加了雜草防除費用。在澳大利亞西部,此雜草已對ALS抑制型除草劑的磺酰脲類和咪唑啉酮類產生了廣泛的抗性,種植者要依賴SAH特別是2,4-滴來防除。在1999年,首次確定野蘿卜在田間對 2,4-滴發展了抗性的事例。隨后在2003、2010和2015年進行的隨機雜草種群調查表明對2,4-滴產生抗性的植物種群比例從2003年的60%增加到2010年的74%。在2015年抗性水平保持沒變。
在2001和2002年從田間收集的2個野蘿卜種群的表型特征表明與敏感種群相比抗性增加了約20倍。賦予這2個種群2,4-滴抗性的性狀為細胞核遺傳和不完全顯性。14C-標記的2,4-滴從2002年種群和 2010年收集的種群的被處理葉中遷移出的能力很弱。對敏感植物應用生長素外排抑制劑1-N-萘基酞氨酸能模仿 2,4-滴遷移受限,在抗性植物中 2,4-滴遷移受限可能是由于ATP-結合盒型B型(ABCB)長距離生長素外排運輸蛋白功能損失所致。野蘿卜的主要抗性機制好像是2,4-滴遷移的下降。
從1999、2001和2002年種群收集的2,4-滴抗性生物型也對MCPA有抗性。對1999年種群中抗MCPA生物型的研究表明其抗性遺傳方式相似于抗2,4-滴生物型的。在測定苗根延長的試驗中,2002年種群對2,4-滴、MCPA、2甲4氯丙酸、麥草畏和1-萘乙酸有抗性,而2010年種群只對苯氧乙酸、2,4-滴和MCPA有抗性。這表明抗性生物型間存在生長素感知和/或信號傳導的差異。對 2002年種群進行的全基因組轉錄組學研究表明生長素誘導的轉錄抑制因子和防御基因迅速下調,此現象在敏感種群中沒有發現。這可能是第二種抗性機制,因為一些對2,4-滴具有高度抗性的種群如敏感植物一樣能有效地從被處理葉中遷移出14C-標記的2,4-滴。
用推薦劑量 2,4-滴處理的和小麥競爭生長的植物與用 2,4-滴處理但沒有競爭生長的植物相比,其生物量減少了75%。這可能有助于確定交互抗性水平和除草劑遷移性變化,和/或抗性基因表達水平不同的植物種群在沒有進行除草劑處理的情況下的適合度。用此信息能夠識別適宜的非化學措施,如早期作物種植,應用高競爭性的作物品種,這有助于更有效的抗性管理策略。
有一些化合物能夠阻礙基因或基因產品改變抗性植物對 SAH的反應或再次激活除草劑的正常遷移能力,發現這些替代性分子可能延長SAH防治作物田中野蘿卜的時間。到目前為止,沒有發現抗性野蘿卜對 2,4-滴解毒代謝的證據(細胞色素 P450依賴或獨立型)。需要持續檢測種群進化代謝抗性,來調整抗性管理策略,更好防治這些雜草。
3 SAH的感知、運輸和代謝
3.1 生長素類除草劑的潛在靶標位點抗性機制:生長素受體
對于SAH,抗性候選機制可能包括靶標位點的改變(生長素受體或特異性生長素轉運體)和如代謝SAH的其他轉運體和酶的非靶點機制。除草劑抗性的1個主要來源為靶標位點抗性。靶標位點抗性頻繁產生于除草劑結合的蛋白質活性位點的突變,使蛋白質對一些除草劑不敏感。田間應用SAH產生抗性還沒有被追溯到核心信號傳導靶標位點。然而,許多這些蛋白的突變可對SAH產生抗性,如對SAH有耐性的擬南芥突變篩選確定大多數生長素信號傳導和運輸蛋白。
