色季拉山急尖長苞冷杉天然更新影響因素
2018-11-30 06:12:26王瑞紅李江榮
浙江農林大學學報 2018年6期
王瑞紅, 李江榮, 潘 剛
(1.西藏農牧學院 高原生態研究所,西藏 林芝860000;2.西藏農牧學院 西藏林芝高山森林生態系統國家野外科學觀測研究站,西藏 林芝860000;3.西藏農牧學院西藏高原森林生態教育部重點實驗室,西藏 林芝860000;4.西藏農牧學院 資源與環境學院,西藏 林芝860000)
急尖長苞冷杉Abies georgeivar.smithii屬于耐蔭性植物,特別是在幼苗期,溫涼和寒冷的氣候區域比較適宜其生長。常在高海拔地區至低海拔的亞高山與高山地帶的谷地、陰坡及半陰坡處形成純林,與喜冷濕的云杉Picea aspereta,落葉松Larix gmelinii,鐵杉Tsuga chinensis,松樹Pinus及闊葉樹形成針葉或針闊混交林。急尖長苞冷杉是西藏東南部陰坡高山林線森林群落的主要建群樹種之一,在色季拉山,主要分布在海拔3 300~4 300 m,分布范圍比較廣泛[1]。但是由于長期過度采伐,林地生態環境惡化,天然更新比較困難,其自然分布區急劇縮小,種群數量逐漸減少[2]。在實地調查中,西藏色季拉山急尖長苞冷杉幼苗很豐富,但是幼苗向幼樹過渡期間死亡率比較高,導致其天然更新效果不理想。目前,國內對于冷杉天然更新影響因素方面的研究比較多,主要集中在不同采伐方式[3-5]、生態學特性[6-7]、海拔與坡向[8-9]、土壤種子庫[10-11]及地被物[9,12-15]等。對于西藏高寒環境下急尖長苞冷杉天然更新影響因素的研究還比較少,主要為林隙對其天然更新的影響[16-17]。本研究通過海拔、坡向、種子大小、林隙等方面展開色季拉山急尖長苞冷杉天然更新影響因素的初步研究,為進一步認識色季拉山原始暗針葉林的天然更新,特別是如何采取措施促進其更新具有重要的實踐意義。
1 研究區概況
研究區位于西藏魯朗境內藏東南的林芝地區。色季拉山山體海拔為2 100~5 300 m,水平走向大致呈西北向東南,由此形成了陽坡與陰坡。由于海拔差異較大,林下發育的土壤也出現明顯的不同。從低海拔到高海拔呈現的土壤類型依次為山地棕壤、山地酸性棕壤、山地漂灰土、亞高山草甸土、高山草甸土、高山寒漠土[18]。研究區域屬典型的亞高山寒溫帶半濕潤氣候,雅魯藏布江為其提供水汽通道,降雨比較豐沛,但干濕季節比較分明。西藏林芝高山森林生態定位研究站數據顯示:降水多集中在每年4-10月,年降水量為875~1 350 mm,年平均氣溫為-0.73℃,7月為最暖月,1月氣溫最低。