新疆部分棉區棉花黃萎病病菌致病力分化的遺傳多樣性分析
2018-10-13 03:27:44李文雯龐學兵易小龍張夢恬王愛英
江蘇農業科學 2018年17期
李文雯, 龐學兵, 易小龍, 李 國, 張夢恬, 王愛英
(石河子大學生命科學學院/農業生物技術重點實驗室,新疆石河子 832000
棉花黃萎病是危害新疆棉花生產的重要因素,而大麗輪枝菌(Verticiliumdahliae)是引發棉花黃萎病最重要的致病菌,因此對其菌落形態、遺傳進化和致病力之間進行相關性分析,探討菌株的起源和發展以及控制策略尤其重要。然而僅靠傳統的形態學分類和致病性檢測進行種間及種內鑒定,難以準確揭示物種間的進化關系。分子標記的高速發展,使得在基因水平上探討物種進化成為可能。多種分子標記如簡單重復序列(simple sequence repeat,簡稱SSR)、簡單重復序列間隔區(inter-simple sequence repeat,簡稱ISSR)、限制性片段多態性(amplified fragment length polymorphism,簡稱AFLP)、單核苷酸多態性(single nucleotide polymorphisms,簡稱SNP)[1-6]等已經被廣泛用于致病菌的遺傳多樣性、致病力、區域來源的相關性研究以及物種起源的推測等。我國目前對大麗輪枝菌的研究大多收集的是不同生態環境與不同寄主來源的菌株,采集更多菌株能更好地豐富我國的致病菌資源。不同地區的菌落形態分布也會顯著不同。目前的研究大多從生物學角度根據大麗輪枝菌在馬鈴薯葡萄糖瓊脂(potato dextrose agar,簡稱PDA)培養基上的菌落形態將其劃分為3種培養類型,即菌絲型、菌核型、中間型[7-8]。本研究通過簡單重復序列標記分析新疆部分棉區棉花黃萎病病菌(大麗輪枝菌)與其區域來源、致病力的關系,并探討其在新疆棉區的菌落形態分布情況。從生物學、系統發育學水平對新疆棉花黃萎病病菌進行系統研究,以期為致病菌的遺傳分化和系統發育研究奠定理論基礎。
1 材料與方法
1.1 試驗材料
棉花品種為新陸早7號(感病品種)、中棉35(耐病品種)、新陸早33號(抗病品種),由筆者所在實驗室保存。
1.2 試驗方法
1.2.1 菌株來源 2015年7月下旬至2016年8月上旬在新疆南疆和北疆的部分棉區采集病樣,并在流水下沖洗 1 h,再將其放入75%乙醇中浸泡1 min,0.1% HgCl2表面消毒2~5 min,用蒸餾水沖洗3次,無菌濾紙吸干水分。在超凈工作臺上將樣品剪成0.5 cm的小莖段,然后放置于PDA培養基上并于25 ℃條件下進行暗培養。待培養基上長出白色菌絲時,打取直徑為 5 mm 的菌餅轉接種到新鮮的PDA培養基上。培養7 d后,用無菌水洗下孢子制成孢子懸浮液,吸取200 μL孢子懸浮液均勻涂布于PDA培養基上,培養條件同上,然后用打孔器取單菌落,分離純化得到菌株。將純化的菌株接種于液體查氏培養基中,并于25 ℃、160 r/min條件下暗培養7 d,搖勻用移液槍吸取孢子懸浮液置于血球計數板上,然后在40倍顯微鏡下觀察計數。用蒸餾水將孢子懸浮液濃度稀釋為1×106CFU/mL,用于致病力檢測試驗。