科學家已確定許多生長素受體蛋白,而受體TIR1為典范。對于擬南芥,TIR1為6個受體組成的家族的一部分,其他 5個為生長素 F-盒蛋白(AFB1-5)。此家族由3對受體組成:TIR1和AFB1,AFB2和 AFB3,AFB4和 AFB5。所有受體間具有很高程度的序列相似性,很高程度的功能冗余。其中任一基因功能突變所致植物基因型與野生型相比幾乎沒有變化,只有疊加突變才使植物的生長和發育受到影響。
TIR1受體結構通過結晶化測定。通過研究TIR1結合生長素和共受體蛋白 AUX/IAA的特性了解TIR1的作用機制(圖 6)。通過篩選大量的生長素和類生長素分子已開發了藥效圖,這些圖能給出每個受體的活性配體的詳細特性。
TIR1和AFB1-5生長素受體的序列幾乎不變,在植物界有高度的序列保守性,特別是結合位點的氨基酸。這表明1處突變會使植物的適合度下降,任何與野生型的偏離都是不利的。如果這是正確的,那么靶標位點抗性不可能產生。受體家族的過多就會使植物系統靈活,因此一個受體效力的減少可被其他補償。適合度的降低只存在于對內源性生長素IAA敏感的情況。因此,了解是否TIR1家族存在初期階段抗性和是否受體家族不同成員對不同 SAH有不同選擇性很重要。有證據表明AFB5和TIR1對不同SAH的選擇性不同,AFB5受體是吡啶甲酸除草劑的主要作用位點,也有證據表明對氨氯吡啶酸不敏感的擬南芥AFB5突變品系對2,4-滴或IAA的敏感性沒變。因此,雜草種群的AFB5同源物可能發生突變,從而對吡啶甲酸除草劑產生抗性,而對內源生長素的敏感性或植物的適合度沒有改變。吡啶甲酸除草劑管理需要說明和阻止此種情況的出現。
共受體 AUX/IAA蛋白應被考慮,因為一旦生長素在識別口袋里,共受體蛋白就結合在生長素的頂部形成共受體復合體(圖6)。不同AUX/IAA與不同濃度生長素結合,共受體的突變會造成對SAH產生抗性這是可能的。基于生長素受體和共受體的靶標位點抗性仍沒有被確定,但可能存在。可能由于受體家族的過剩,不同骨架的SAH會保持活性。
3.2 生長素類除草劑的潛在靶標位點抗性機制:生長素運輸蛋白
運輸蛋白能夠賦予靶標位點和非靶標位點抗性(圖7)。植物細胞主要是通過AUXin抗性1(AUX1)載體選擇性吸收生長素。最初從對 2,4-滴抗性的植物體內分離到AUX1突變體,已知這個靶標位點易發生變化。AUX1結構仍沒有被描述,但可用煙草細胞培養進行放射性標記的 2,4-滴累積生測來測定AUX1的活性。已研究了多樣的類生長素結構,預期可得到AUX1選擇性的藥效圖。數個SAH骨架不是AUX1的底物,但化合物仍與SAH一樣有活性。所以,SAH抗性好像不可能由AUX1突變產生或質膜中AUX1豐度的降低所致。
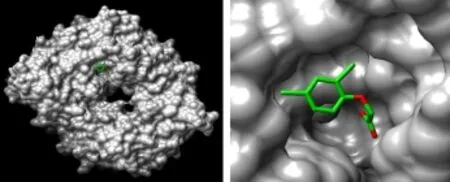
圖6 與2,4-滴結合的生長素受體TIR1的結構
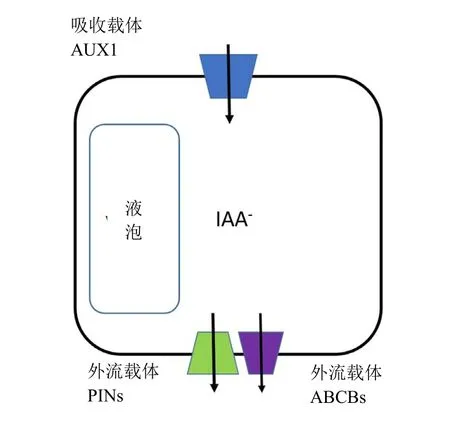
圖7 生長素運輸蛋白的排列
選擇性生長素外流產生了極性生長素運輸,使植物細胞產生極性并維持。有2個重要的外流載體蛋白家族 PIN-FORMED(PIN)和 ABCB蛋白。PIN具有植物特異性,ABCB是引起藥物抗性產生的ATP驅動轉運體家族的一部分。假如PIN和ABCB是復雜的膜結合蛋白,那么目前其結構未知。此外,沒有適于研究它們藥理學的廣泛使用的轉運生測。已知有少許選擇性運輸抑制劑,其中萘基酞基氨酸已被登記為除草劑。
