中亞熱帶不同母質和森林類型土壤生態酶化學計量特征
2018-09-28 08:45:02張星星楊柳明李一清林燕語鄭憲志楚海燕楊玉盛
生態學報 2018年16期
張星星,楊柳明,*,陳 忠,李一清,3,林燕語,鄭憲志,楚海燕,楊玉盛
1 福建師范大學地理科學學院,福州 350007 2 濕潤亞熱帶山地生態國家重點實驗室培育基地,福州 350007 3 夏威夷大學希羅分校農林和自然資源管理學院,希羅 96720
土壤微生物和植物分泌的胞外酶是土壤有機質分解的關鍵媒介,是細胞代謝的產物,其活性與產量和碳底物及氮、磷養分有效性密切相關[1- 2]。通常認為,微生物用于獲取碳、氮、磷的主要胞外酶比值(βG∶NAG∶AP),即生態酶化學計量比,能夠反映微生物群落的新陳代謝及養分需求與環境中養分有效性之間的生物地球化學平衡模式,這可以用來衡量土壤微生物能量和養分資源限制狀況[3- 4]。
現有研究表明,多種生物因子和非生物因子都能夠影響土壤生態酶化學計量比。氣候通過調節土壤微生物的生長速率和養分有效性從而改變土壤生態酶化學計量比[5- 6],并且土壤生態酶化學計量比亦受土壤養分狀態、pH[7]、土壤C∶N∶P以及土壤質地[8]等非生物因子的影響;例如Peng等在中國北方溫帶草原研究發現土壤養分有效性主要影響了土壤生態酶化學計量[2]。此外,其亦可能受生物因子的影響,如地上生物量[9]、根生物量[10]以及微生物生物量化學計量比[11]的影響,Allison等研究亦表明微生物量C∶N∶P比值不同的微生物群落能夠通過分泌特定的胞外酶從復雜的基質中獲取限制性養分,從而改變土壤生態酶化學計量比[12]。雖然,生物和非生物因子對土壤生態酶化學計量比的影響已經受到關注,但是各種因子對土壤生態酶化學計量比影響的貢獻比例仍不明確。
土壤母質和森林類型都是影響森林土壤養分有效性的重要因素。土壤母質決定土壤養分的初始供應,并且影響土壤對養分的保持能力[13],如趙汝東等[14]研究發現花崗巖發育的馬尾松人工林土壤養分、生化強度、磷酸酶活性等較砂巖發育的土壤高。而森林類型則通過不同凋落物輸入改變土壤養分的有效性和循環。