1.2.2 黃萎病病菌致病力檢測 采用撕底蘸根法檢測黃萎病病菌的致病力,具體操作為用硫酸對棉花種子進行脫絨,然后在50~60 ℃的溫水中浸泡催芽2 h,播種于直徑為15 cm的紙缽(盛有蛭石與營養土,其體積比為1 ∶1)中,每個紙缽播種2~3粒種子,置于光—暗周期為8 h—16 h、溫度為 25 ℃ 的溫室中培養。待棉花長出2張真葉時接種大麗輪枝菌孢子懸浮液,每個紙缽接種50 mL,每個菌株接種3個紙缽,侵染前剪切棉花須根形成傷口,以利于病菌侵染。接種后每7 d統計1次發病情況,連續統計1個月。最后計算病情指數,分析不同地區間的致病力差異。發病病級的確定參照文獻[9],以病情指數<25.0為弱致病力菌株,25.0≤病情指數≤35.5為中等致病力菌株,病情指數>35.5為強致病力菌株。
病情指數=100×∑(各級病株數×相應病級)/(調查總株數×最高病級)。
1.2.3 黃萎病病菌的形態觀察 將不同來源的棉花黃萎病病菌菌株接種于PDA培養基上活化,吸取孢子懸浮液涂布于PDA培養基上,挑取單菌落置于培養基上,倒置于培養箱內,25 ℃暗培養7 d后觀察,根據菌落是否產生微菌核以及微菌核產量的多少,將菌株劃分為菌核型(微菌核產量多,菌絲濃密)、菌絲型(不產微菌核)、中間型(微菌核產量少,周邊膜狀),分析各個菌落類型在不同致病力類群中的占比,進而確定致病力和培養類型的關系。每個菌株3次重復。
1.2.4 遺傳多樣性分析
1.2.4.1 基因組DNA的提取 按照Raeder等的十六烷基三甲基溴化銨(hexadecyl trimethyl ammonium bromide,簡稱CTAB)法[10]提取棉花黃萎病病菌的DNA,用于后續SSR分子標記的檢測。
1.2.4.2 SSR分子標記檢測 使用實驗室前期篩選得到的9條SSR特異性引物(表1)對46株大麗輪枝菌進行SSR檢測,采用0、1矩陣記錄條帶數。二進制矩陣記錄了46個菌株9條引物產生的所有條帶。使用POPGENE32計算各個棉區大麗輪枝菌的遺傳多樣性,分析各個地區間的菌株遺傳多樣性與菌株致病力類型分布的關系。使用GenALEX 6.502中的AMONA和PCOA分析種群內和種群間的基因變異得到多樣性數值(多樣性分化系數Phi PT),進一步分析得到多樣性分布。根據“1.2.2”節的致病力劃分標準,將菌株劃分為強、中、弱等3個致病力類型,計算各個致病類型在基因多樣性上存在的區別。