更具有挑戰性的是在已有生化知識基礎上評估外流蛋白作為抗性位點的潛力。外流載體的上調可能促進底物生長素從細胞中排出,通過清除除草劑產生抗性。在考慮內源性生長素濃度和梯度時,PIN起主要作用。對PIN對不同SAH的選擇性知之甚少,但已知PIN的活性為自我增強型,因此增加生長素劑量可能補充或減少質膜中 PIN的損失而增加活性。高劑量SAH能促進PIN補充,但它們如何影響流出和抗性將取決于PIN蛋白對除草劑的選擇性。生長素輸送機制和PIN蛋白的藥理選擇性的更多知識有助于解釋PIN在SAH作用中的角色。
ABCB運輸蛋白在非植物真核表達系統中被很好地表征,目前正在不斷地了解植物中ABCB運輸蛋白的重要性。植物ABCB的初步同源模型已被提出,但還不足以精煉用于了解底物的選擇性。植物ABCB與生長素外流有特異性相關性,與依賴于它們正確的功能的物理反應有關。此家族具有許多小分子底物和質膜的ABCB豐度的變化導致對小分子藥物的抗性。更好地了解此家族植物運輸蛋白關系到避免 SAH抗性的進化。對 2,4-滴抗性因情形與ABCB轉運體活性有關,雖然導致細胞內累積量降低的受損的長距離運輸和沒有增加的細胞外排好像是抗性的基礎。
3.3 生長素類除草劑的潛在非靶標位點抗性機制:生長素的代謝
農藥的非靶標位點抗性與酶把農藥代謝為無害物質有關。使內源生長素IAA失活的主要途徑為結合。結合IAA的酶可能被認為是使合成生長素失活的候選物和抗性的潛在原因。植物體內IAA結合有3種主要途徑:通過GH3蛋白,生長素與氨基酸結合;通過葡萄糖基轉移酶與糖結合;通過 GST3與谷胱甘肽結合。已確定與IAA和水楊酸結合的GH3蛋白結構。原則上,結合酶可能是抗性產生的原因,但任何一個酶識別所有不同 SAH骨架為底物是不可能的。
通常細胞色素P450酶對小分子的氧化,可能是除草劑和包括殺蟲劑在內的其他農藥解毒的一個機制。植物對二氯喹啉酸抗性與這些酶有關。主要催化內源生長素IAA的途徑被確定為是IAA氧化酶,而不是細胞色素P450。是否此酶也將以SAH為底物還未知。雖然內源性IAA氧化酶可能是抗性產生原因,但對多種SAH的抗性是不可能的。氧化已成為最近開發耐麥草畏和 2,4-滴作物的機制。在這些情況下,細菌來源的2種不同的氧化酶被引入賦予抗性。
4 SAH發現的最近創新
在20世紀90年代晚期氯氨吡啶酸的發現促進了陶氏益農對吡啶羧酸SAH結構-活性關系的研究。這導致發現1類新穎SAH,即吡啶羧酸的衍生物芳基吡啶甲酸類。SAH的酸部分功能對其在植物韌皮部中的移動和通過離子捕獲在細胞中的累積重要。對于吡啶羧酸除草劑二氯吡啶酸、氨氯吡啶酸、氯氨吡啶酸和芳基吡啶甲酸類除草劑[氟氯吡啶酯(ArylexTMActive)和氯氟吡啶酯(RinskorTMActive)],羧酸為與TIR1和AFB1-5受體進行主要結合的功能團。芳基吡啶甲酸被制劑化為酯,這些酯在植物體內可能迅速水解為游離羧酸。
芳基吡啶甲酸具有獨特的除草活性,與吡啶羧酸除草劑相比相對低的使用劑量(相當于30 g酸/hm2或更少)防效增加。類似物DAS402除草活性顯著高于氯氨吡啶酸(圖8),但其土壤半衰期太長(大于240 d),不能滿足預期商業用途的法規要求。在DAS402結構中加入可代謝的部分,可增加其在土壤中的降解。這導致發現了 4-氨基-3-氯-6-(4-氯-2-氟-3-甲氧基苯基)-2-吡啶甲酸甲酯(氟氯吡啶酯)(圖8)。氟氯吡啶酯是活性高的廣譜SAH,對小麥和大麥高度安全,土壤半衰期為10-30 d,比DAS402的半衰期少很多。在2015年被首次商業化用于谷物。
