鹽脅迫對(duì)茄子光合特性、葉綠素?zé)晒饧氨Wo(hù)酶活性的影響
2018-11-09 09:06:30趙躍鋒張清華
山西農(nóng)業(yè)科學(xué) 2018年11期
趙躍鋒,陳 昆,張清華
(商丘市農(nóng)林科學(xué)院蔬菜研究所,河南商丘476000)
大棚和日光溫室等是保護(hù)地栽培的主要方式,但是保護(hù)地土壤長(zhǎng)期受到棚膜覆蓋,土壤不能得到雨水淋洗加劇了鹽分聚集,易出現(xiàn)次生鹽漬化現(xiàn)象,而鹽脅迫可顯著抑制植物的生長(zhǎng)發(fā)育,對(duì)種子形態(tài)建成[1]、生物膜結(jié)構(gòu)及其透性等生理生化指標(biāo)產(chǎn)生不利影響[2]。在鹽脅迫下,植物細(xì)胞會(huì)發(fā)生脫水現(xiàn)象,膜上的酶功能發(fā)生紊亂,各種代謝開始無序進(jìn)行[3],植物細(xì)胞的活性氧增加,光能利用率和CO2同化受到抑制,蛋白質(zhì)與核酸受到損傷,植株發(fā)育進(jìn)程加速,營養(yǎng)生長(zhǎng)期和花期縮短,組織和器官生長(zhǎng)受到抑制[4]。茄子果實(shí)鮮嫩,富含蛋白質(zhì)和多種維生素,營養(yǎng)價(jià)值較高,而保護(hù)地茄子是冬季供應(yīng)市場(chǎng)的主要蔬菜之一,對(duì)于彌補(bǔ)冬季市場(chǎng)空白有重要意義[5]。
本研究從種子萌發(fā)、光合指標(biāo)及葉綠素?zé)晒鈪?shù)等方面深入研究茄子種子萌發(fā)及幼苗生長(zhǎng)發(fā)育對(duì)NaCl脅迫的響應(yīng)特性,旨在探明茄子的耐鹽特性,為溫室茄子栽培提供理論依據(jù)。
1 材料和方法
1.1 試驗(yàn)材料
供試茄子品種為商茄1號(hào),該品種由商丘市農(nóng)林科學(xué)院蔬菜研究所培育,溫室和露地均可種植。
1.2 試驗(yàn)設(shè)計(jì)
2017年11月進(jìn)行穴盤播種,出苗后選取長(zhǎng)勢(shì)健壯、高度基本相同的幼苗,移栽到泡沫箱進(jìn)行鹽脅迫水培試驗(yàn),在塑料箱中添加NaCl濃度分別為0,15,65,115,165 mmol/L的 Hoagland 營養(yǎng)液,每個(gè)泡沫箱栽培24棵,每個(gè)處理5箱,3次重復(fù),處理14 d后對(duì)相關(guān)生理指標(biāo)進(jìn)行測(cè)定。
1.3 測(cè)定項(xiàng)目及方法
茄子幼苗葉片光合速率(Pn)、蒸騰速率(Tr)、氣孔導(dǎo)度(Gs)和胞間 CO2濃度(Ci)采用美國產(chǎn)CIRAS-2便攜式光合儀測(cè)定,每處理測(cè)量3次,3次重復(fù),取平均值。用FMS-2型脈沖調(diào)制式葉綠素?zé)晒夥治鰞x(英國Hansatech公司生產(chǎn))測(cè)定葉綠素?zé)晒鈪?shù)Fo,F(xiàn)m等,測(cè)定時(shí)CO2濃度為(390±10)mmol/L,光照強(qiáng)度為(900±10)lx,溫度(28±2)℃。采用GIANNOPOLITIS等[6]的方法測(cè)定超氧化物歧化酶(SOD)活性,采用ZHANG等[7]的方法測(cè)定過氧化物酶(POD)、過氧化氫酶(CAT)和抗壞血酸過氧化物酶(APX)活性。
1.4 數(shù)據(jù)分析
數(shù)據(jù)圖表處理使用Microsoft Excel 2007軟件,數(shù)據(jù)統(tǒng)計(jì)分析使用SAS軟件。
