奶牛隱性乳房炎致病性大腸桿菌AI-2信號分子檢測
2018-12-05 08:59:40曹素芳孔蘭芳皇甫和平陳文定
中國獸醫(yī)雜志 2018年8期
關(guān)鍵詞:檢測
曹素芳,吳 迪,孔蘭芳,皇甫和平,陳文定
(1.河南牧業(yè)經(jīng)濟(jì)學(xué)院動物醫(yī)學(xué)院,河南 鄭州 450046;2.河南農(nóng)業(yè)大學(xué),河南 鄭州 450002)
奶牛乳房炎是指奶牛乳腺組織受到微生物感染、機(jī)械損傷、熱損傷和化學(xué)物品刺激而引起的一種炎癥。在乳房炎中隱性乳引起產(chǎn)奶量急劇下降,奶品質(zhì)下降。致病性大腸桿菌是導(dǎo)致奶牛隱性乳房炎最主要的乳房炎最為多見,發(fā)病率達(dá)到60%以上,是主要致病菌之一。
研究發(fā)現(xiàn),為了適應(yīng)復(fù)雜的生活環(huán)境,細(xì)菌通過產(chǎn)生的信號分子(Autoinducer,AI)進(jìn)行細(xì)胞間或細(xì)胞自身內(nèi)部的信息交流,統(tǒng)一協(xié)調(diào)細(xì)菌間的群體行動[1],共同完成個體無法完成的生物學(xué)功能。細(xì)菌群體間信號交流現(xiàn)象稱為細(xì)菌密度感應(yīng)現(xiàn)象。研究顯示,革蘭陰性菌中的LuxS/AI-2密度感應(yīng)系統(tǒng),可通過合成AI-2分子調(diào)節(jié)細(xì)菌多種生物學(xué)功能[2-3]。為了探索牛源大腸桿菌AI-2信號分子產(chǎn)生情況,本試驗以分離于河南省某奶牛場隱性乳房炎中致病性大腸桿菌為研究對象,利用哈維弧菌BB170檢測致病性大腸桿菌AI-2產(chǎn)生情況,探索不同培養(yǎng)條件及不同生長時期大腸桿菌分泌AI-2分子活性,應(yīng)用實時熒光定量PCR研究其催化基因luxS和pfs的轉(zhuǎn)錄水平,為進(jìn)一步探討牛源致病性大腸桿菌的致病機(jī)理奠定基礎(chǔ)。
1 材料與方法
1.1 菌種 大腸桿菌E80814、W41、EA701205、E30707和E285株分離于河南新鄉(xiāng)某奶牛場;哈維弧菌BB170和BB152均由韓先干博士惠贈;DH5α本室保存。
1.2 主要試劑 L-Arg、Marine Broth、Vitamin-free Casa -mino Acids,均購自Difco公司;TRIZol,購自Promega公司。
1.3 方法