在氟氯吡啶酯的開發過程中, 吡啶甲酸骨架的數個重要特征被確定:分子的2-羧酸和4-氨基功能團對高活性和廣譜性重要;3-氯功能團進一步增強了除草活性。對氟氯吡啶酯的吡啶甲酸部分5-位的SAR的研究發現了另一有商業用途的芳基吡啶甲酸類除草劑。氟氯吡啶酯的5-氟類似物不但具有高活性和廣譜性,而且對水稻有優異的選擇性。其相對應的羧酸[4-氨基-3-氯-6-(4-氯-2-氟-3-甲氧基苯基)-5-氟-2-吡啶甲酸]的土壤半衰期為 10~30 d。進一步優化開發了氯氟吡啶酯,用于水稻和其他作物田防治一年生禾本科雜草稗草和一些莎草科雜草(莎草屬)。在2017年首次獲批登記。
芳基吡啶甲酸除草劑的發現有力地表明有許多機會來開發新的SAH。芳基吡啶甲酸活性的多樣性使SAH更廣泛地用于雜草管理市場,如谷物、水稻、牧場和道路用地。
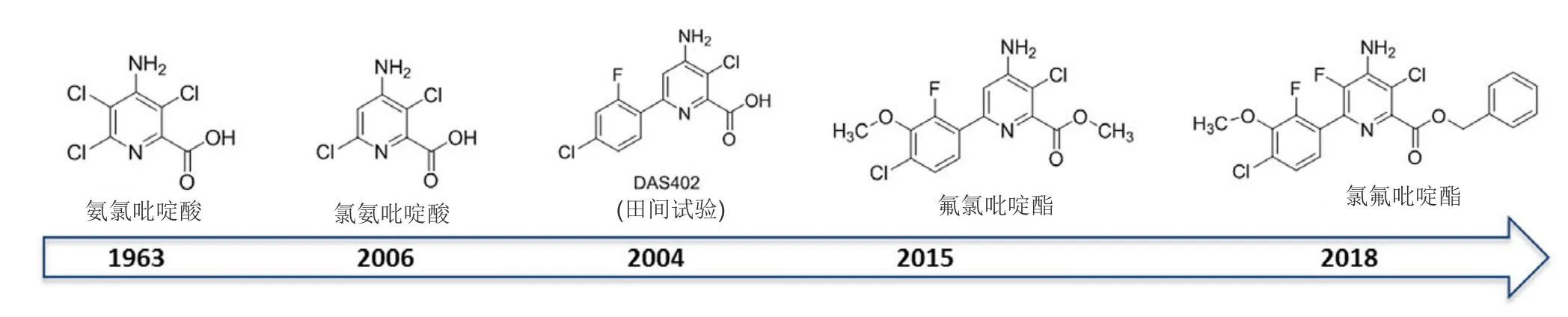
圖8 在相隔43年中發現了氨氯吡啶酸和氯氨吡啶酸
5 管理耐SAH作物的抗性
已開發了新的除草劑制劑用于陶氏益農最近開發的耐 2,4-滴作物和孟山都開發的耐麥草畏作物。Enlist DuoTM(2,4-滴-膽堿+草甘膦)和 Enlist OneTM(2,4-滴-膽堿)除草劑被開發用于EnlistTM雜草防治系統和采用VaporGrip?技術生產的Xtendimax(麥草畏)被用于Roundup Ready?Xtend作物系統。陶氏益農和孟山都已開發了管理項目以確保這些耐除草劑作物系統能被長期持續地使用,為把其整合入除草劑抗性管理項目提供指導。此外,美國環保局(USEPA)正在制定規章制度,強調需要種植者采用最好的除草劑抗性管理措施以進一步持續使用這些作物系統。
隨著抗SAH作物的引入,從監管和管理角度廣泛考慮了除草劑的抗性管理。這些工具將有助于進行雜草管理項目和防治一些抗除草劑雜草品種。廣泛采用的延緩雜草種群除草劑抗性選擇和管理現有抗性雜草種群的最好的管理措施形成管理這些技術的基礎。這些最好的管理措施是美國USEPA規定的登記條件的重點。
從20世紀50年代到2001年,在美國,除草劑登記者必須滿足有關除草劑抗性管理的少許要求。在 2001年后登記者被要求用不利影響報告程序(Adverse Effects Reporting process)向USEPA報告已確認的抗除草劑雜草品種。此外,USEPA表明登記者可能自發地在除草劑產品標簽上表明作用機制,標明除草劑抗性和阻止或延緩除草劑抗性進化的最好管理措施。在2014年,USEPA首次發布了Enlist Duo除草劑登記條件,要求要具有除草劑抗性管理計劃。在2016年,發布了Xtendimax (采用Vaporgrip技術生產)除草劑的相似批準條件。這些 SAH雜草管理系統的 USEPA除草劑抗性管理計劃由田間檢測和補救,培訓和信息,評估,最好的管理措施和報告組成。