如楊玉盛[15]研究表明天然常綠闊葉林轉變為杉木人工林,土壤C、N、P等養分流失及生產力下降,這主要是由于天然林轉變成人工林后,凋落物數量顯著降低的結果。關于土壤母質和森林類型對土壤養分的影響已有大量報道,然而這兩種因素將如何通過調控土壤生物和非生物因子,從而對土壤生態酶化學計量特征產生影響仍需要深入研究。因此,本研究分別選取了砂巖和花崗巖土壤的米櫧林和杉木林為研究對象,測定了土壤的物理化學性質、微生物生物量碳、氮和磷以及酶活性,研究母巖和森林類型對土壤酶活性及生態酶化學計量特征的影響,有助于完善土壤生態酶化學計量比的理論研究體系,為認識區域土壤微生物能量和養分資源限制狀況提供新的視角。
1 材料與方法
1.1 研究區域概況
試驗區位于格氏栲自然保護區(26°11′N,117°28′E)和金絲灣森林公園陳大林業采育場(26°19′N,117°36′E),兩者在直線距離上不超過30 km,在行政區劃上二者都屬于福建省三明市。三明市區域內,平均海波300 m,平均坡度25°—35°,其屬中亞熱帶季風氣候,具有冬冷夏熱、雨熱同季的特點,多年平均氣溫20.1℃,年降水量1670 mm,年均蒸發量1585 mm,相對濕度81%,其中格氏栲自然保護區土壤以砂巖發育的紅壤為主,金絲灣公園陳大林業采育場土壤以花崗巖發育的紅壤為主。
本研究分別選取了格氏栲自然保護區和金絲灣森林公園陳大林業采育場的米櫧林和杉木林。其中格氏栲自然保護區的米櫧林約200年沒有受到人為的干擾,建群樹種為米櫧,而米櫧(Castanopsiscarlesii),格氏栲(Castanopsiskawakamii),杜英(Elaeocarpusdecipiens),木荷(Schimasuperba)等為主要樹種。杉木林是經過米櫧林皆伐、火燒等形成的人工純林,而杜莖山(Maesajaponica),毛冬青(Ilexpubescens),華山姜(Alpiniachinensis)等為主要林下植被。金絲灣森林公園陳大林業采育場的米櫧林約50年沒有受到人為干擾,建群樹種為米櫧,其中米櫧(Castanopsiscarlesii)、閩粵栲(Castanopsisfissa)、黃丹木姜子(Litseaelongata)、新木姜子(Neolitseaaurata)、木莢紅豆(Ormosiaxylocarpa)等為主要樹種。