根據不同的區域來源將菌株劃分為4個地區(和碩縣、焉耆回族自治縣、瑪納斯縣、呼圖壁縣等)類型,根據多樣性數值計算4個地區類型之間和類型內的多樣性百分率,以了解不同致病力和不同地區來源是否具有相關性。擴增體系為25 μL,其中包含1.0 μL模板、13.5 μL 2×EasyTaqSuperm Mix,各1.0 μL 上、下游引物,用ddH2O補齊25.0 μL。

表1 不同引物的最佳退火溫度
擴增參數為95 ℃預變性 5 min;95 ℃變性1 min,退火(退火溫度見表1)1 min,72 ℃延伸3 min,30個循環;72 ℃延伸 10 min。
2 結果與分析
2.1 供試菌株的培養類型和致病力檢測
采用隨機分點采樣法從新疆南疆和北疆部分棉區的發病棉田分別采集57株棉花病株,分離、純化、鑒定得到46株致病力不同的大麗輪枝菌,具體見表2。
注:++表示強致病力;+表示中等致病力;-表示弱致病力。
2.2 致病力檢測分析
由表3可知,4個地區棉花黃萎病強致病力菌株占總體菌株的32.61%,中等致病力菌株占43.48%,弱致病力菌株占23.91%。說明新疆棉區的棉花黃萎病病菌以中等致病力菌株為主,強致病力菌株所占比例次之,弱致病力菌株較少。瑪納斯棉區的棉花黃萎病病菌強致病力菌株所占比例最高,呼圖壁棉區的強致病力菌株所占的比例最少。
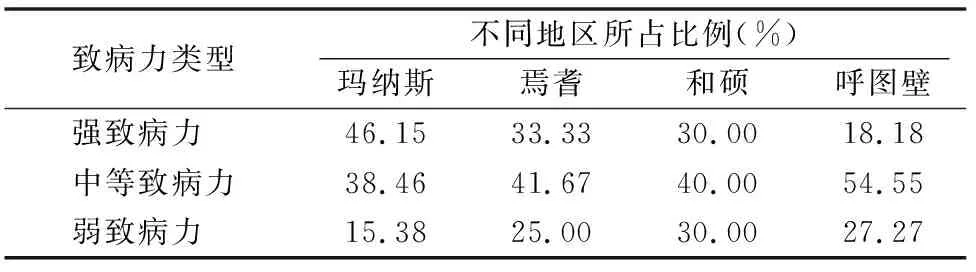
表3 不同地區致病力類型所占供試菌株的比例
2.3 不同致病力菌株的菌落培養類型分析
由表4可知,大部分菌株屬于菌核型,占比為52.17%;菌絲型次之,占比為26.09%;中間型最少,占比為21.74%。說明菌核型菌株屬于新疆地區普遍存在的優勢種群。強致病力類型中的菌核型菌株和菌絲型菌株分別占該致病力類型總株數的66.67%、13.33%,中等致病力類型中的菌核型和菌絲型分別占該致病力類型總株數的50.00%、30.00%,弱致病力類型中的菌核型和菌絲型類型均占該致病力類型總株數的36.36%。

表4 46株大麗輪枝菌的培養類型在不同致病力類型里的分布情況
3個致病力類型雖然都出現了菌絲型和菌核型菌株,但不同培養類型的致病力類型分布不同,其中在強致病力類型和中等致病力類型中菌核型菌株占據主導地位,菌絲型菌株在弱致病力類型中相對于其他2種致病力類型占據較多比例,且隨著致病力類型總體致病力的逐漸減弱,菌核型比例減少,菌絲型比例增大,說明菌落的形態與黃萎病病菌的致病力類型有一定的相關性。
2.4 遺傳多樣性分析