1.3.1 奶牛隱性乳房炎中大腸桿菌信號分子AI-2的制備 參照文獻(xiàn)[4]方法,稍加改進(jìn)。具體如下:挑取陽性對照菌BB152單個菌落加入5 mL的MB液體培養(yǎng)基中,28 ℃振蕩16~18 h,將過夜菌液稀釋100倍后接入5 mL MB中,28 ℃振蕩培養(yǎng)過夜,13 000 r/min(4 ℃)離心10 min,取上清,過濾除菌,-80 ℃保存?zhèn)溆谩?/p>
挑取奶牛隱性乳房炎致病性大腸桿菌E80814、W41、EA701205、E30707、E285株及陰性對照菌DH5α單個菌落,分別接種5 mL LB液體培養(yǎng)基中,37 ℃振蕩培養(yǎng)16~18 h,培養(yǎng)的菌液再稀釋100倍后分別加入5 mL LB液體培養(yǎng)基中,37 ℃培養(yǎng)過夜,13 000 r/min(4 ℃)離心10 min,取上清,過濾除菌,-80 ℃保存?zhèn)溆肹5]。
1.3.2 哈維弧菌BB170的培養(yǎng) 挑取哈維弧菌BB170單個菌落加入5 mL MB液體中,28 ℃振蕩培養(yǎng)過夜,過夜菌液稀釋100倍后加入5 mL MB液體中,28 ℃振蕩培養(yǎng)過夜,使其OD620值達(dá)到0.4~0.5。
1.3.3 檢測AI-2的活性 參照文獻(xiàn)[5]方法,稍加改進(jìn)。具體如下:將上述BB170菌液用新鮮配制的AB培養(yǎng)基進(jìn)行5 000倍稀釋,分別加入10%E80814、W41、EA701205、E30707和E285培養(yǎng)上清,陽性菌BB152培養(yǎng)上清,陰性菌DH5α培養(yǎng)上清,28 ℃ 振蕩培養(yǎng)5 h后用全自動多功能酶標(biāo)儀檢測BB170的發(fā)光情況,每組培養(yǎng)物做3個重復(fù)孔。
1.3.4 檢測奶牛隱性乳房炎的大腸桿菌不同生長時期AI-2的表達(dá) 取E285單個菌落接種于5 mL的LB液體培養(yǎng)基中,37 ℃振蕩培養(yǎng)過夜,過夜菌液進(jìn)行100倍稀釋后加入LB液體培養(yǎng)基中,37 ℃培養(yǎng)至遲緩期(OD600值=0.1),對數(shù)前期(OD600值=0.7),對數(shù)中期(OD600值=1.6),對數(shù)后期(OD600值=2.3),穩(wěn)定期(OD600值=2.6),按照1.2.1的方法制備培養(yǎng)上清,-80 ℃冷凍備用。AI-2檢測同1.2.3,沉淀用于提取細(xì)菌總RNA。