杉木林是經過米櫧林皆伐、火燒等形成的人工純林,狗骨柴(Tricalysiadubia)、毛冬青(Ilexpubescens)、芒萁(Dicranopterisdichotoma)等為主要林下植被。
1.2 研究方法
1.2.1 土壤采集
土壤樣品于2016年3月分別在三明格氏栲自然保護區和金絲灣公園陳大林業采育場的米櫧林和杉木林4個樣地中采集(每個樣地有3個20 m×20 m的樣方)。在每個樣方中用土鉆按樣方“S”型隨機取15—20個點,并在0—10 cm土層取樣。每個樣方的土壤混合均勻后放入便攜式冰盒中并立即帶回實驗室。去除土壤中可見的根及石礫等,分為兩部分,一部分儲存在4℃冰箱中,用于土壤酶活性的檢測,另外一部分土樣過2 mm的篩后再分為2部分,一部分儲存在4℃冰箱中,用于土壤微生物量的檢測,其余土樣風干后,用于其他基本理化性質的分析。
1.2.2 土壤理化性質的測定
土壤有機碳和全氮采用碳氮元素分析儀(Elementar Vario EL Ⅲ)測定;土壤總磷采用HClO4-H2SO4消煮-鉬銻抗比色法提取后,用連續流動分析儀(Skalar san++,Skalar,荷蘭)測定;土壤質地采用激光粒度儀(MasterSizer 2000,Malvern,Englang)測定。
1.2.3 土壤微生物量的測定
土壤微生物量采用氯仿熏蒸-浸提法[16- 17],稱取兩份5 g過2 mm篩的鮮土,分別作為熏蒸和未熏蒸處理。微生物量碳、氮(MBC、MBN)用0.5 mol/L K2SO4溶液浸提,其中4∶1的水土比,震蕩離心后,其上清液過0.45 μm濾膜抽濾。浸提液分別用總有機碳分析儀(TOC-VCPH/CPN,Shimadzu,日本)測定微生物量碳,使用連續流動分析儀(San++, Skalar,荷蘭)測定微生物量氮。土壤微生物量磷(MBP)用0.5 mol/L NaHCO3溶液浸提,其中4∶1的水土比,震蕩離心后,其上清液過濾紙后,用連續流動分析儀(San++, Skalar,荷蘭)測定微生物量磷。微生物量碳、氮、磷的計算公式分別如下:
