sFRP2和Wnt/β-catenin通路在結直腸癌發(fā)生發(fā)展中的作用
2018-12-14 11:59:30史友權崇楊湯東熊清泉黃玉琴周懷成張琪金芝祥5王道榮
中華結直腸疾病電子雜志 2018年6期
關鍵詞:實驗
史友權 崇楊 湯東 熊清泉,4 黃玉琴,5 周懷成 張琪 金芝祥,5王道榮
近年來結直腸癌發(fā)病率愈來愈高,在全球腫瘤中發(fā)病率位于第三位,致死率位于第二位[1]。有研究認為 sFRP2(Secreted Frizzled-related proteins 2)表達減少或消失,導致與Wnt蛋白競爭性結合減少,Wnt/β-catenin通路被激活,β-catenin異常表達從而促進結直腸癌的發(fā)生發(fā)展[2-3]。目前關于sFRP2對結直腸癌細胞株HCT116細胞的增殖、遷移、侵襲等研究鮮見相關報道。我們檢測了正常結直腸黏膜組織和結直腸癌組織中sFRP2和β-catenin的表達,以及在結直腸癌細胞株HCT116轉染上調sFRP2后,對比轉染前后細胞增殖、遷移和侵襲方面的變化,以探討sFRP2和Wnt/β-catenin通路在結直腸癌發(fā)生發(fā)展中的作用。
材料與方法
一、材料
1.細胞和組織:結直腸癌細胞株HCT116來源于江蘇省蘇北人民醫(yī)院實驗中心。谷歌生物科技有限公司幫助構建包含sFRP2蛋白全長的pcDNA3.1質粒。結直腸組織標本來源于江蘇省蘇北人民醫(yī)院2015~2017年手術切除或內鏡活檢確診的標本,正常結直腸黏膜標本32例(非結直腸癌患者),結直腸癌標本32例,共64例,所有結直腸癌患者術前均未行放化療,臨床病理特征詳見表1。
2.主要試劑:兔抗人sFRP2多克隆抗體(Abcam)、兔抗人β-catenin抗體(Santa Cruz)、免疫組化試劑盒(凱基生物)、胰蛋白酶(凱基生物)、Western Blot試劑盒(凱基生物)、CCK-8試劑盒(Dojindo)、胎牛血清(杭州四季青)、DMEM培養(yǎng)基(Hyclone)、脂質體Lipofectamine TM3000(Invitrogen)、Transwell小室(Corning Corstart)和Matrigel基質膠(BD)。
二、實驗方法