2 結(jié)果與分析
2.1 鹽脅迫對(duì)茄子幼苗光合特性的影響
由表1可知,不同濃度的鹽脅迫對(duì)茄子幼苗光合特性的影響不同。在15 mmol/L NaCl脅迫下,茄子光合速率低于對(duì)照,但二者差異不顯著;繼續(xù)增加NaCl濃度,光合速率呈逐漸降低的變化趨勢(shì)且顯著低于對(duì)照,至165 mmol/L時(shí),茄子幼苗光合速率最低,僅為對(duì)照的52.67%,說明高濃度的鹽脅迫對(duì)茄子幼苗的光合速率有抑制作用。鹽脅迫條件下,氣孔導(dǎo)度和蒸騰速率的變化趨勢(shì)與光合速率一致,均隨鹽脅迫強(qiáng)度的增加而降低,與光合速率呈現(xiàn)正相關(guān),說明低濃度的鹽脅迫對(duì)茄子幼苗氣孔導(dǎo)度限制較小,高濃度條件下限制性較強(qiáng);胞間CO2濃度隨鹽脅迫強(qiáng)度的增強(qiáng)呈現(xiàn)逐漸升高的變化趨勢(shì),表現(xiàn)出與光合速率負(fù)相關(guān)的關(guān)系,至165 mmol/L時(shí)胞間CO2濃度值最大,較對(duì)照增加56.88%,與對(duì)照間差異極顯著(P<0.01)。
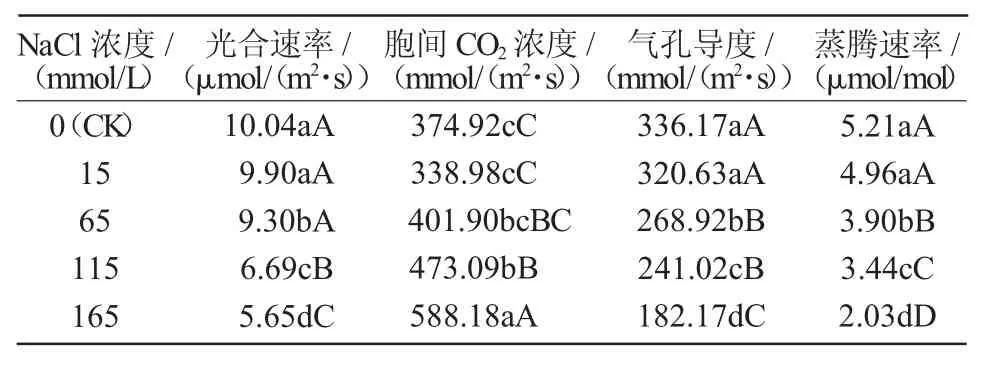
表1 鹽脅迫對(duì)茄子幼苗光合特性的影響
2.2 鹽脅迫對(duì)茄子葉綠素?zé)晒鈪?shù)的影響
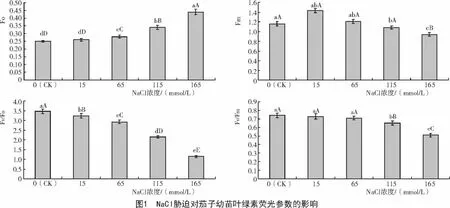
從圖1可以看出,鹽脅迫下Fo均高于對(duì)照,且隨其脅迫強(qiáng)度的增強(qiáng)呈現(xiàn)升高的趨勢(shì),但15 mmol/L條件下,F(xiàn)o與對(duì)照間差異不顯著,表明低鹽條件下對(duì)茄子幼苗葉綠素?fù)p傷較小;當(dāng)NaCl濃度上升到165 mmol/L時(shí),F(xiàn)o極顯著高于對(duì)照,這說明高鹽脅迫能夠?qū)η炎佑酌缛~片PSⅡ造成傷害。暗適應(yīng)下最大熒光產(chǎn)量(Fm)可用于表征PSⅡ的電子傳遞情況。Fm隨鹽脅迫強(qiáng)度的增加呈現(xiàn)單峰變化趨勢(shì),低鹽脅迫(15 mmol/L)Fm高于對(duì)照,繼續(xù)增加NaCl濃度Fm降低,但在0,15,65 mmol/L時(shí),三者差異不顯著;115,165 mmol/L時(shí)顯著低于對(duì)照,較對(duì)照分別降低6.90%和18.97%。從圖1還可以得出,F(xiàn)o與Fm呈現(xiàn)負(fù)相關(guān)關(guān)系。NaCl處理下茄子幼苗Fv/Fo和Fv/Fm二者的變化規(guī)律類似,均隨NaCl脅迫強(qiáng)度的增加而降低,說明鹽脅迫對(duì)茄子葉片PS II的原初光能轉(zhuǎn)化效率有抑制作用,并降低PS II反應(yīng)中心的潛在活性。
2.3 鹽脅迫對(duì)茄子幼苗保護(hù)酶活性的影響
由表2可知,不同濃度的鹽脅迫對(duì)茄子幼苗保護(hù)酶活性的影響存在差異。SOD活性隨脅迫強(qiáng)度的增加逐漸升高,至65 mmol/L時(shí)達(dá)到最大值,較對(duì)照提高35.68%,與對(duì)照差異極顯著(P<0.01);繼續(xù)增加NaCl濃度SOD活性開始降低,至165 mmol/L時(shí)最低,較對(duì)照降低13.84%,可見低濃度的鹽脅迫下植株機(jī)體通過啟動(dòng)保護(hù)系統(tǒng)提高保護(hù)酶活性抵御逆境脅迫,但高濃度的鹽脅迫則使保護(hù)酶受到損傷。POD,CAT變化規(guī)律與SOD類似。APX活性在15 mmol/L時(shí)達(dá)到最大值,較對(duì)照增加89.21%,隨鹽脅迫濃度的增強(qiáng)APX活性開始降低。