MBC=△EC/kC;MBN=△EN/kN;MBP=△EP/kP
式中,△EC為熏蒸和未熏蒸的土壤有機碳含量的差值,kC為轉換系數0.38;△EN為熏蒸和未熏蒸土壤有機氮含量的差值,kN為轉換系數0.45;△EP為熏蒸和未熏蒸為土壤磷含量的差值,kP為轉換系數0.4。
1.2.4 土壤生態酶活性測定
參照Saiya-Cork等[18]研究方法測定3種水解酶的活性(AP、βG和NAG),用傘形酮(MUB)做標示底物,稱取1 g土壤放于含有125 mL 50 mmol/L、pH值為5.0的醋酸緩沖液的廣口瓶中,用磁力攪拌器攪拌5分鐘使其均質化。攪拌后將懸浮液用移液器取200 μL到96孔微孔板。微孔板在黑暗中20℃孵育4 h后,向每個微孔板中加入10 μL、1 mol/L NaOH溶液使其反應停止,用多功能酶標儀(Synergy H4,美國)測定水解酶的熒光度。各種酶縮寫及所用底物見下表1。
1.3 數據統計
因為亮氨酸氨基肽酶的含量(LAP)通常較低,本文根據Turner等[19]的研究方法,將分解幾丁質的β-N-乙酰氨基葡萄糖苷酶(β-N-acetylglucosaminidase NAG)的氮相關酶作為氮循環的特征酶進行計算。C∶N酶活性的比值,通過βG:NAG進行計算,C∶P酶活性的比值,通過βG:AP進行計算,N∶P酶活性的比值,通過NAG:AP進行計算。用Excel 2013軟件對數據進行處理后,用SPSS 19.0和Canoco Software 5.0軟件對數據進行分析,主要由Origin 9.0進行繪圖。不同母巖發育的土壤生物及非生物因子屬性的差異使用單因素方差分析(One-way ANOVA),米櫧林和杉木林土壤酶活性和生態酶化學計量比的差異采用獨立樣本T檢驗,母質、森林類型及母質和森林類型的交互作用對酶活性及生態酶化學計量比的影響采用雙因素方差分析(two-way ANOVA),影響酶活性的環境因素均采用Pearson相關分析和冗余分析(RDA)來分析。
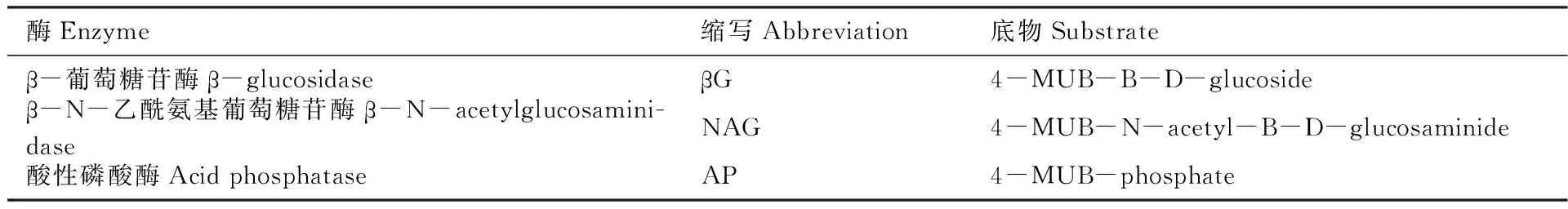
表1 土壤酶活性的種類、縮寫和所用底物
2 結果與分析