1.免疫組化(SP):選取上述結直腸組織標本,經福爾馬林固定,脫水,石蠟包埋后切片。依次使用二甲苯常規(guī)脫蠟、酒精梯度復水和抗原修復后,根據免疫組化試劑盒說明書進行后續(xù)實驗。每批均設置陰性和陽性對照,陰性對照:PBS替代一抗,陽性對照:已確定的sFRP2、β-catenin表達陽性的結直腸組織。
我們根據以下標準判定sFRP2和β-catenin的染色情況:(1)sFRP2染色結果根據染色強弱和細胞陽性表達率進行綜合評分。1)染色強弱:0分(無染色),1分(淡黃色),2分(棕黃色),3分(棕褐色)。2)細胞的陽性表達率:0分(≤25%),1分(26%至50%),2分(51%至75%),3分(大于76%)。綜合兩者進行評分,依據評分判定結果,陽性(+):兩項評分之和大于3;陰性(-):兩項評分之和小于等于3分。(2)β-catenin:細胞出現黃色或棕黃色顆粒為陽性表達。膜正常表達:細胞膜染色程度大于70%;膜表達缺失:小于等于70%;胞漿或胞核陽性表達:胞漿或胞核陽性率在同類細胞中比例大于10%,兩者都稱為異位表達。異位表達和膜表達缺失統(tǒng)稱異常表達[4]。
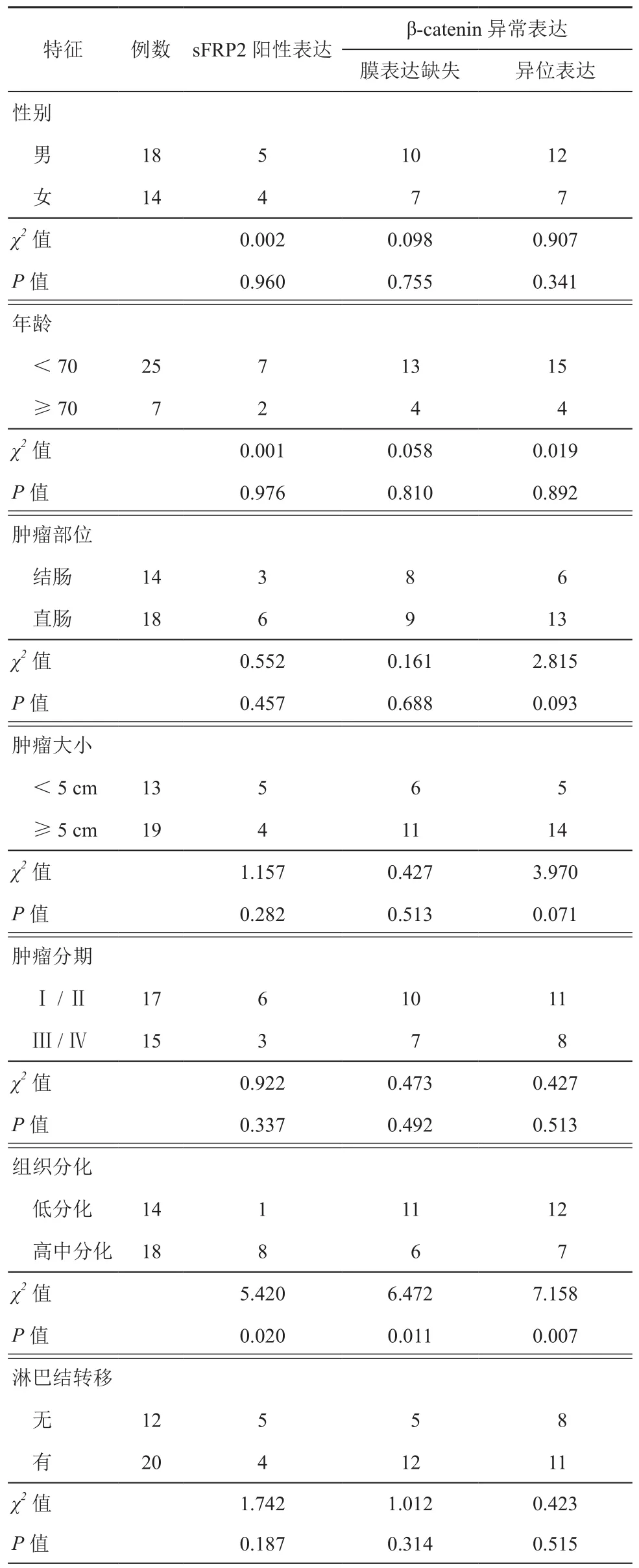
表1 sFRP2和β-catenin與結直腸癌臨床特點的關系(例)
2.細胞培養(yǎng)及轉染方法:培養(yǎng)條件為含10%胎牛血清的DMEM培養(yǎng)基+培養(yǎng)箱(37℃和5%CO2)。使用LipofectamineTM3000轉染試劑在細胞對數生長期轉染pcDNA3.1質粒。實驗分為3組:(1)對照組:未轉染。(2)空載質粒組:轉染空白質粒。(3)sFRP2轉染組。轉染過程依據轉染試劑說明書進行。
3.Western blot:細胞經過處理后,加入RIPA裂解緩沖液。首先使用SDS/PAGE凝膠電泳分離蛋白質樣品,將其轉移到PVDF膜上,配備5%脫脂奶粉封閉液,將PVDF膜上浸泡1 h,依次與兔抗人sFRP2多克隆抗體(1:1000)和兔二抗(1:1000)反應,后進行曝光顯影。
4.CCK-8實驗:將1.0×104個細胞接種于96孔板,根據CCK-8試劑盒產品說明書進行細胞計數。簡而言之,向每個孔中加入10 μL CCK-8溶液,并將它們孵育1 h,然后在450 nm下測量吸光度,實驗重復3次。