1.3.5 不同培養(yǎng)條件奶牛隱性乳房炎大腸桿菌的AI-2的活性檢測 取E285單個菌落接種于5 mL的LB液體培養(yǎng)基中,37 ℃培養(yǎng)過夜,按1∶100稀釋后分別加入含0.5%葡萄糖、0.5%甘露醇、0.5%乳糖、0.5%蔗糖、0.5%麥芽糖及0.5%NaCl的LB液體培養(yǎng)基中,37 ℃培養(yǎng)至對數(shù)期(OD600值=1.2),按照1.2.1的方法收集AI-2,-80 ℃ 冷凍備用,AI-2的活性檢測同1.2.3。沉淀用于提取細(xì)菌總RNA。
1.3.6pfs和LuxSmRNA的水平檢測
1.3.6.1 引物設(shè)計 根據(jù)GenBank 中大腸桿菌pfs和luxS基因序列分別設(shè)計特異性引物,pfs-F(5′-ACGCCATTACCGTTAAGATG-3′)和pfs-R(5′-AGTGATGCCAGAAAGAGGGA-3′),擴(kuò)增長度為169 bp;LuxS-F(5′-CGGCAACAGCCAGGAACTCA-3′)和LuxS-R(5′-GCGAAAATCCGCCACAACTT-3′),擴(kuò)增長度為81 bp,同時設(shè)計內(nèi)參引物dnaE-F(5′-CTGGCA TTGTCATGGACTCT-3′)和dnaE-R(5′-GCGATGATCTT GATCT TCAT-3′),擴(kuò)增長度約為80 bp。
1.3.6.2 細(xì)菌總RNA的提取 取適量的1.2.4和1.2.5中的細(xì)菌沉淀,加入1 mL TRIZol,混合均勻,再加入200 μL氯仿,振蕩混勻,20 ℃~25 ℃靜置3 min;13 000 r/min(4 ℃)離心15 min,吸取上清,加入500 μL 異丙醇,振蕩均勻,13 000 r/min(4 ℃)離心10 min,去上清,用75%乙醇洗滌,真空干燥,加入30 μL無RNase的ddH2O溶解,-80 ℃保存?zhèn)溆谩?/p>
1.3.6.3 反轉(zhuǎn)錄合成cDNA 取16 μL提取的細(xì)菌總RNA,加入1 μL的隨機(jī)引物,70 ℃溫育5 min,冰浴2 min,再依次加5 μL 5×M-MLV Buffer, 2 μL 4dNTP,1 μL M-MLV酶,1 μL RNase抑制劑,混勻,42 ℃溫育1 h,-80 ℃保存?zhèn)溆谩?/p>
1.3.6.4 Real-time PCR法檢測mRNA轉(zhuǎn)錄水平 以dnaE為內(nèi)參,應(yīng)用real-time PCR檢測luxS和pfs基因mRNA轉(zhuǎn)錄情況。將獲得的cDNA 進(jìn)行5倍稀釋,制定標(biāo)準(zhǔn)曲線,每個稀釋度重復(fù)兩孔,待測樣品做重復(fù)3次。