該區域喬木以原始暗針葉林急尖長苞冷杉為主,還有少量的林芝云杉Picea likiangensisvar.linzhiensis和方枝柏Sabina saltuaria;林下灌木生長繁茂,主要有西南花楸Sorbus rehderiana,紅景天Rhodiola rosea,杯萼忍冬Lonicera lanceolata,懸鉤子Rubus corchorifolius,蛾眉薔薇Omeiensis,林芝杜鵑Rhododendron tanastylumvar.lingzhiense等;草本主要有龍膽Gentiana scabra,委陵菜Potentilla chinensis,西南草莓Fragaria moupinensis,卷葉黃精Polygonatum cirrhifolium,野丁香Syringa persica等;地被層主要包括枯枝落葉層和苔蘚層,其中苔蘚層比較發達,尤其是陰坡,厚度達10 cm以上,蓋度約90%。
2 研究方法
2.1 樣地設置
采用梯度格局法設置樣地,在陰坡與陽坡均在海拔3 700~4 200 m處,每升高100 m設置1塊樣地,共設置樣地12塊,樣地面積均為30 m×30 m。采用鄰接格子法把樣地劃分成9個10 m×10 m的樣方,各樣方設置5個2 m×2 m的小樣方進行灌木調查,并在2 m×2 m的樣方中設置5個1 m×1 m的小樣方調查草本、苔蘚植物、凋落物厚度及幼苗數量。在樣地中或附近的林下、林緣、林隙處分別設置3個1 m×1 m的小樣方,記錄各小樣方內幼苗數量,測量樣方內各幼苗高度、周邊苔蘚層厚度及凋落物厚度。
2.2 測量方法
2.2.1 球果采集 對海拔3 700~4 200 m樣地中急尖長苞冷杉每木檢尺,各海拔均選取標準木,待種子成熟初期,將樹冠分上層、中層、下層共3層,每層用高枝剪取下5個球果,標記裝袋帶回實驗室。
2.2.2 種子特征及質量測量 從球果中手剝出種子,待種子自然風干后,隨機選取各海拔陰坡與陽坡各50粒種子,用游標卡尺測量種子帶翅長度、去翅長度、種子寬度、種子厚度,用100粒法測量其千粒重。
2.2.3 苔蘚、凋落物厚度及幼苗數量的測量 在海拔3 700~4 200m處設置的5個1 m×1 m小樣方中記錄幼苗數量,四周分別測量苔蘚植物厚度及凋落物厚度,另在林下、林緣、林隙處設置的小樣方中記錄每個小樣方內幼苗的數量,用直尺測量樣方四周苔蘚層厚度及凋落物厚度。
2.3 數據分析