2.1 不同森林類型土壤非生物因子和生物因子屬性
土壤母質和森林類型均影響土壤非生物及生物因子(表2)。總體上,花崗巖發育的土壤pH、砂粒均高于砂巖發育的土壤,與此相反,TP、TN、粉粒和粘粒低于砂巖發育的土壤。其中,砂巖發育的土壤米櫧林SOC、TN、TP、MBC、MBN和MBP的含量均高于杉木林,花崗巖發育的土壤則表現出相反的趨勢,即米櫧林SOC、TN、TP、MBC、MBN和MBP的含量均低于杉木林,結果表明土壤pH、TP及質地組成可能主要取決于土壤母質,而SOC、TN和其它生物因子主要是土壤母質和林分類型共同作用的結果。
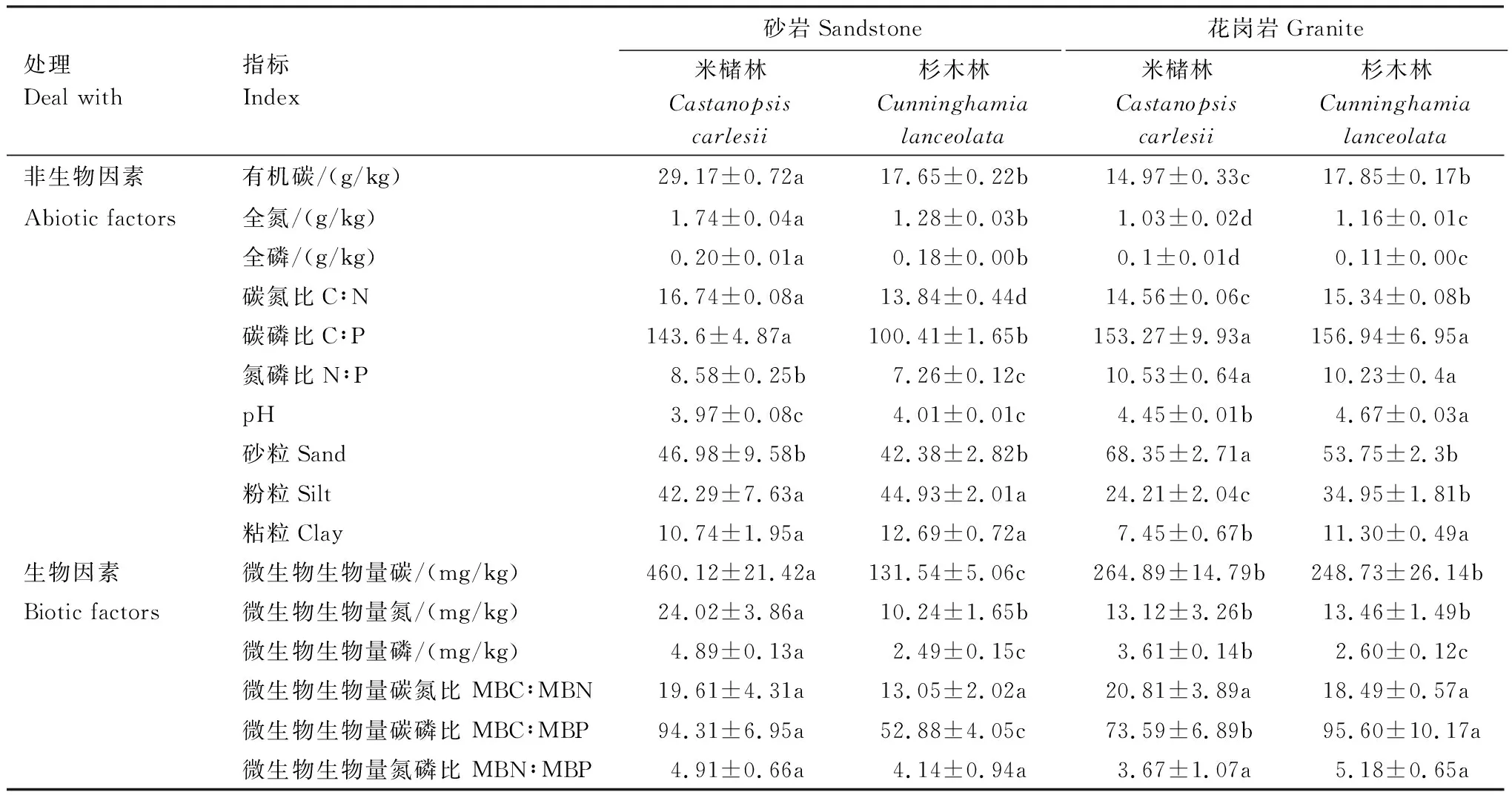
表2 不同森林類型土壤非生物因子和生物因子的含量
表中數值為平均值±標準差,同行不同小寫字母表示相同指標不同林分間的差異顯著(P< 0.05)
2.2 土壤母質和森林類型對土壤酶活性的影響
土壤母質和森林類型顯著影響土壤AP、βG和NAG 3種土壤酶活性(圖1),并且母質和森林類型存在交互作用(表3)。AP、βG和NAG 3種酶活性的變化范圍分別為77.04—257.52、12.73—59.42、33.52—86.30 nmol g-1h-1,其中砂巖發育的土壤,米櫧林βG和NAG活性顯著低于杉木林,而AP的活性則高于杉木林,花崗巖發育的土壤βG、NAG和AP活性顯著高于杉木林。此外,砂巖發育的土壤AP活性顯著低于花崗巖發育的土壤,說明相對于砂巖發育的土壤,花崗巖發育的土壤中微生物受磷養分限制的強度更大,而土壤母質對βG和NAG活性影響則因林分類型的不同而不同。
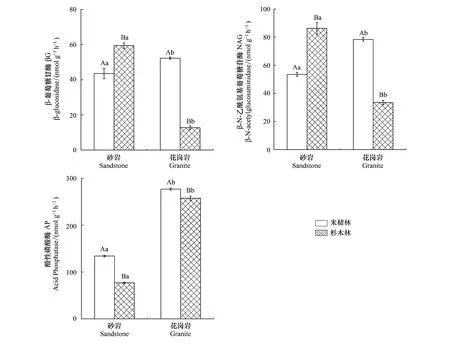
圖1 不同森林類型土壤酶活性的含量
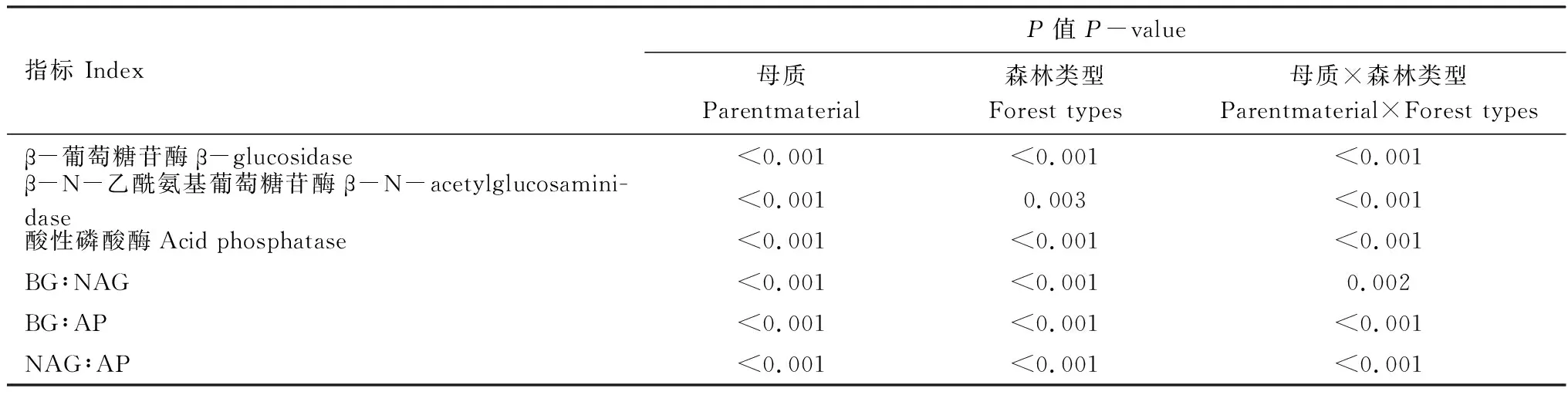
表3 母質、森林類型及其交互作用對土壤酶活性和生態酶的化學計量比影響的雙因素方差分析
2.3 土壤母質和森林類型對土壤生態酶化學計量比的影響
