表達(dá)豬白細(xì)胞介素2(pIL-2)重組乳酸桿菌的構(gòu)建
2018-12-05 08:59:46沙萬(wàn)里劉仲藜韋舒婉李國(guó)江
中國(guó)獸醫(yī)雜志 2018年8期
沙萬(wàn)里,閆 滿,劉仲藜,韋舒婉,李國(guó)江,2
(1.吉林農(nóng)業(yè)科技學(xué)院動(dòng)物科技學(xué)院,吉林 吉林 132101;2.預(yù)防獸醫(yī)學(xué)吉林省重點(diǎn)實(shí)驗(yàn)室,吉林 吉林 132101)
隨著國(guó)家規(guī)模化養(yǎng)殖技術(shù)的推廣以及飼養(yǎng)環(huán)境惡化程度的加劇,禽流感、豬流感等各種傳染類(lèi)疾病對(duì)養(yǎng)殖業(yè)甚至人類(lèi)的危害越來(lái)越嚴(yán)重,防治難度越來(lái)越大,傳統(tǒng)防治方法已凸顯其不足[1]。對(duì)新型免疫治療劑、免疫增強(qiáng)劑以及基因工程疫苗的需求尤為迫切。
目前,白細(xì)胞介素2(IL-2)已是一種在預(yù)防和治療疾病方面?zhèn)涫荜P(guān)注的重要細(xì)胞因子。IL-2具有廣泛生物活性,可促進(jìn)T、B淋巴細(xì)胞、NK細(xì)胞增殖與分化,促進(jìn)干擾素(Interferon,IFN)、腫瘤壞死因子(Tumor necrosis factor,TNF)等細(xì)胞因子分泌[2],提高動(dòng)物機(jī)體細(xì)胞免疫和體液免疫能力,解除機(jī)體免疫抑制,增強(qiáng)疫苗免疫效果[3],降低疫苗副反應(yīng),治療寄生蟲(chóng)感染。IL-2在抗病毒、抗感染、抗毒素、抗腫瘤及免疫調(diào)節(jié)中發(fā)揮重要作用,在動(dòng)物疾病防治中倍受關(guān)注,可作為免疫佐劑增強(qiáng)疫苗免疫效果[4]。
乳酸菌可通過(guò)口服途徑影響人類(lèi)和動(dòng)物胃腸道從而影響機(jī)體的健康,作為一種工程菌,它受到廣泛的關(guān)注并得以應(yīng)用。乳酸菌作為口服給藥系統(tǒng)及其用途已經(jīng)被不斷地開(kāi)發(fā),大部分乳酸菌在消化道可以存活并且一些可持續(xù)定植于腸內(nèi),有些菌系是人類(lèi)共生菌群的成員[5]。乳酸菌能夠遞呈生物分子通過(guò)M細(xì)胞或樹(shù)突狀細(xì)胞穿過(guò)上皮細(xì)胞。乳酸菌作為載體遞呈生物分子到腸胃道中治療疾病的研究日益成熟,包括肽療法,低聚糖和核酸的大分子口服藥物載體(或大分子的口服給藥方式)是現(xiàn)代生物技術(shù)制藥的一項(xiàng)主要挑戰(zhàn)[6]。其優(yōu)點(diǎn)是避免免疫原性的附加風(fēng)險(xiǎn)、伴隨非腸道投藥的帶來(lái)的疼痛和不適以及低廉的加工成本。將乳酸菌作為表達(dá)載體能夠遞呈生物分子至腸道中,通過(guò)與特異的細(xì)胞相互作用而發(fā)揮調(diào)節(jié)機(jī)體免疫能力[7-9]。而口服給藥系統(tǒng)又是替代非腸道給藥的有效途徑,可以減少因長(zhǎng)期用給藥病人帶來(lái)的不便和痛苦,具有廣闊的市場(chǎng)前景。本研究通過(guò)PCR的方法在pMD18-T-pIL-2質(zhì)粒上擴(kuò)增pIL-2基因,并于乳酸桿菌表達(dá)載體pSIP409鏈接,應(yīng)用重組乳酸桿菌表達(dá)豬白細(xì)胞介素2(pIL-2)。為制備新型微生態(tài)制劑防治動(dòng)物重大疫病以及降低抗生素殘留促進(jìn)動(dòng)物性食品安全具有重要意義。