數據統計分析使用SPSS 19.0,利用Excel 2010制作圖表。
3 結果與分析
3.1 海拔與坡向對急尖長苞冷杉天然更新的影響
3.1.1 不同海拔與坡向種子特征差異性分析 海拔與坡向作為重要的環境因子,其對溫度、濕度、光照等諸多環境因子產生直接的影響。這些因子對急尖長苞冷杉種子去翅長度、帶翅長度、寬度及厚度均產生影響。對色季拉山種子特征研究表明:在陰坡,種子帶翅長度、種子去翅長度、種子寬度、種子厚度在海拔3 800或3 900 m處達到最大值,最大值分別為16.87,10.54,3.85,2.31 mm,最小值分別為8.03,4.27,1.69,0.85 mm;在陽坡,種子帶翅長度、種子去翅長度、種子寬度、種子厚度亦是在海拔3 800或3 900 m達最大,最大值分別為 17.44,11.52,4.18,2.93 mm,最小值分別為8.79,4.85,1.81,0.72 mm;陽坡種子帶翅長度、種子去翅長度、種子寬度、種子厚度要略好于陰坡。由表1可以得出:在陰坡,急尖長苞冷杉種子去翅長度、寬度、厚度均表現為差異不顯著,但是種子帶翅長度表現為差異顯著(P<0.05,F=2.467);在陽坡,種子去翅長度、種子寬度、種子厚度表現為差異性不顯著(P>0.05),而種子帶翅長度表現為差異顯著(P<0.05,F=2.527)。由表2可以得出:種子特征均表現為差異不顯著。可見,色季拉山急尖長苞冷杉種子形狀相對穩定,同一坡向不同海拔對種子形狀影響較小,同一海拔不同坡向對種子形狀影響不明顯。

表1 不同海拔種子特征差異性分析Table 1 Difference analysis of seeds characteristic on different elevations
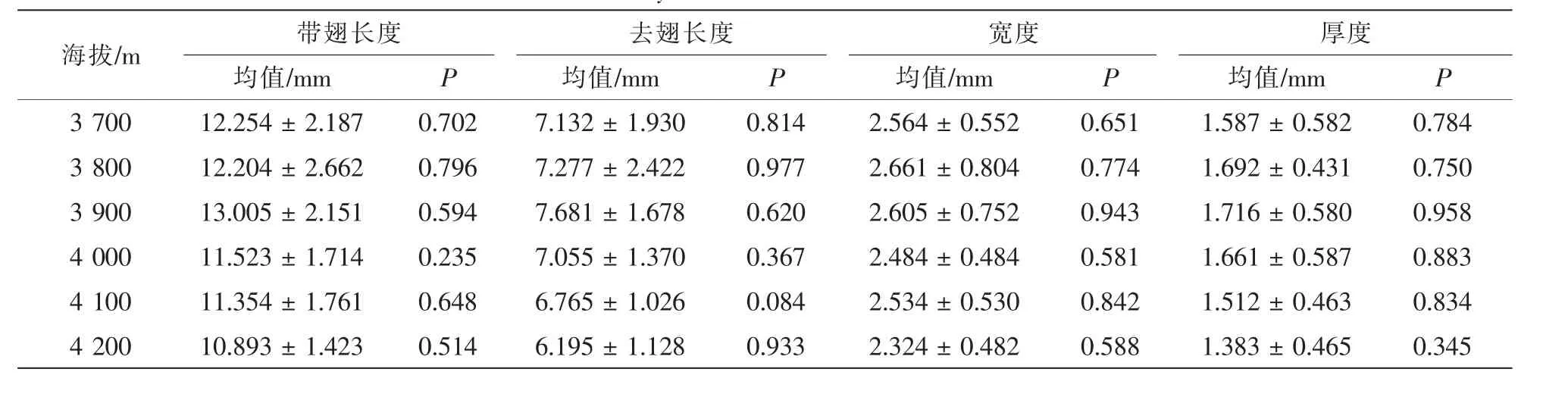
表2 不同坡向種子特征差異性分析Table 2 Difference analysis of seeds characteristic on different slopes
3.1.2 種子質量對天然更新的影響 種子是植物生活史中的一個重要環節,關系著種群未來的命運。種子千粒重在一定程度上可以預知種子的飽滿度,同時也是衡量種子品質的重要指標,對種子萌發、休眠、出土、幼苗發育及幼苗生長產生影響,從而影響急尖長苞冷杉林的天然更新。急尖長苞冷杉種子帶翅且比較小,主要靠重力擴散及借助風力,擴散能力比較強,擴散距離比較遠,可以為急尖長苞冷杉提供更多的生長空間。