在20 μL體系中加入10 μL 的Real- time PCR Master Mix,1 μL的cDNA,0.5 μL的上游引物,0.5 μL的下游引物,8 μL 的ddH2O,混勻后50 ℃ 溫育2 min后進(jìn)行擴(kuò)增,95 ℃ 變性10 min后進(jìn)行循環(huán),95 ℃ 15 s,60 ℃ 1 min,40cycle。應(yīng)用2-△△Ct(Livak)法計算luxS基因和pfs基因的mRNA。
2 結(jié)果
2.1 不同的奶牛隱性乳房炎大腸桿菌AI-2的檢測 以BB152為陽性對照,DH5α為陰性對照,利用哈維弧菌BB170檢測分離于奶牛隱性乳房炎的致病性大腸桿菌AI-2的產(chǎn)生情況,結(jié)果表明,分離于奶牛隱性乳房炎致病性大腸桿菌E285、E80814、W41、E30707和EA701205株均有AI-2信號分子產(chǎn)生,但產(chǎn)生的AI-2的量均低于陽性對照(圖1-1)。
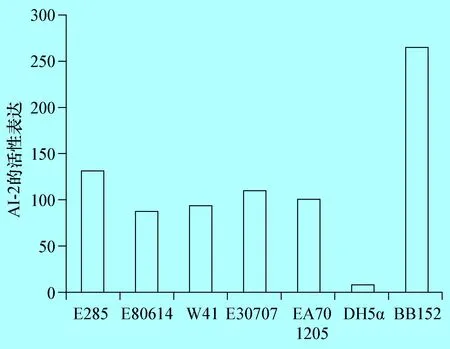
圖1不同的牛致病性大腸桿菌AI-2的檢測
2.2 不同生長時期E285株AI-2檢測 對不同生長時期的牛致病性大腸桿菌E285株AI-2活性檢測結(jié)果表明,在遲緩期、對數(shù)前期E285的基本沒有AI-2的表達(dá),從對數(shù)中期開始E285細(xì)菌開始大量表達(dá)AI-2,到對數(shù)后期達(dá)到高峰,為陰性對照的12倍,隨后下降,到穩(wěn)定期已下降為陰性對照的3.6倍(圖2)。
2.3 檢測不同培養(yǎng)條件下奶牛隱性乳房炎大腸桿菌AI-2產(chǎn)生 將致病性大腸桿菌E285株分別接種于含0.5%蔗糖、0.5%麥芽糖、0.5%葡萄糖、0.5%甘露醇、0.5%NaCl及0.5%乳糖液體LB培養(yǎng)基中,檢測其AI-2活性,結(jié)果表明,LB液體中加入0.5%蔗糖、0.5%麥芽糖、0.5%葡萄糖、0.5%甘露醇、0.5%NaCl及0.5%乳糖時,AI-2的活性明顯提高(P<0.01),均高于陰、陽性對照。分別為陰性對照的3.8倍、3.4倍、4.6倍、3.7倍、3.5倍、5.3倍(圖3),其中加入乳糖后E285株的AI-2上調(diào)倍數(shù)最大。
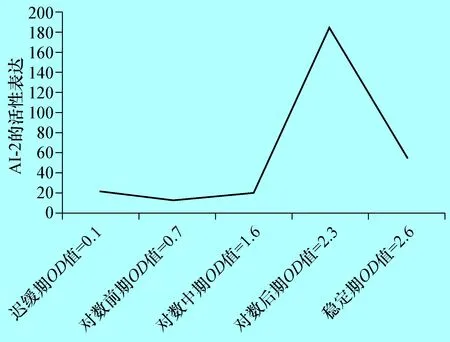
圖2E285株在不同生長時期AI-2活性檢測
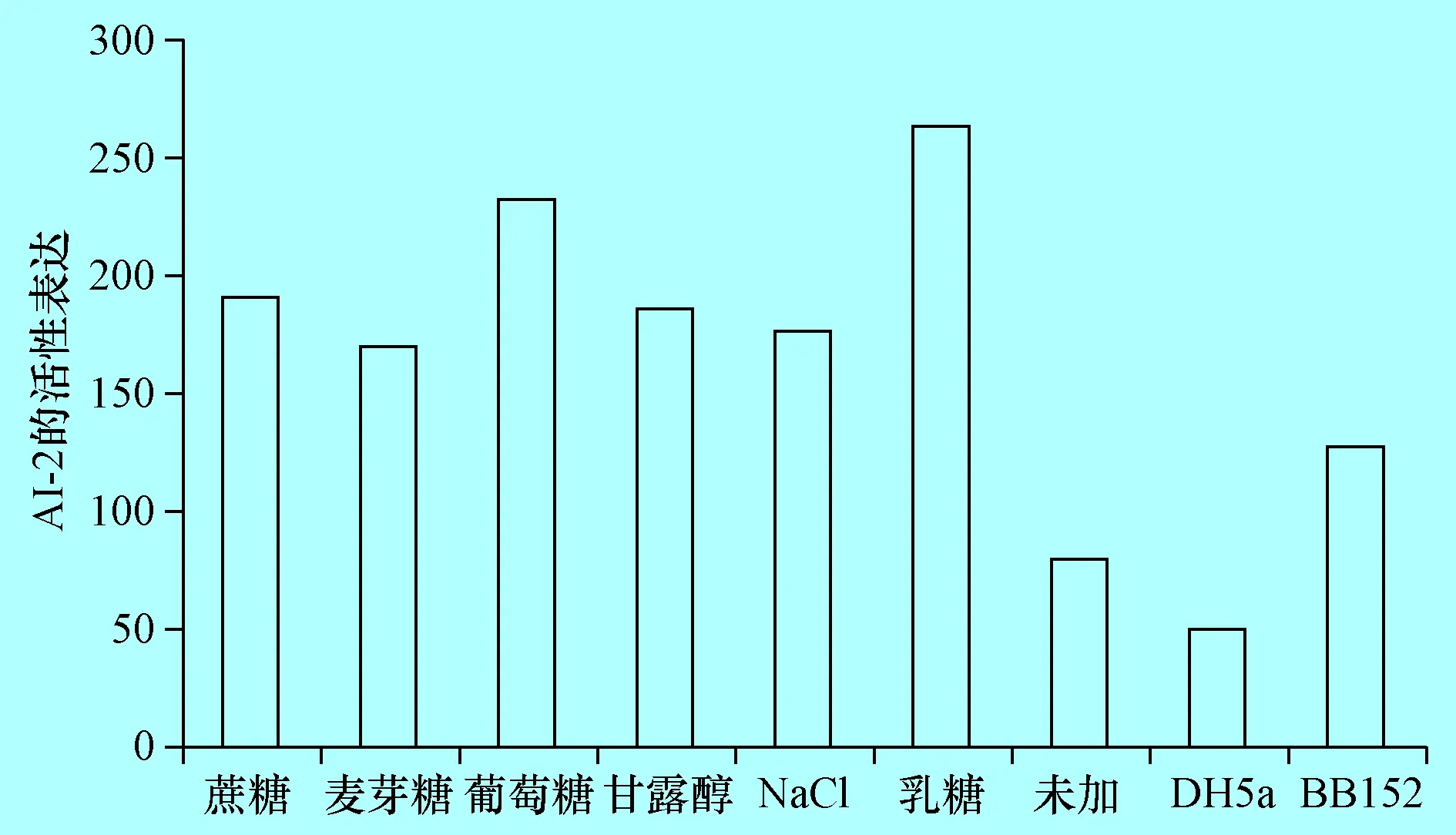
圖3E285不同培養(yǎng)條件下AI-2活性檢測(P<0.01)