土壤母質和森林類型亦顯著影響土壤生態酶化學計量比(圖2),并且母質和森林類型存在交互作用(表3)。βG:NAG、βG:AP和NAG:AP的變化范圍分別為0.38—0.81、0.05—0.77、0.13—1.12,其中砂巖發育的土壤,米櫧林βG:AP和NAG:AP顯著低于杉木林,而βG:NAG則顯著高于杉木林。花崗巖發育的土壤,米櫧林βG:NAG、βG:AP 和NAG:AP都顯著高于杉木林,并且砂巖發育的土壤βG:AP和NAG:AP都顯著高于花崗巖發育的土壤。
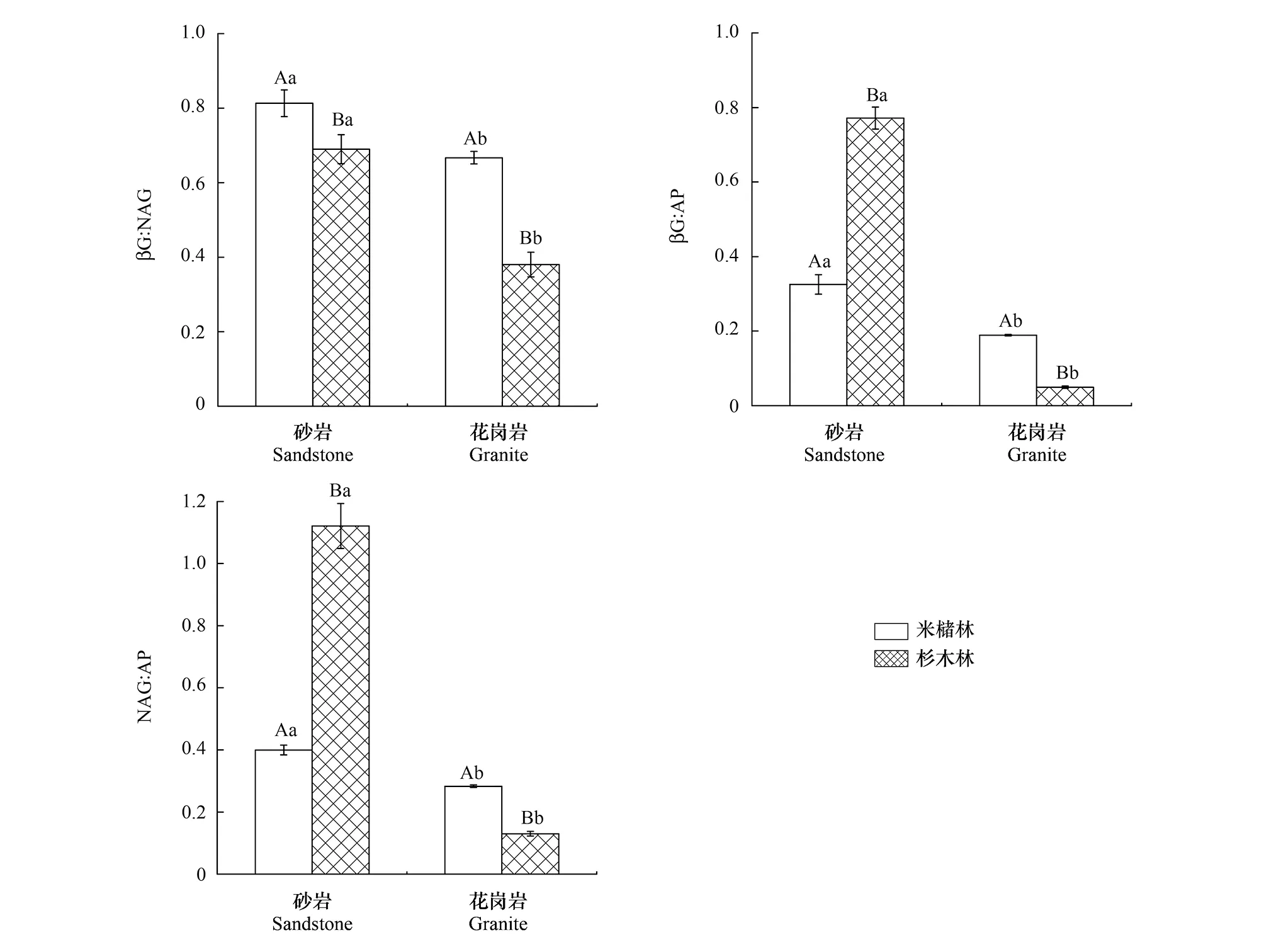
圖2 不同森林類型土壤生態酶化學計量比
2.4 土壤生態酶化學計量比與土壤非生物和生物因子的相關性
以土壤酶活性和生態酶化學計量比為響應變量,以生物因子和非生物因子為解釋變量進行RDA分析,第一軸解釋了變量的77.87%,第二軸解釋了變量的19.96%,并且土壤pH、TP和MBC分別解釋土壤酶活性和生態酶化學計量比的56.9%、27.9%和12.3%(圖3),此外,從RDA分析可知,土壤酶活性和生態酶化學計量比可以明顯的將四種林分類型區分開。相關性分析結果表明,土壤βG:AP和NAG:AP與C∶P、N∶P、MBC∶MBP、MBC∶MBN以及砂粒呈顯著的負相關關系,土壤βG:NAG、βG:AP和NAG:AP與pH呈顯著的負相關關系,TP與土壤βG:NAG、βG:AP呈顯著的正相關關系(表4)。
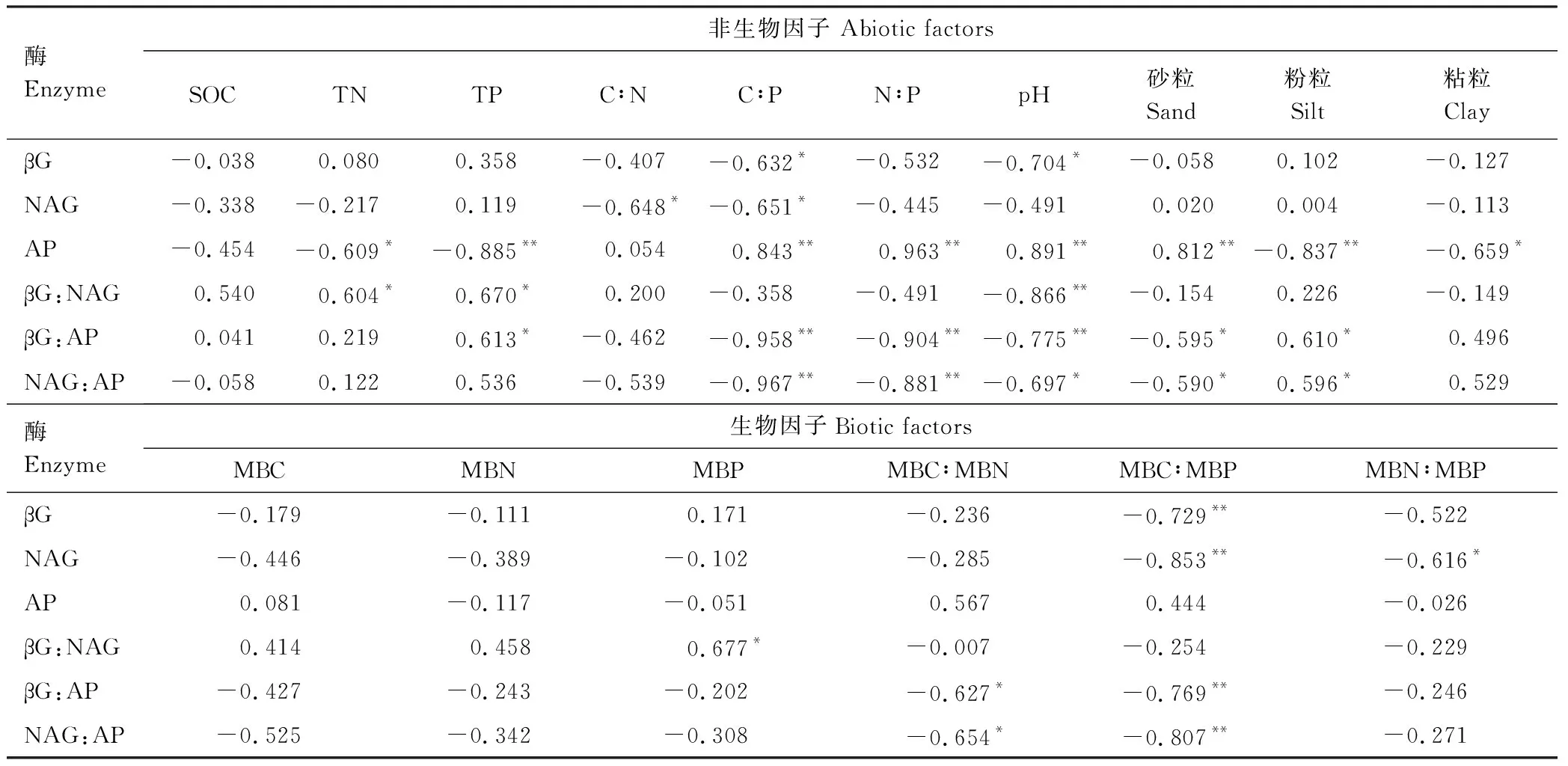
表4 土壤酶活性、生態酶化學計量比與非生物和生物因子的相關性系數
*:P<0.05;**:P<0.01
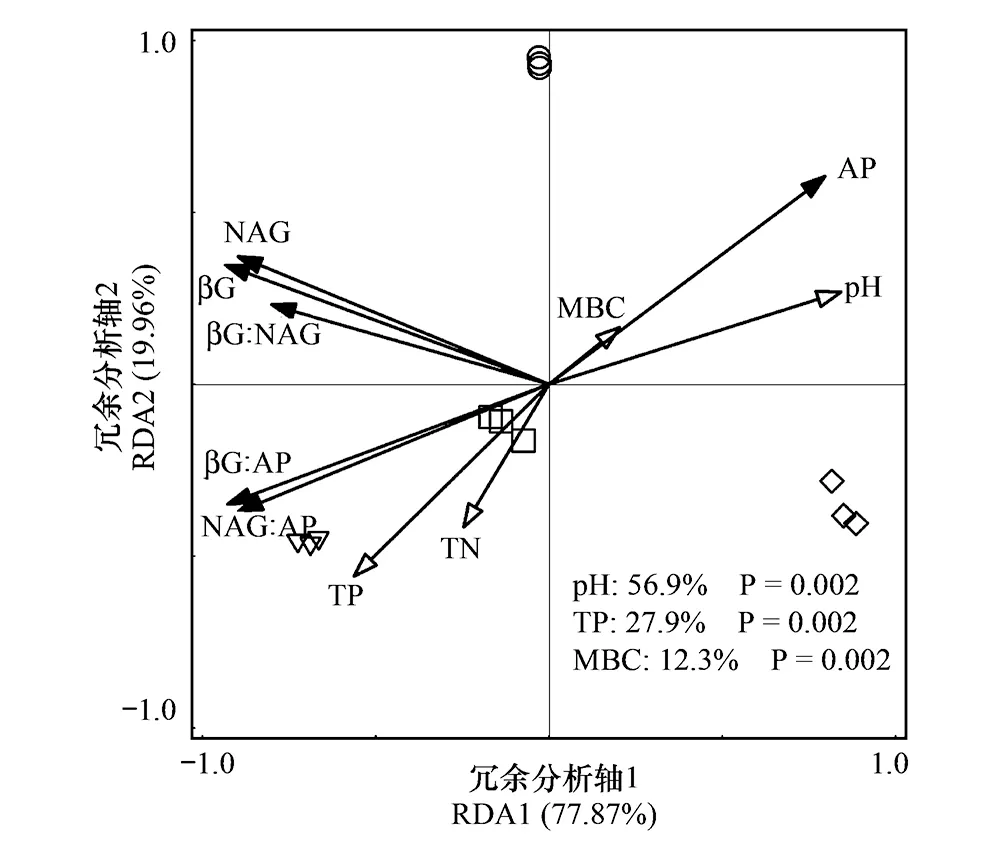
圖3 土壤酶活性和生態酶化學計量比與非生物及生物因子的冗余分析
3 討論
3.1 土壤生態酶化學計量比特征與養分限制
土壤βG∶NAG∶AP對數轉換后的比值為1∶1.14∶1.42,與全球生態系統1∶1∶1的比值相偏離,其中βG∶AP平均值為0.33,低于全球水平0.62[7],而NAG∶AP平均值為0.48,與全球水平0.44[7]相接近,說明相對于土壤碳底物有效性來說,本區域土壤微生物可能主要存在一定的磷限制,這與大多數的研究一致,認為熱帶亞熱帶地區磷是生態系統生產力的限制因子[20]。