武高林等[19]研究表明:較小的種子擴散能力強,有利于種群建植和天然更新,能減少動物捕食的數量,可形成持久土壤種子庫。在色季拉山,陰坡種子千粒重分布范圍為3.32~12.86 g,海拔3 900 m處種子千粒重達到最優值,平均為10.46 g;陽坡種子千粒重分布范圍為3.72~14.68 g,在海拔3 800 m處最佳,平均為12.37 g;種子千粒重在陰坡與陽坡隨著海拔的升高均呈先升高后下降的趨勢;總體來看,陽坡種子質量要優于陰坡。由表3可以得出:陰坡千粒重(P=0.008,F=3.500)和陽坡千粒重(P=0.003,F=4.217)均表現為差異顯著;由表4可以得出: 在海拔 3 800 m(P=0.038,F=5.043)和3 900 m(P=0.044,F=4.671)處種子千粒重表現為差異顯著;3 700,4 000,4 100,4 200 m處均表現為差異不顯著。

表3 不同坡向種子質量差異性分析Table 3 Difference analysis of seeds weight on different slopes

表4 不同海拔種子質量差異性分析Table 4 Difference analysis of seeds weight on different elevations
3.1.3 不同海拔與坡向天然更新幼苗數量 由圖1可以得出:隨著海拔的升高急尖長苞冷杉幼苗數量總體上呈現先增加后減少的趨勢,在海拔3 900 m處達到最大值,陰坡為15.2株·m-2,陽坡為14.2株·m-2;在海拔4 000 m處,由于調查樣地處于一亂石灘,水土流失嚴重,保水能力較差,因此,此處幼苗數量急劇下降;高海拔處幼苗數量遞減,主要是由于海拔升高氣溫會逐漸降低,過低的溫度致使土壤溫度常處于凍結狀態,幼苗根系很難從土壤中吸收水分,其根系的生長受到抑制,再加上光照強度增大,紫外線增強,水分散失比較多,易造成幼苗生理性干旱而死亡;在研究海拔范圍內陰坡單位面積內幼苗數量均高于陽坡,主要原因為陽坡光照強烈、溫度較高、土壤含水量較低,對耐蔭性樹種急尖長苞冷杉幼苗的發育不利。
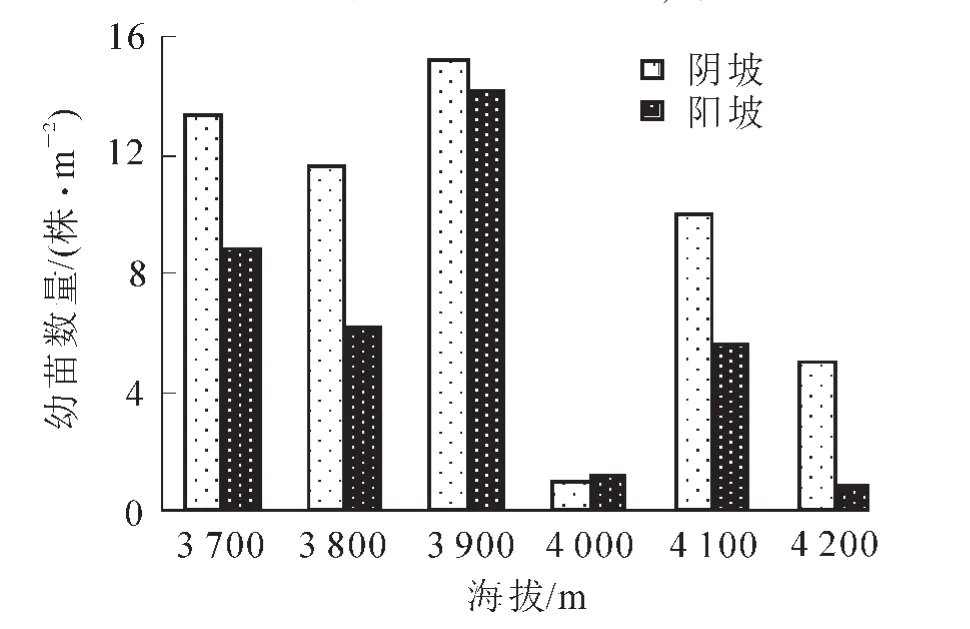
圖1 單位面積內不同海拔不同坡向幼苗數量Figure 1 Number of seedlings in per unit area on different elevations and slopes
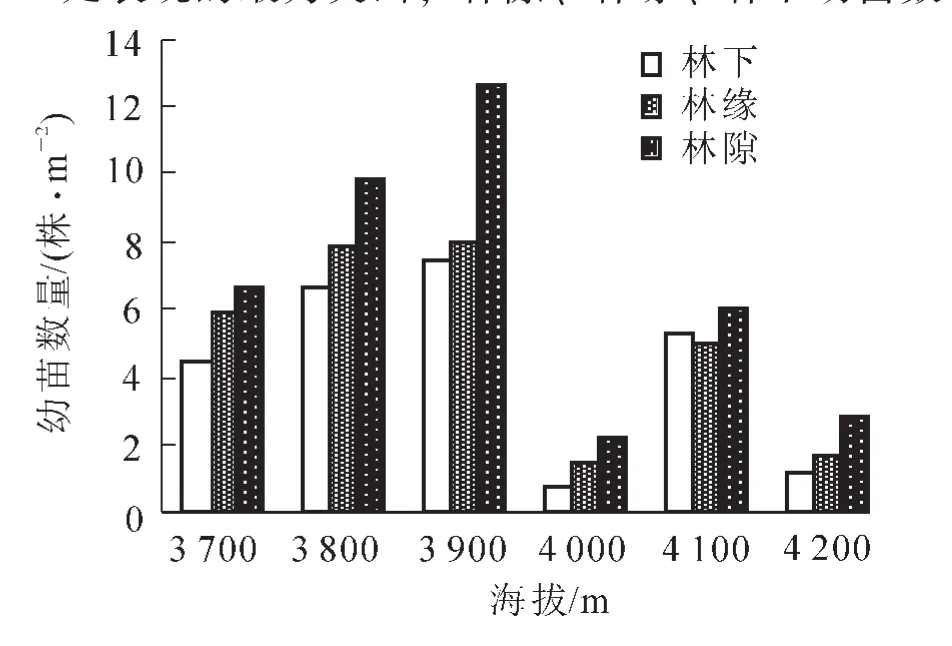