5.劃痕實驗:將HCT116細胞接種于6孔板中,融合至80%~90%時,在孔板中心軸處用槍頭沿直線輕輕劃痕,然后用PBS除去細胞碎片。無血清培養(yǎng)基培養(yǎng),在0 h、48 h進行觀察拍照。測量劃痕間的距離,實驗重復3次。
6.Transwell實驗:Transwell小室底部膜由Matrigel基質膠包被(濾膜孔徑為8 μm)。上室:無血清DMEM培養(yǎng)基(含細胞),下室:10%胎牛血清DMEM培養(yǎng)基,孵育24 h。除去上室細胞后,用多聚甲醛中固定15 min,染色20 min。隨機選取5個視野(100倍)進行細胞計數,取其平均值。
三、統(tǒng)計學分析
應用SPSS 21.0統(tǒng)計軟件。t檢驗應用于計量資料,χ2檢驗或Fisher精準檢驗應用于計數資料,Spearman分析用于檢驗相關性。組間比較選用獨立樣本t檢驗或單因素方差分析。P<0.05為差異有統(tǒng)計學意義。
結 果
一、免疫組化
1.sFRP2和β-catenin在結直腸組織中的表達:我們分別對正常結直腸黏膜組織和癌組織兩組進行了免疫組化檢測,正常結直腸黏膜組織中sFRP2主要在細胞質里表達(圖1A),β-catenin主要在細胞膜上表達(圖1C)。結直腸癌組織中sFRP2低表達或不表達(圖1B)。而β-catenin異常表達增加(圖1D)。兩組在sFRP2的陽性率、β-catenin膜表達缺失率及異位表達率等方面表達差異具有統(tǒng)計學意義(P<0.01),正常黏膜組織中sFRP2的陽性率顯著高于結直腸癌組,結直腸癌組中β-catenin膜表達缺失率和異位表達率癌顯著高于正常結直腸黏膜組。詳見表2。
2.sFRP2和β-catenin與結直腸癌臨床特點的關系:如表1所示,sFRP2陽性表達、β-catenin膜表達缺失、異位表達與腫瘤組織分化存在明顯差異(χ2=5.420,P=0.020;χ2=6.472,P=0.011;χ2=7.158,P=0.007),而與性別、年齡、腫瘤部位、腫瘤大小、腫瘤分期和淋巴結轉移無關(P>0.05)。
3. 相關性分析:通過Spearman分析,我們研究發(fā)現sFRP2表達分別與β-catenin膜表達缺失(r=-0.39,P=0.03)及β-catenin異位表達(r=-0.47,P=0.01)負相關,β-catenin異位表達與β-catenin膜表達缺失正相關(r=0.50,P=0.004),且相關性均顯著。詳見表3。