1 材料與方法
1.1 載體和細(xì)胞 大腸桿菌-乳酸菌穿梭表達(dá)載體pSIP409由Senior Research Scientist Norwegian Food Research Institute贈(zèng)送;LactobacillusplantarumNC8、重組質(zhì)粒pMD18-T-pIL-2和E.coliBL21由吉林農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院動(dòng)物微生態(tài)學(xué)研究室保存;E.coliDH5α,購(gòu)自TIANGEN公司;豬腸道上皮細(xì)胞(IPEC-J2)由吉林農(nóng)業(yè)大學(xué)動(dòng)物科技學(xué)院王春鳳教授惠贈(zèng);產(chǎn)腸毒素大腸桿菌(ETEC)菌株,購(gòu)自中國(guó)獸醫(yī)藥品監(jiān)察所。
1.2 主要試劑 TRIZol、ExTaqDNA聚合酶、KpnI、HINDⅢ限制性內(nèi)切酶、dNTPs、反轉(zhuǎn)錄酶和T4 DNA連接酶,購(gòu)自TaKaRa公司;豬IL-2單克隆抗體,購(gòu)自美國(guó)BIO-RAD公司;質(zhì)粒、DNA凝膠回收試劑盒,購(gòu)自O(shè)MEGA公司;T4 DNA連接酶,反轉(zhuǎn)錄試劑盒,購(gòu)自上海捷瑞生物工程有限公司;EB購(gòu)自愛(ài)普華美生物技術(shù)(北京)有限責(zé)任公司;PVDF轉(zhuǎn)移膜,購(gòu)自Gleman公司;其他常用的主要試劑均為進(jìn)口產(chǎn)品或國(guó)產(chǎn)分析純產(chǎn)品。
1.3 主要儀器設(shè)備 PCR儀(TC-25/H,杭州博日科技有限公司);數(shù)顯恒溫測(cè)速雙功能氣浴缸(ZD-88,江蘇金壇市華城開(kāi)元實(shí)驗(yàn)儀器廠);電熱恒溫水槽(DK-8D,上海精宏實(shí)驗(yàn)儀器設(shè)備有限公司);凝膠成像系統(tǒng)(美國(guó)UVP公司,76-0312-02);水平電泳儀(DYY-6B,北京六一儀器廠);041BR 02682垂直電泳儀(美國(guó)BIO-RAD公司);高壓蒸汽滅菌鍋(西班牙Selecta公司,presoclave75);低溫超速離心機(jī)(3K30,德國(guó)Sigma公司);電穿孔儀(411BR4513,美國(guó)BIO-RAD公司);超聲波細(xì)胞粉碎機(jī)(寧波新芝生物有限公司,JY88-Ⅱ);Uvmini-1240分光光度計(jì)(島津SHIMADZU);RPMI1640細(xì)胞培養(yǎng)基,購(gòu)自上海江萊生物科技有限公司;CO2培養(yǎng)箱,購(gòu)自日本三洋公司。
1.4 重組乳酸桿菌(pSIP-409-pIL-2)載體的構(gòu)建 將重組質(zhì)粒pMD18-T-pIL-2和表達(dá)載體pSIP409用KpnI、HindⅢ限制性內(nèi)切酶進(jìn)行雙酶切。瓊脂糖凝膠電泳后,回收目的片段,用T4 DNA連接酶將回收的外源目的基因片段pIL-2與表達(dá)載體pSIP-409連接。并將重組后的pSIP-409-pIL-2載體轉(zhuǎn)化至DH5α感受肽細(xì)胞中。