圖2 海拔3 700~4 200 m不同位置幼苗數量Figure 2 Number of seedlings at different positions of 3 700~4 200 m
3.2 林隙對天然更新的影響
林隙是指森林群落中1株以上林冠層樹木死亡形成的將會由新個體占有與更新的空間[20]。羅大慶等[17]認為在亞高山急尖長苞冷杉林中,干基折斷是形成林隙的主要方式,其次為干中折斷與拔根風倒,枯立的較少,且風是形成林隙的主要外力。在色季拉山,急尖長苞冷杉林中林隙廣泛分布且面積較大,四周可達10 m,是其天然更新的起點和自然演替的重要機制之一。由圖2得出:不同海拔單位面積幼苗數量均表現為林隙>林緣>林下,其中在海拔3 900 m處表現的最為突出,林隙、林緣、林下幼苗數量分別為12.7,8.0,7.5株·m-2;在林隙、林緣及林下均表現為隨著海拔的升高急尖長苞冷杉幼苗數量呈現先增加后減少的趨勢。
3.3 苔蘚、凋落物層對天然更新的影響
苔蘚層、凋落物層位于森林植被層與土壤層之間,是森林生態系統的重要組成部分,同時也是林地水汽交換的重要界面[21-24]。由表5可以得出:苔蘚層厚度、凋落物厚度均大于死亡幼苗根長,因此導致較多的幼苗死亡,達3.000株·m-2;不同海拔死亡幼苗根長差異性不顯著,苔蘚厚度、幼苗數量差異性顯著,凋落物厚度、死亡幼苗數量差異性極顯著。主要有以下原因:苔蘚與凋落物具有遮光,減少光照的作用,有利于急尖長苞冷杉種子萌發、幼苗生長;苔蘚、凋落物覆蓋林地,可以減少動物對種子的覓食,且凋落物層的分解會增加林地養分,促進急尖長苞冷杉幼苗的生長;另外,苔蘚層、凋落物層具有明顯的持水功能,特別是在干旱脅迫條件下,苔蘚層、凋落物層的水分可以暫時補給給幼苗;但是由于苔蘚與凋落物的機械阻礙作用,使萌發種子根系在較短的時間很難扎進土壤,隨著急尖長苞冷杉幼苗的生長,對水分、光照及其他營養元素的需求增多,使冷杉幼苗缺失水分及其他營養元素而死亡。

表5 苔蘚厚度、凋落物厚度、死亡幼苗根長、死亡幼苗數量差異性分析Table 5 Difference analysis of thickness of moss layer,thickness of litter,length of death seeding and number of death seeding
3.4 相關性分析