Cleveland等[21]在熱帶施磷實驗的結果發現,施磷顯著提高土壤呼吸,并且大多數的研究表明酸性磷酸酶(AP)的活性與環境中磷的有效性呈負相關的關系[12]。相關研究發現,施用磷肥可以使土壤中磷的含量增高,而酸性磷酸酶的活性則降低[22]。一般來說,當微生物存在磷限制時,微生物就會分泌更多的磷酸酶促進土壤有機磷礦化,緩解自身的磷限制,微生物分泌的磷酸酶增多,土壤βG∶AP、NAG∶AP的值則降低。
由于土壤酶活性受土壤碳底物和養分的影響,且C、N、P相關的酶活性之間存在權衡關系,因此土壤生態酶化學計量比存在空間的異質性[23- 24]。本研究發現土壤AP的活性顯著高于βG、NAG的活性,并且砂巖發育的土壤AP的活性顯著低于花崗巖發育的土壤,而砂巖發育的土壤βG∶AP、NAG∶AP則顯著高于花崗巖發育的土壤,說明相對于砂巖發育的土壤,花崗巖發育的土壤微生物受磷養分限制的程度更嚴重,不同母巖發育的土壤,其磷素含量存在一定的差異[25],因此,在一定程度上母質可能是影響土壤磷有效性的關鍵因子。然而土壤微生物磷養分限制的程度除受母質類型的影響,亦與林分類型密切相關。從花崗巖發育的土壤上來看,本研究發現米櫧林βG∶AP和NAG∶AP的值都顯著高于杉木林,同時米櫧林βG和NAG的活性也顯著高于杉木林,這表明在花崗巖發育的土壤上,相對于米櫧林,杉木林土壤微生物可能存在磷限制更嚴重,在碳、磷含量差異不顯著的情況下,這可能主要是由于花崗巖發育的杉木林土壤粘粒含量較米櫧林高所導致(表2),一般認為土壤粘粒含量越高,土壤對磷吸附能力越強,土壤供磷能力則越弱。但是砂巖發育的土壤則存在相反的趨勢,βG∶AP 和NAG∶AP的值都顯著低于杉木林,說明相對于砂巖發育的杉木林土壤,米櫧林土壤微生物可能存在磷限制更嚴重,這主要是由于砂巖發育的米櫧林土壤微生物可利用的碳底物和氮養分含量顯著高于杉木林(表2)。
3.2 非生物和生物因子對生態酶化學計量比的影響