將試驗所得的檢測條帶先在Excel中編輯成0、1矩陣,然后將0、1矩陣轉換成txt文件,導入到POPGENE32中進行dominant模塊計算,得到4個不同棉區來源的棉花黃萎病病菌菌株類型間的遺傳分化系數(Gst)為0.243 3,每代遷移數(Nm)為1.554 9,群體總基因多樣度(Ht)為0.336 1,各棉區群體內基因多樣度(Hs)為0.254 3。瑪納斯、焉耆、呼圖壁、和碩等4個地區的Shannon’s信息指數(Ⅰ)分別為0.421 3、0.338 5、0.356 8、0.429 3,Nei’s遺傳多樣性指數(H)分別為0.277 9、0.221 6、0.232 7、0.285 2。分析發現,焉耆地區的Shannon’s信息指數和Nei’s遺傳多樣性指數最低,說明該地區黃萎病病菌菌株之間具有較高的親緣關系。
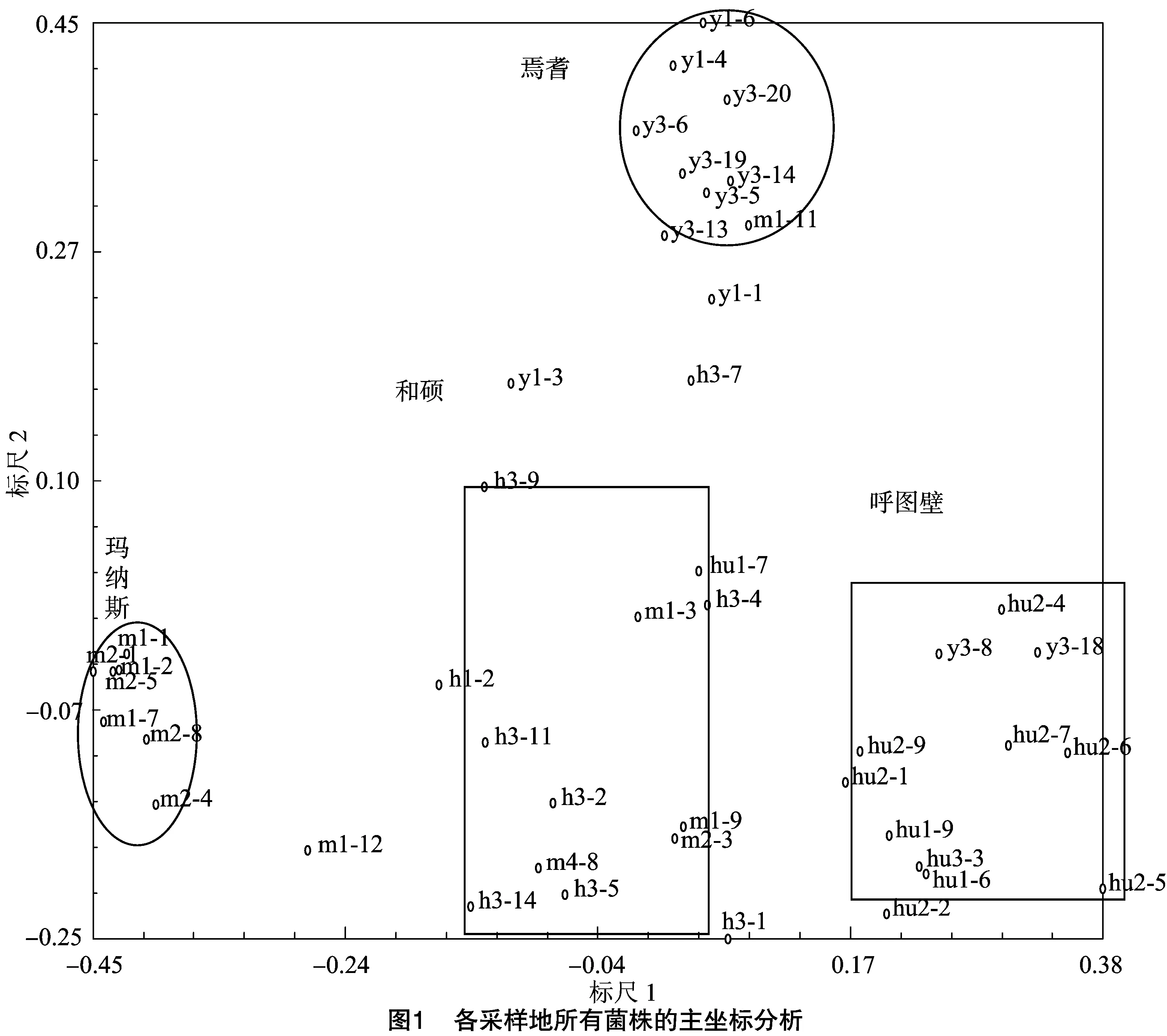
使用NTSYS軟件基于46株菌株的遺傳相似系數進行主坐標分析,得到各個菌株之間的二維分布。由圖1可知,瑪納斯棉區與呼圖壁棉區的菌株地理位置較近,但遺傳關系卻相對較遠。瑪納斯與和碩棉區的菌株親緣關系較近。各個致病力類型和培養類型相同的菌株沒有聚攏在一起,分布松散。但同一地區的菌株能夠大致聚集在一起,在焉耆和瑪納斯地區,來自相同棉區菌株之間的親緣關系較近,因此各個菌株聚集得較為緊密,而和碩與呼圖壁地區菌株的親緣關系較遠,因此各個菌株分布松散。同時可以看出,焉耆地區和其他3個地區之間的菌株親緣關系較遠。
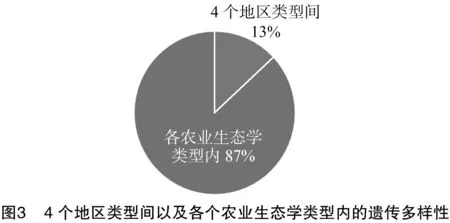
3種不同致病力類型之間的基因多樣性差異不大。由圖2可知,不同類型間的基因多樣性(6%)遠遠小于不同類型內的基因多樣性(94%),說明致病力不同在多樣性上體現微弱的差異,因此基因多樣性分子標記不能作為致病力評估指標。由圖3可知,4個區域來源的菌株在同一個地區內的變異較大(基因多樣性為87%),不同地區間的變異較小(基因多樣性只有13%)。雖然地區間的基因多樣性有差異,但是這種差異極小,遠遠低于地區內,但也可以說明菌株的基因多樣性與區域來源有一定的相關性。POPGENE32的分析結果也顯示,基因流動使得個體遷移對菌株基因多樣性起到一定作用。不同區域來源的黃萎病病菌沒有因為區域來源不同而形成地理隔離種群結構,說明地理環境的自然選擇壓力未造成菌株的基因差異。
3 討論
本研究的菌株致病力類型與培養性狀類型之間存在一定的相關性,強致病力菌株大多為菌核型,而弱致病力菌株中菌絲型較多。在3個致病力類型中,中等致病力菌株在4個采樣地都有廣泛分布,可能是由于在侵染過程中發病比強致病力型菌株輕緩,具有更大的繁殖潛力,在植物未致死的情況下可以獲得更大的生物量,保存在病殘體中作為來年的侵染源。弱致病力菌株由于毒力較弱,定殖量較少。因此,中等致病力菌株成為優勢種群。