二、Western blot檢測
Western blot結果驗證了轉染后sFRP2的表達水平顯著高于轉染前(t=25.430,P=0.001),β-catenin表 達 水 平 亦 較 前 提 高(t=15.000,P=0.001),β-actin作為內參。如圖2所示。
三、CCK-8實驗
通過CCK-8方法檢測發(fā)現,sFRP2轉染組分別與對照組和空載質粒組比較[12 h:(t=0.016,P=0.988)和(t=0.041,P=0.970),24 h:(t=0.155,P=0.884)和(t=0.452,P=0.675),36 h:(t=0.695,P=0.525)和(t=1.173,P=0.306),48 h:(t=1.254,P=0.278)和(t=1.693,P=0.166),60 h:(t=3.440,P=0.026)和(t=3.576,P=0.023)],細胞增殖速度明顯減慢(圖3),說明sFRP2抑制了結直腸癌細胞HCT116的增殖。
四、劃痕實驗
通過比較sFRP2轉染組、空載質粒組和對照組三組HCT116細胞劃痕兩側直線間的距離,來判斷sFRP2對HCT116細胞遷移能力的影響。結果發(fā)現:sFRP2轉染組劃痕兩側距離與空白組、空載質粒組比較,遷移速度明顯較慢[(t=16.890,P=0.001)和(t=7.206,P=0.002)],表明sFRP2抑制了癌細胞的遷移。如圖4所示。
五、Transwell實驗
通過Transwell實驗我們發(fā)現,對照組透膜(195.39±8.68)個,sFRP2轉染組透膜(75.69±7.19)個,對照組透膜細胞數明顯多于sFRP2轉染組(t=25.459,P=0.001),如圖5所示。
討 論

圖1 sFRP2和β-catenin在結直腸組織中的的表達。1A:正常結腸組織(sFRP2);1B:結直腸癌組(sFRP2);1C:正常結腸組織(β-catenin);1D:結直腸癌組(β-catenin)(免疫組化SP法×400)
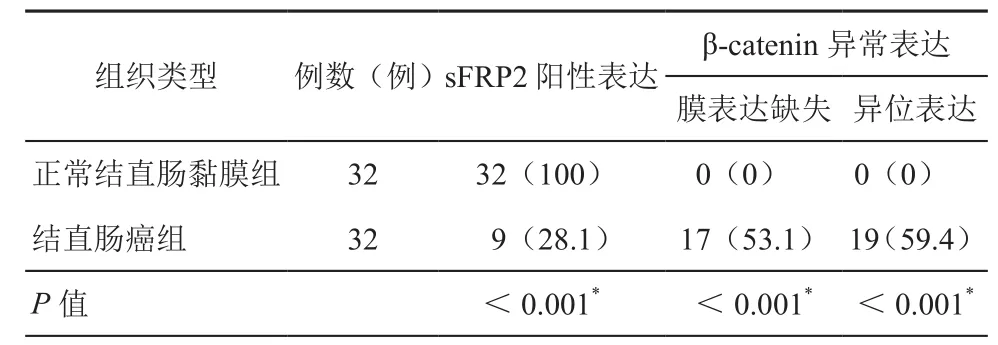
表2 sFRP2及β-catenin在兩組中的表達情況[例(%)]
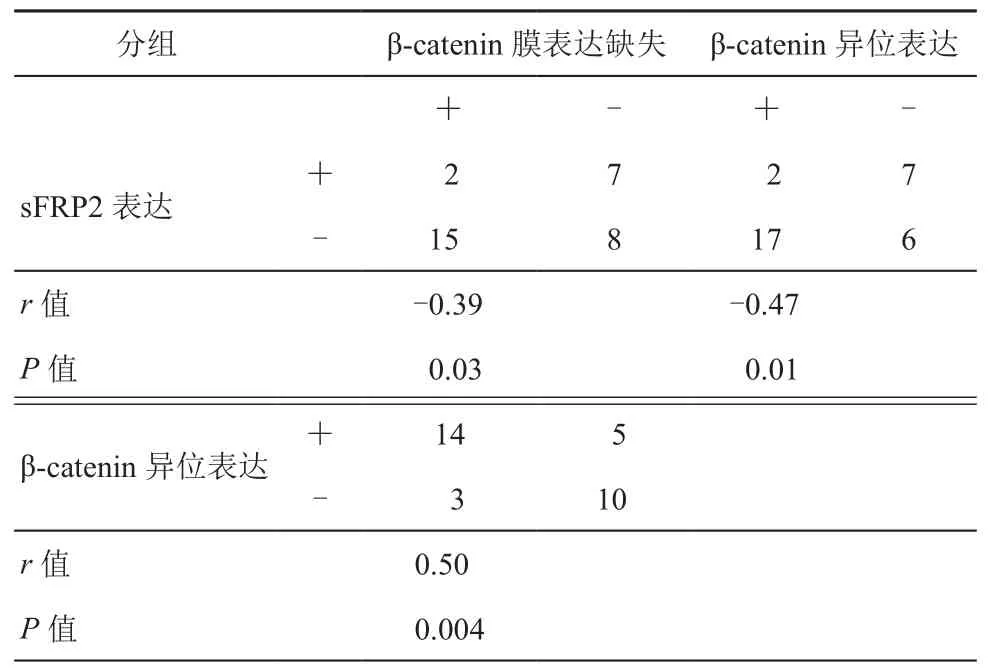