1.5 重組乳酸桿菌(pSIP-409-pIL-2)載體的鑒定 選取培養(yǎng)基中生長(zhǎng)較好單菌落(疑似陽(yáng)性重組菌)加入5 mL LB液體培養(yǎng)基中,再加入50 μL Em(使培養(yǎng)基Em濃度為200 μg/mL)37 ℃搖床180 r/min震蕩培養(yǎng)過(guò)夜。提取重組質(zhì)粒pSIP-409-pIL-2 進(jìn)行PCR與雙酶切鑒定。并將PCR和雙酶切初步鑒定的pSIP-409-pIL-2質(zhì)粒送往吉林省庫(kù)美生物科技有限公司進(jìn)行測(cè)序分析。
1.6 重組乳酸桿菌的構(gòu)建 利用電轉(zhuǎn)化儀將重組質(zhì)粒pSIP409-pIL-2轉(zhuǎn)化至制備好的乳酸菌感受態(tài)細(xì)胞。均勻涂布于紅霉素抗性的MRS平板上。將生長(zhǎng)較好的重組陽(yáng)性菌單菌落接種于含有紅霉素抗性的MRS液體培養(yǎng)基中,30 ℃厭氧培養(yǎng)24 h,取800 μL菌液與200 μL 80%甘油加入1.5 mL離心管中,封口置于-20 ℃保菌。然后將剩余菌液12 000 r/min離心1 min,棄上清后向菌體加入50 μL溶菌酶混勻,并置于37 ℃的水浴中裂解30 min,提取質(zhì)粒,進(jìn)行PCR與雙酶切鑒定。
1.7 重組乳酸桿菌中IL-2的表達(dá) 將重組乳酸桿菌接種于紅霉素抗性的MRS液體培養(yǎng)基中,30 ℃厭氧培養(yǎng)24 h。將培養(yǎng)物以1∶100擴(kuò)大培養(yǎng)一代至OD600值為0.3,加入SppIP誘導(dǎo)表達(dá)5 h和7 h。調(diào)菌液至相同濃度,離心收集菌體。以上述方法誘導(dǎo)含空載體的乳酸桿菌5 h樣品作對(duì)照。制備蛋白樣品進(jìn)行12%凝膠SDS-PAGE電泳分析。
1.8 Western Blot檢測(cè)重組乳酸桿菌中IL-2的表達(dá) SDS-PAGE電泳結(jié)束后,將蛋白轉(zhuǎn)印至PVDF膜上,膜用5%脫脂奶粉室溫封閉1.5 h后,PBST洗膜4次,用PBST按照1∶1 000稀釋豬IL-2單克隆抗體,加到膜上,4 ℃過(guò)夜,次日PBST洗膜4次,再加入用PBST稀釋的HRP標(biāo)記的二抗(1∶5 000),室溫孵育1.5 h,PBST洗膜4次,用ECL顯色,掃描記錄。
1.9 免疫熒光法檢測(cè)重組乳酸菌對(duì)豬腸道上皮細(xì)胞的粘附性 取高壓滅菌后的蓋玻片,置于24孔板中,取IPEC-J2細(xì)胞每孔500 μL接種過(guò)夜,次日細(xì)胞長(zhǎng)至80%密度,吸取培養(yǎng)液,PBS清洗3次,每孔加入0.5 mL菌懸液與1640培養(yǎng)液0.5 mL,37 ℃培養(yǎng)2 h吸取培養(yǎng)液,PBS清洗3次。加入0.5 mL ETEC菌懸液與0.5 mL 1640培養(yǎng)液,37 ℃培養(yǎng)2 h,PBS清洗5次,4%預(yù)冷甲醛室溫固定20~30 min,PBS洗5次。0.2%Triton X-100作用2~5min,PBS洗5次。100 mL/L的小牛血清白蛋白室溫封閉30 min,棄上清。加入相應(yīng)抗體,放在濕盒里,4 ℃過(guò)夜,PBS漂洗3次,每次10 min,加入相應(yīng)二抗,室溫2 h,PBS漂洗3次,95%甘油封片。
2 結(jié)果