2.4 奶牛隱性乳房炎大腸桿菌E285株不同生長時期luxS與pfsmRNA的檢測 根據(jù)real-time PCR擴(kuò)增曲線顯示dnaE基因、LuxS基因和pfs基因擴(kuò)增效率均到達(dá)96%-104%(圖略),應(yīng)用2-△△Ct(Livak)法分析luxS基因和pfs基因的mRNA水平,結(jié)果顯示,從遲緩期開始E285株luxS的mRNA水平顯著增加(P<0.01),到對數(shù)前期達(dá)到最高,為遲緩期的8.7倍,隨后就慢慢降低,到穩(wěn)定期時mRNA水平降至陰性對照的3.6倍(圖4),這與E285株在不同生長期AI-2的表達(dá)變化結(jié)果一致。pfs基因的mRNA的轉(zhuǎn)錄結(jié)果分析表明,其轉(zhuǎn)錄水平在前期并未有大的變化,只有在穩(wěn)定期達(dá)到最高,是遲緩期的5.3倍。
2.5 E285株不同培養(yǎng)條件下luxS基因和pfs基因mRNA的分析 當(dāng)LB液體中分別加入0.5%蔗糖、麥芽糖、葡萄糖、甘露醇、NaCl和乳糖時,E285株luxS基因的mRNA均明顯上調(diào)(P<0.01),分別為未加的2.39倍、2.12倍、2.89倍,2.33、2.21倍和3.31倍,加入乳糖luxS基因的mRNA量最高,與AI-2活性表達(dá)一致。對pfs基因的mRNA分析結(jié)果表明,加入蔗糖、葡萄糖后明顯下降(P<0.01),分別為未加時的0.82倍和0.9倍,而加入麥芽糖、甘露醇、NaCl和乳糖時上調(diào)明顯,分別為未加時的2.1倍、1.96倍、2.03倍、3.21倍(圖5),與AI-2活性表達(dá)變化無相關(guān)性。
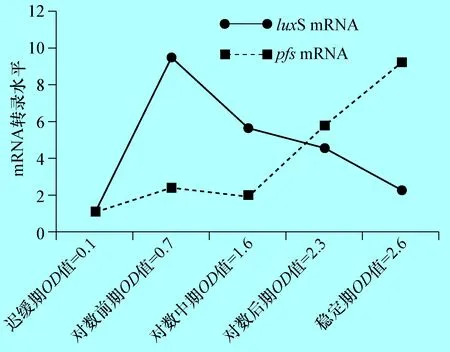
圖4 E285株在不同生長時期luxS基因和pfs基因的mRNA轉(zhuǎn)錄 (P<0.01)
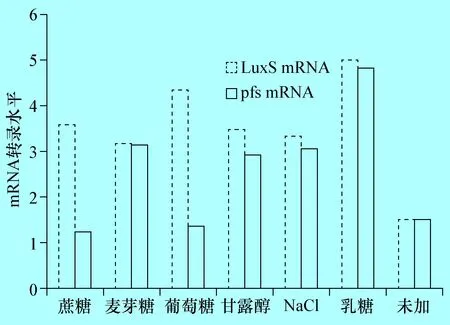
圖5 E285株在不同培養(yǎng)條件下luxS基因和pfs基因的mRNA轉(zhuǎn)錄
3 討論
研究證明,革蘭陰性菌存在luxS/AI-2密度感應(yīng)系統(tǒng),該系統(tǒng)調(diào)控著細(xì)菌群體活動,與細(xì)菌的致病性、毒力基因的表達(dá)和生物發(fā)光等有關(guān)。徐蘋[6]等研究弗氏志賀菌的信號分子AI-2時發(fā)現(xiàn),弗氏志賀菌在生長過程中分泌的AI-2具有生物學(xué)活性,通過獲得基因重組AI-2菌株構(gòu)建發(fā)現(xiàn),密度感應(yīng)系統(tǒng),重組菌株具有更好的生長優(yōu)勢。羅哲[7]等通過構(gòu)建luxS基因缺失株的傷寒沙門菌,與野生型菌相比,缺失株不能使報告菌株發(fā)光,證明luxS基因可以介導(dǎo)AI-2的合成,并引起細(xì)菌的發(fā)光。