表3 結直腸癌中sFRP2和β-catenin表達的相關性(例)
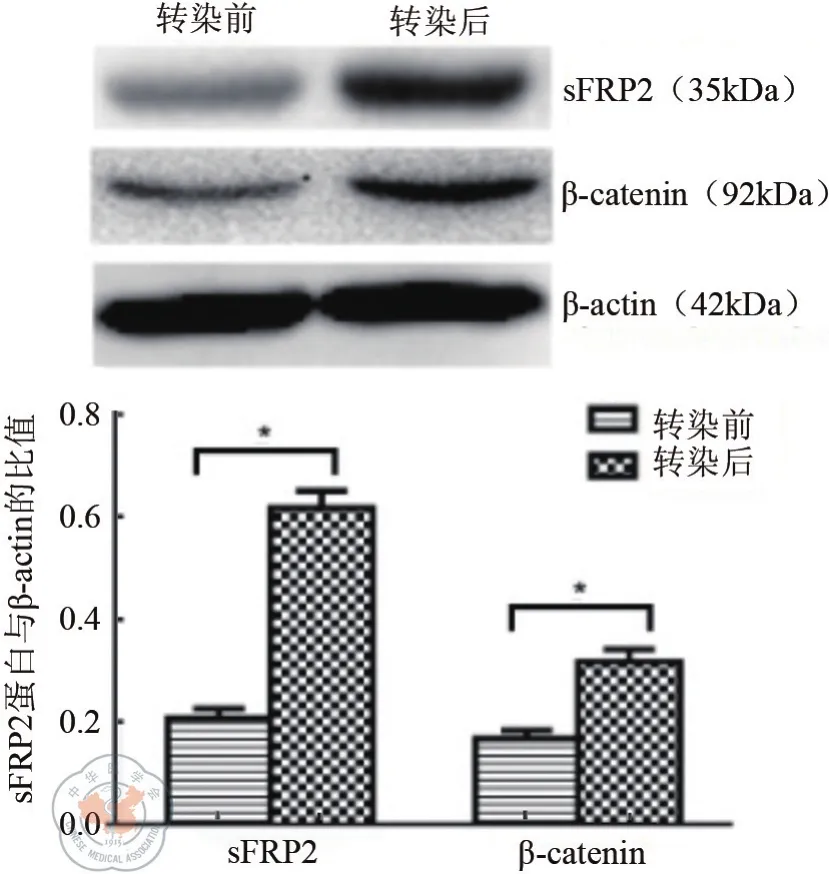
圖2 轉染前后HCT116細胞中sFRP2、β-catenin的表達(sFRP2及β-catenin轉染前后相比*P<0.05)
sFRP2是sFRP家族7個成員之一,結構上與卷曲蛋白相似,均含有半胱氨酸富集區(qū)(cysteine rich domain,CRD)。一方面,sFRP2通過CRD與Frizzed競爭結合到Wnt蛋白上抑制Wnt通路;另一方面,sFRP2與Frizzed結合生成無功能的復合物,封閉Wnt信號通路[5],從而抑制腫瘤的發(fā)生。當sFRP2表達減少時,與Wnt蛋白競爭性結合減少,不能阻斷或封閉Wnt通路,導致Wnt通路激活,便起到了促癌的作用[6-7]。我們研究發(fā)現sFRP2在正常結直腸黏膜組中的陽性表達率(100%)顯著大于結直腸癌組(28.1%),差異具有統(tǒng)計學意義(P<0.05),這與劉寧等[8]研究結果相一致。有研究報道結直腸癌中sFRP2甲基化,可導致sFRP2表達水平下降或沉默[9-11]。

圖3 sFRP2對結直腸癌HCT116細胞生長曲線的影響(sFRP2轉染組與對照組相比*P<0.05)

圖4 sFRP2對結直腸癌細胞HCT116細胞遷移能力的影響(sFRP2轉染組與對照組相比*P<0.05)
Wnt是分泌型、脂質修飾的糖蛋白,其在胚胎發(fā)育過程中具有許多關鍵作用,并促進成年人的組織平衡。它們可以調節(jié)細胞的分化、增殖、遷移、存活和干細胞自我更新[12-14]。異常Wnt信號與許多疾病相關,特別是癌癥[15]。Wnt/β-catenin通路是目前結直腸癌發(fā)生進展機制研究的熱點[16],當Wnt通路激活時,β-catenin不被降解,進入細胞核并積聚,β-catenin出現膜表達缺失和異位表達,與轉錄因子相互作用,啟動轉錄,調控基因表達,從而促進腫瘤的發(fā)生和發(fā)展[17]。我們研究發(fā)現:β-catenin膜表達缺失率在結直腸癌組(53.7%)顯著大于正常結直腸黏膜(0%),β-catenin異位表達率在結直腸癌組(61.1%)顯著大于正常結直腸黏膜(0%),且差異具有統(tǒng)計學意義(P<0.05)。這與相關研究[8]結果相一致。