需要利益相關者間加強合作來開發和實施有效的除草劑管理項目,限制田間抗性進化,確保除草劑能被長期持續地使用。同時除草劑開發者和登記者、農民、管理者、推廣者、專家、作物顧問、經銷商和零售商也要進行技術管理。行業鼓勵雜草管理相關利益方繼續促進把除草劑作為綜合的雜草管理方法的一部分,通過延遲田間除草劑抗性進化使這些有價值的工具被繼續使用。
6 總 結
本文目的是介紹SAH的植物抗性知識,為未來開發SAH產品帶來更好的機會。SAH藥理學獨特、復雜。其復雜的作用機制,對農業生產有益的影響和持久性是其用于雜草管理體系的自白書。
SAH在商業使用 70多年后可被繼續應用,部分原因為它們一般不被作為雜草防除的唯一工具。最近對SAH的作用機制和作用位點的觀點,有過剩的生長素受體、生長素反應型基因的改變和多個生化途徑的動態平衡的中斷,這說明不能以簡單假說解釋許多田間抗性生物型產生的原因。這也說明靶標位點抗性發展慢的原因,以及可能和適合度代價有關。最近采用最新分子和生化工具進行的新抗性雜草生物型的調查為未來抗性風險(特別是非靶標位點抗性機制)提供了信息和為新 SAH提供了潛在的延緩風險的策略。
目前也難以證實簡單的作用位點改變導致對SAH親和性下降而產生抗性。代謝增加和植物吸收降低的假設常不足以解釋生物型對SAH的反應。在對化學、蛋白質和基因調節間相互作用的了解下和應用改進的分析工具,現在了解了運輸的降低可能為 SAH抗性產生的潛在原因,增加的代謝(例如虞美人)可能或不可能(例如刺萵苣和野蘿卜)與 SAH抗性有關。對這些機制每一種影響的了解已證明是困難的,因為在復雜生長素反應途徑中存在多個和冗余的變量。
在筆者收集的觀點中,由于靶標位點(結合蛋白,TIR1/AFB1-5))改變導致的抗生物型的風險相對低。筆者也認為生長素感知和反應途徑的復雜相互作用通過靶標蛋白的過度表達(抗草甘膦地膚和長芒莧)形成的抗性可能性也低。由于 SAH的使用繼續增加,抗性選擇風險將也增加。筆者認為螯合作用和/或抗性運輸機制的改變(例如 ABCB轉運體,其他)可能發展和擴展。需要重點關注導致對其他除草劑作用機制共進化抗性的對 SAH運輸改變的性狀的選擇,這在多抗藥細菌已發現。SAH的差異代謝也已在一些雜草生物型發現,了解這些抗性機制對開發和實施更有效抗性管理策略重要。
通過增加知識和分析能力可能獲得這些見識。但是,對細胞的藥理學和植物體內長距離運輸機制了解的少。雜草科學和聯合學科為更好地了解這些機制提供了機會。在使用70多年后,具有獨特的雜草防治譜和不同化學特性新 SAH繼續被發現的事實支持此觀點,即未來有很多機會來進一步對SAH進行創新。
另一經驗為整個雜草防除專家群體包括行業科學家、學術界、管理機構和農民需要意識到和充分支持實施最好的除草劑抗性管理措施。“多樣性”是維持有效長期雜草管理的最主要的原因。例如合用不同作用機制的土壤殘留除草劑和芽后除草劑以及非化學雜草管理方法(即覆蓋作物、策略耕作、作物輪作和收獲時雜草種子防治措施)將可能會使雜草管理工具切實可行,能使現代農業蓬勃發展,為后代提供安全的糧食。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
營銷界(2015年23期)2015-02-28 22:06:18
現代農業(2015年5期)2015-02-28 18:40:49
中國質量與標準導報(2013年8期)2013-03-11 19:54:07