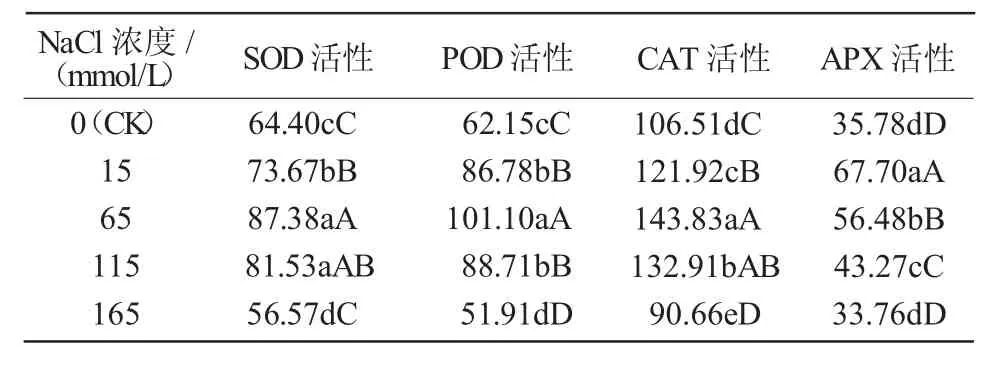
表2鹽脅迫對(duì)茄子幼苗保護(hù)酶活性的影響U/(g·min)
3 結(jié)論與討論
低鹽脅迫下茄子幼苗光合速率、氣孔導(dǎo)度和胞間CO2濃度低于對(duì)照,這表明氣孔限制是致使光合速率降低的主要因素,主要因?yàn)辂}脅迫致使植株細(xì)胞失水發(fā)生滲透脅迫,氣孔關(guān)閉阻礙了外界CO2氣體向細(xì)胞內(nèi)擴(kuò)散,光合底物的減少抑制了光合速率的提高[8]。而在高鹽(NaCl濃度為65~165 mmol/L)條件下,葉片光合速率和氣孔導(dǎo)度低于對(duì)照,而胞間CO2濃度高于對(duì)照,這表明非氣孔限制是此時(shí)光合速率下降的主要因素,主要因?yàn)楦啕}脅迫損傷了葉綠體結(jié)構(gòu),細(xì)胞光合能力下降。
葉綠素?zé)晒饪煞从嘲ü饽艿奈铡⒓ぐl(fā)能的傳遞及光化學(xué)反應(yīng)等在內(nèi)的光合原初反應(yīng)過程,同時(shí)還與電子傳遞、CO2固定和ATP合成有關(guān)[9]。Fo(initial fluorescence)可反映植物葉片光合中心光系統(tǒng)II在完全開放時(shí)的熒光產(chǎn)量[10]。本試驗(yàn)結(jié)果表明,茄子幼苗葉片F(xiàn)o隨鹽脅迫強(qiáng)度的增強(qiáng)而升高,這表明鹽脅迫可能導(dǎo)致PSII與天線色素(LHC)分離[11]。Fm(maximumfluorescence)可反映光照下葉片最大的可變熒光值。Fm在鹽脅迫下呈降低趨勢(shì),表明逆境使葉片受到光抑制[12]。茄子幼苗葉片F(xiàn)v/Fo和Fv/Fm低于對(duì)照,說明NaCl作為逆境因子降低了葉片PSII反應(yīng)中心捕獲光能的效率,這進(jìn)一步解釋了光合速率降低的原因。
超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、過氧化物酶(POD)等是植物體保護(hù)酶系統(tǒng)中重要的保護(hù)酶[13],對(duì)保持細(xì)胞內(nèi)活性氧產(chǎn)生與清除之間的平衡、維持細(xì)胞的完整與穩(wěn)定有重要作用,并能提高植物對(duì)逆境的適應(yīng)能力。周靜等[14]通過對(duì)辣椒幼苗的研究指出,隨著NaCl濃度的增加,辣椒幼苗葉片SOD,POD和CAT活性呈現(xiàn)先升高再降低的變化趨勢(shì)。克熱木伊力等[15]研究表明,阿月混子葉片SOD,CAT,POD活性在鹽脅迫0~250 mmol/L時(shí)逐漸升高,濃度繼續(xù)升高上述指標(biāo)降低。本試驗(yàn)結(jié)果表明,4種保護(hù)酶均隨鹽脅迫強(qiáng)度的增加呈現(xiàn)先升高后降低的變化規(guī)律,與前人研究結(jié)論一致,這可能因?yàn)辂}脅迫增強(qiáng)了抗氧化酶基因在轉(zhuǎn)錄水平上的表達(dá)[16],而高濃度的鹽脅迫造成膜損傷破壞了保護(hù)酶系統(tǒng)。