2.1 重組質(zhì)粒pSIP-409-pIL-2的PCR鑒定和雙酶切鑒定 將提取的重組質(zhì)粒pSIP-409-pIL-2適當(dāng)稀釋后作為模版,通過(guò)PCR擴(kuò)增進(jìn)行鑒定。擴(kuò)增產(chǎn)物在465 bp處出現(xiàn)pIL-2目基因條帶,與預(yù)期結(jié)果相符(圖1)。用KpnI和HindⅢ 限制性內(nèi)切酶對(duì)重組質(zhì)粒pSIP-409-pIL-2進(jìn)行酶切鑒定,結(jié)果位于5 627 bp和465 bp處出現(xiàn)載體帶和目的條帶,說(shuō)明IL-2基因與表達(dá)載體正確連接(圖2)。
2.2 目的基因的測(cè)序鑒定 吉林省庫(kù)美生物科技有限公司進(jìn)行測(cè)序分析結(jié)果,同GenBank上登陸的豬白細(xì)胞介素-2基因進(jìn)行同源性比較。結(jié)果表明,本試驗(yàn)克隆得到的pIL-2基因序列與公布的pIL-2基因序列同源性達(dá)到100%(圖3)。

圖1 PCR擴(kuò)增產(chǎn)物瓊脂糖電泳
M: DL-2 000 Marker; 1: pSIP-409-pIL-2; 2: pSIP-409

圖2 雙酶切瓊脂糖電泳
M1:λDNA/HindⅢ Marker; M2: DL-2 000 Marker; 1-6: pSIP-409-pIL-2酶切產(chǎn)物

圖3 豬白細(xì)胞介素2 PCR產(chǎn)物測(cè)序結(jié)果
2.3 重組乳酸桿菌的表達(dá) 將含有重組質(zhì)粒pSIP-409-pIL-2的乳酸桿菌誘導(dǎo)表達(dá)后,用12%SDS-PAGE凝膠電泳分析,與含有空載體的乳酸桿菌相比,重組的乳酸桿菌又到后5 h、7 h有pIL-2蛋白表達(dá),約在17.4 kDa處有明顯的條帶(圖4)。

圖4 重組乳酸桿菌誘導(dǎo)表達(dá)產(chǎn)物SDS-PAGE電泳
M:低分子量蛋白質(zhì)Marker; 1:L. pSIP-409; 2:L. pSIP-409-pIL-2誘導(dǎo)5 h后表達(dá)產(chǎn)物; 3:L. pSIP-409-pIL-2誘導(dǎo)7 h后表達(dá)產(chǎn)物
2.4 Western Blotting鑒定pIL-2蛋白的表達(dá) 如圖5所示,17.4 kDa處有一條明顯的蛋白印跡條帶,而含有空載體的對(duì)照組卻無(wú)印跡條帶。證實(shí)重組乳酸桿菌表達(dá)的pIL-2蛋白具有反應(yīng)原性。

圖5 Western Blotting檢測(cè)pIL-2蛋白
M:低分子量蛋白質(zhì)Marker; 1:L. pSIP-409-pIL-2; 3:L. pSIP-4092.5 pSIP409-IL-2重組乳酸菌對(duì)IPEC-J2豬腸道上皮細(xì)胞粘附性
通過(guò)與IPEC-J2豬腸道上皮細(xì)胞共培養(yǎng),豬白細(xì)胞介素-2重組乳酸菌與IPEC-J2豬腸道上皮細(xì)胞具有較好的粘附性(圖6);空載體乳酸菌組IPEC-J2豬腸道上皮細(xì)胞結(jié)構(gòu)松散,細(xì)胞間粘連蛋白表達(dá)量少,而豬白細(xì)胞介素-2重組乳酸菌組IPEC-J2豬腸道上皮細(xì)胞結(jié)構(gòu)緊密,細(xì)胞間粘連蛋白表達(dá)量較多。
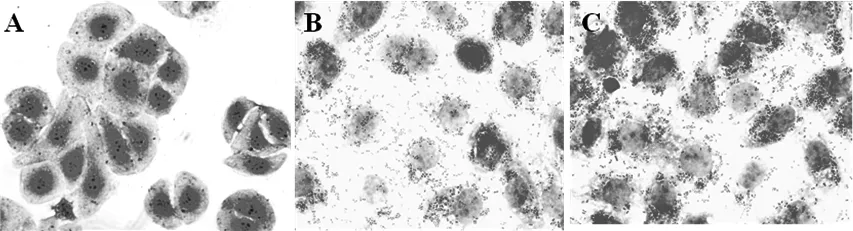
圖6 pSIP409-IL-2重組乳酸菌對(duì)IPEC-J2豬腸道上皮細(xì)胞粘附性 (×400)
A:PBS; B:pSIP-409; C:pSIP-409-pIL-2
3 討論