圖5 sFRP2對結直腸癌細胞HCT116細胞侵襲能力的影響(sFRP2轉染組與對照組相比*P<0.05)
本研究發(fā)現sFRP2的表達和β-catenin異常表達均與組織分化明顯相關(P<0.05),而與性別、年齡、腫瘤部位、腫瘤大小、腫瘤分期及淋巴結轉移無關(P>0.05),這與相關研究[8,18-19]結果相一致。我們通過檢測發(fā)現,發(fā)現sFRP2的陽性率與β-catenin的膜表達缺失率及異位表達率均呈負相關,β-catenin的膜表達缺失率和異位表達率呈正相關,與劉寧等[8]和潘世杰[20]相關研究報道一致。主要因為sFRP2的表達減少,導致Wnt通路激活,從而引起β-catenin表達的變化。
目前,在結直腸癌細胞株HCT116中轉染上調sFRP2后研究其對細胞的增殖、遷移和侵襲的研究尚缺乏。我們研究發(fā)現sFRP2上調后細胞增殖速度顯著減慢,抑制HCT116細胞的增殖。肖燦[21]通過質粒轉染上調人舌鱗癌細胞系Tca8113細胞中sFRP2的表達,發(fā)現sFRP2的過表達顯著抑制了細胞的增殖。sFRP2表達水平下降導致Wnt/β-catenin通路的激活,進而β-catenin進入胞漿和胞核并積累,與CyclinD1啟動子中的LEF-1位點相互作用,激活轉錄過程,細胞周期蛋白依賴性激酶(CDKs)被激活后,誘導磷酸化,促進基因轉錄,促使細胞完成由G1期到S期的轉換,并進入增殖期[22-23]。所以sFRP2表達水平升高阻斷或封閉Wnt/β-catenin通路,則抑制了細胞的增殖[24]。通過劃痕實驗,我們發(fā)現sFRP2抑制了細胞的遷移能力。通過Transwell實驗我們發(fā)現sFRP2轉染組透膜細胞數明顯小于對照組透膜細胞數,說明sFRP2抑制HCT116細胞的侵襲能力。但是我們的實驗細胞比較單一,僅一種結直腸癌細胞,仍需要進一步在其他結直腸癌細胞中進行驗證。
綜上所述,sFRP2是一種抑癌基因,通過調控Wnt/β-catenin通路而發(fā)揮其作用。sFRP2結直腸癌細胞株HCT116中轉染上調后,明顯抑制了結直腸癌細胞的增殖、遷移和侵襲。希望本研究能為結直腸癌的治療提供新的靶點和思路。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