本試驗以DH5α為陰性對照,BB152為陽性對照,檢測了分離于奶牛隱性乳房炎致病性大腸桿菌中AI-2產(chǎn)生情況,結(jié)果表明,所獲得的5株均可以產(chǎn)生AI-2,其中E285株AI-2活性最高,但均低于陽性對照,不同的菌株間產(chǎn)生AI-2的水平差異很大,究其原因可能是由于菌株之間的差異導(dǎo)致而成的。
本試驗中對E285株不同生長時期中AI-2信號分子檢測結(jié)果表明,在對數(shù)后期AI-2表達(dá)量最高,與Jones M B等[8]、羅哲等[8]報道的傷寒沙門菌的AI-2信號分子的表達(dá)量最高是在對數(shù)中期的結(jié)果不一致,其原因可能是菌種不同。本試驗結(jié)果顯示,E285株的AI-2信號分子的變化規(guī)律與luxS表達(dá)量呈正相關(guān),而與pfs的表達(dá)量沒有明顯的相關(guān)性。Moslehi-Jenabian S[9]等報道了嗜酸乳桿菌中AI-2信號分子的活性與luxS的表達(dá)量之間的關(guān)系,結(jié)果與本研究相一致。Han等[10]研究證明,細(xì)菌中的S-核糖同型半胱氨酸對細(xì)菌中的luxS表達(dá)量有一定的影響。對數(shù)生長期前,S-核糖同型半胱氨酸濃度較低,luxS表達(dá)量是AI-2分子數(shù)量的直接決定因素。細(xì)菌對數(shù)生長期后,S-核糖同型半胱氨酸濃度大幅提高,導(dǎo)致pfs的表達(dá)量也顯著上升,而pfs的表達(dá)有助于S-核糖同型半胱氨酸的生物降解。
在培養(yǎng)E285株時加入麥芽糖、葡萄糖、甘露醇、乳糖及NaCl后,AI-2 信號分子的活性顯著提升,其中加入乳糖誘導(dǎo)產(chǎn)生的AI-2最多,活性最強(qiáng),與豬鏈球菌、炭疽桿菌、鼠傷寒沙門菌等結(jié)果相符,推測其原因可能是麥芽糖、葡萄糖、乳糖等濃度提高導(dǎo)致糖酵解途徑的活躍,甘油-3-磷酸脫氫酶得到了累積,從而抑制了基因的表達(dá)量,lsr 轉(zhuǎn)運(yùn)蛋白可以將AI-2信號分子從胞外運(yùn)輸?shù)桨麅?nèi),其表達(dá)量的降低使得AI-2信號分子在胞外大量累積。但也有報道稱,麥芽糖和葡萄糖提供了充足的碳源,AI-2信號分子被大量的運(yùn)輸?shù)桨馑耓11]。本試驗中加乳糖時沒有導(dǎo)致AI-2下降,與白灝[6]報道的禽致病性大腸桿菌結(jié)果不同,推測可能是牛源致病性大腸桿菌對蔗糖的代謝途徑的不同。
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