IL-2能夠誘導(dǎo)IL-2Rα和IL-2Rβ的表達(dá),而經(jīng)由IL-2Rβ 的異二聚體和γc胞質(zhì)區(qū)的IL-2信號(hào)可以導(dǎo)致至少3條信號(hào)通路(磷酸肌醇3-激酶、Ras-MAP 激酶和JAK-STAT 通路)的激活[10-11],同時(shí)也能夠激活STAT3 和STAT1。這3種不同方式的信號(hào)通路能夠調(diào)節(jié)細(xì)胞生長(zhǎng)、存活、誘導(dǎo)細(xì)胞凋亡和分化。IL-2促進(jìn)CD8+分化為效應(yīng)/記憶細(xì)胞毒性T淋巴細(xì)胞[12]。因此,IL-2的關(guān)鍵作用在于調(diào)節(jié)TH細(xì)胞的分化,發(fā)揮其生物學(xué)效應(yīng)。
在體外,Th 17的分化是通過(guò)IL-6和TGF-β共同介導(dǎo)的。抗IFNγ和抗IL-4被特有的添加阻止了Th 1和Th 2的分化,更有意義的是,IL-2信號(hào)能消除Th 17細(xì)胞的產(chǎn)生。因?yàn)镮L-6信號(hào)通過(guò)STAT3激活,而IL-2信號(hào)通過(guò)STAT5激活[13]。通過(guò)STAT5激活的IL-2在IL17a基因位點(diǎn)和STAT3結(jié)合位點(diǎn)相互競(jìng)爭(zhēng),抑制IL17a的轉(zhuǎn)錄。IL-2抑制IL-6Rα 和 gp130的表達(dá),然而,相反的是這些受體和IL17a的表達(dá)增加在IL2-/-T細(xì)胞中[14]。IL-2在Th17的調(diào)節(jié)中起著復(fù)雜作用。在添加CD4+細(xì)胞后,IL-2能促進(jìn)CD8+T細(xì)胞分化成效應(yīng)/記憶細(xì)胞毒性T淋巴細(xì)胞,這一過(guò)程依賴于IL-2/IL-2R信號(hào)的增強(qiáng), 說(shuō)明IL-2在記憶性CD8+T細(xì)胞擴(kuò)增過(guò)程中起著關(guān)鍵作用。增加IL-2/IL-2R信號(hào)能促進(jìn)效應(yīng)細(xì)胞毒性T淋巴細(xì)胞的分化[15]。
但同時(shí)也存在許多問(wèn)題如表達(dá)載體的選用:真核表達(dá)載體在真核細(xì)胞內(nèi)能穩(wěn)定地表達(dá)外源目的基因,但蛋白表達(dá)量較低、純化工藝復(fù)雜,從而提高規(guī)模化生產(chǎn)成本,而原核表達(dá)載體表達(dá)外源蛋白量高、成本低、可應(yīng)用性強(qiáng),但同時(shí)容易形成難溶的包涵體蛋白,純化后所表達(dá)蛋白構(gòu)象是否具有較高生物活性[16];外源性與內(nèi)源性差異:外源IL-2畢竟不是自身所產(chǎn)生,在調(diào)節(jié)機(jī)體免疫系統(tǒng)的同時(shí)存在潛在的副作用;給藥途徑:采用注射方法和口服建立黏膜免疫是兩種不同的途徑,針對(duì)不同的疾病究竟哪種方法的免疫和治療效果最佳還有待摸索;添加劑量與時(shí)相:大量試驗(yàn)表明,低劑量使用IL-2效果不佳,而高劑量使用又會(huì)產(chǎn)生自身免疫抑制,導(dǎo)致自身免疫系統(tǒng)失衡,IL-2半衰期短,有時(shí)還可在體內(nèi)誘導(dǎo)產(chǎn)生一定抗體,這也是研制長(zhǎng)效劑型IL-2產(chǎn)品制約因素之一;以及抗原表位的優(yōu)化和動(dòng)物安全性等方面還有待于進(jìn)一步研究。