從相關性分析中可以得出(表6):幼苗數量與林分郁閉度、帶翅長度、千粒重、凋落物厚度均呈現顯著正相關,與苔蘚厚度呈現極顯著正相關。這說明色季拉山急尖長苞冷杉林分郁閉度、帶翅長度、千粒重、苔蘚厚度、凋落物厚度可能有利于急尖長苞冷杉天然更新,苔蘚厚度對其天然更新的影響最大。
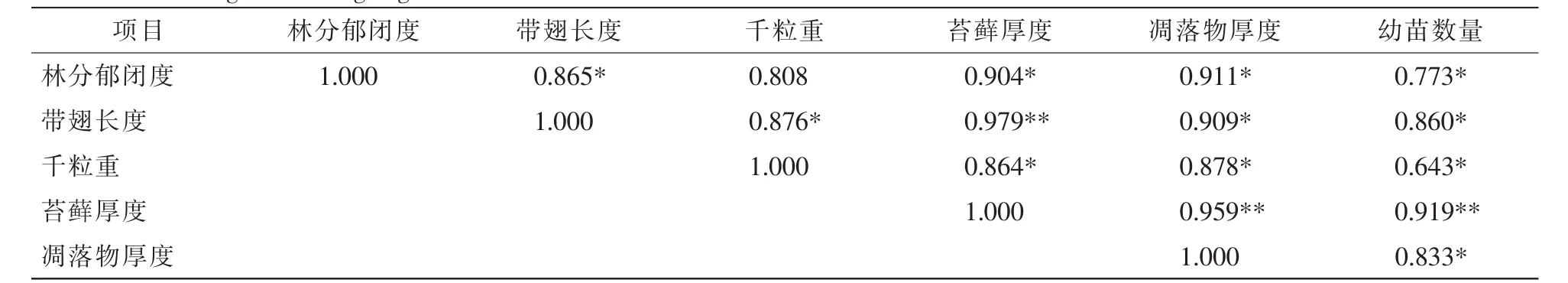
表6 急尖長苞冷杉天然更新影響因子與幼苗數量之間的相關系數Table 6 Correlation coefficients among factors influencing natural regeneration between each influencing factor and the number of seedlings of Abies georgei var.smithii
4 討論與結論
由于海拔與坡向的不同,急尖長苞冷杉種子帶翅長度、種子去翅長度、種子寬度、種子厚度均存在差異性,尤其是陰坡與陽坡種子帶翅長度均表現為顯著差異。色季拉山急尖長苞冷杉種子形狀相對穩定,同一坡向不同海拔對種子形狀影響較小,同一海拔不同坡向對種子形狀影響不明顯。急尖長苞冷杉冷杉種子較小,擴散距離比較遠,可以為急尖長苞冷杉提供更多的生長空間。陰坡種子千粒重分布范圍為3.32~12.86 g,海拔3 900 m處種子質量達到最優值,平均為10.46 g;陽坡種子千粒重分布范圍為3.72~14.68 g,在海拔3 800 m處最佳,平均為12.37 g;種子千粒重在陰坡與陽坡隨著海拔的升高均呈現先升高后下降的趨勢;總體來看,陽坡種子質量要優于陰坡。急尖長苞冷杉種子在海拔3 800,3 900 m處千粒重表現為顯著差異;在陰坡與陽坡,種子千粒重均表現為顯著差異。在海拔3 700~4 200 m,急尖長苞冷杉天然更新幼苗數量均表現為林隙>林緣>林下。幼苗數量與林分郁閉度、帶翅長度、千粒重、凋落物厚度均呈顯著正相關,苔蘚厚度呈極顯著正相關,但是苔蘚層厚度、凋落物厚度均大于死亡幼苗根長,這樣會使急尖長苞冷杉幼苗缺失水分及其他營養元素而死亡,可見苔蘚層、凋落物層對急尖長苞冷杉幼苗定居和生長發育具有雙重影響。林隙處天然更新幼苗數量較好于林下與林緣,主要是由于林隙內氣溫、土壤溫度及濕度等小氣候因子產生影響,改變了急尖長苞冷杉生長的物理環境和生物環境,對維持色季拉山急尖長苞冷杉種群的穩定性有重要作用。急尖長苞冷杉林天然更新的影響因素還有很多,比如土壤種子庫、自身的生長基質、伴生物種間的競爭、苔蘚及枯枝落葉層的化感作用、自身的化感作用等,因此,對于急尖長苞冷杉天然更新的影響因素還需進一步的研究。