遺傳多樣性分析發現,類群內的多樣性明顯高于類群間的多樣性,說明不同棉區來源的大麗輪枝菌菌株間存在一定的基因流動,也說明總體的遺傳分化豐富。多態性較豐富的地區如和碩,強致病力菌株占比沒有明顯大于其他地區;而多態性較低的地區如焉耆,弱致病力菌株占比也沒有明顯大于其他地區。菌株多樣性指數較高的類型,沒有相對較強的致病力。因此認為,種群的基因多樣性高低不會對致病力的強弱產生影響,進一步證明遺傳多樣性沒有在大麗輪枝菌的致病力強弱方面表現出明顯差異。
菌株多樣性分析結果顯示,焉耆和呼圖壁雖然在地理位置上相聚較近,然而卻在遺傳多樣性分化上表現出很遠的親緣關系。北疆地區的瑪納斯和和碩在地理位置上相聚較遠,在遺傳多樣性聚類中卻表現出比較近的親緣關系,因此大麗輪枝菌沒有在不同的地理位置產生與多樣性相關的差異性。但基因相似性系數得到的主坐標分析圖顯示各個地區各個菌株相互之間都存在不同程度的豐富多態性,說明同一地區的各個菌株之間遺傳分化嚴重,多樣的遺傳分化也為菌株適應過程中的致病力分化改變提供了可能。
棉花黃萎病病菌的不同致病力類型間以及種群內的遺傳多樣性十分豐富,卻不能用于致病力強弱的辨別,遺傳多樣性與致病力強弱的相關性不是十分明顯,可以判斷這種遺傳多樣性不是由致病力強弱差異所引起。而遺傳多樣性和棉區區域來源表現出一定的相關性,可能是由不同棉區種植的棉花品種不同所導致的[11-12]。除此之外,致病力分化的菌株遺傳多樣性還有其他來源,例如突變、基因漂移以及有性、無性重組等,因此病原微生物可以在種內產生遺傳多樣性[13]。現有資料顯示,某些植物內生微生物在與宿主發生聯系時,可增強寄主的抗病性、提高植物的生命力[14]。反過來寄主較高的多樣性也會引起病原菌新基因型的出現,并且伴隨新的生物學特性,例如病原菌的適應性和對殺真菌劑的抗性等。本研究中和碩地區菌株的遺傳多樣性較高,說明該地區菌株的適應力相對其他地區強,有更廣泛的分布;而新疆焉耆地區遺傳多樣性低,推測大麗輪枝菌還沒有充分和寄主作用而發生變異,可能是由于引入大麗輪枝菌的時間相對較短,該地區棉花黃萎病發生的時間相比其他地區較晚。

4 結論
在新疆部分主要產棉地區采集了黃萎病的田間病株,對其病原菌進行培養類型觀察,通過遺傳多樣性和致病力分化研究發現,致病力強弱與菌株的培養類型有關聯性,強致病力類型存在更多的菌核型菌株。分子標記的遺傳多態性很豐富,由此可以說明,多態性可以反映其致病力多樣化的進程,部分地區采用輪作的種植模式,可能會為病原菌提供協同進化的相關寄主,而寄主的多元化將促進病原菌的分化。瑪納斯地區以強致病力類型菌株為主,呼圖壁的菌株以中等致病力類型為主。新疆地區的菌株多樣性和病株來源地有一定的相關性,但基因的多樣性標記不能作為不同致病力的鑒別標志。各個致病菌的變異類型在不同棉區分布廣泛且適應性強,反映了不同菌株致病力的分化潛力,可為進一步探討棉花黃萎病病菌遺傳變異提供方向。