土壤非生物和生物因子通過調節微生物代謝作用,影響土壤酶活性和生態酶化學計量比。冗余分析結果表明土壤pH、總磷(TP)和微生物量碳(MBC)分別解釋土壤酶活性和生態酶化學計量比變異的56.9%、27.9%和12.3%,說明土壤非生物因子對土壤酶活性和生態酶化學計量比的影響強于生物因子,這與Peng等[2]在我國不同氣候帶溫帶草原及Bowles等[26]在美國加利福尼亞不同管理強度的農業土壤中的研究結果相一致。此外,本研究結果表明土壤pH值是影響土壤生態酶化學計量比的主要因素,并且Pearson相關分析結果發現土壤βG∶AP和NAG∶AP與pH呈顯著的負相關關系,說明pH值較大時,βG∶AP和NAG∶AP的值越小,酸性磷酸酶活性相對增高,土壤受到磷養分的限制越嚴重,這與Xu等[24]對我國不同區域森林土壤的研究結果及Sinsabaugh等[7]全球尺度的Meta分析的結果相一致,其可能原因是土壤pH通過影響土壤微生物生物量和群落組成、酶的空間構象以及酶與土壤顆粒之間的結合狀態,從而調控土壤酶活性和生態酶化學計量比[27]。
土壤酶活性及生態酶化學計量比還受土壤總磷(TP)、C∶P及N∶P的影響,相關關系分析結果表明βG∶AP和NAG∶AP分別與C∶P、N∶P呈極顯著的負相關關系(表4),并且βG∶AP與土壤總磷呈顯著正相關關系,說明土壤生態酶化學計量比亦受土壤養分化學計量比和土壤養分含量的影響,本結果與Xu等[24]和Peng等[2]的研究相一致,其可能原因是土壤酶主要來源于土壤微生物、植物根系分泌、植物降解的產物以及動物殘體[28],而微生物生物量化學計量比隨土壤和植物組織養分化學計量比的改變而改變,因此土壤生態酶計量比亦能夠受土壤養分化學計量比的影響,可以反映土壤養分限制狀況[24]。
綜上所述,土壤母質可能主要通過影響土壤pH和TP含量而調控土壤酶活性和生態酶化學計量比。一般認為對于全磷含量相差不大的酸性土壤,pH值越高,有效磷含量就越高,而土壤酸性磷酸酶活性則相對更低,但是本研究中土壤pH較高的花崗巖發育土壤磷酸酶活性顯著高于砂巖,其可能原因是來自母質風化的土壤總磷含量低于砂巖,微生物可利用磷含量相對較低。森林類型則通過調控土壤C∶P和N∶P調控土壤酶活性和生態化學計量比,其可能原因是不同林分類型的森林凋落物數量和質量的不同,導致土壤有機碳數量和質量、C∶P及N∶P不同,從而改變土壤微生物可以利用C、N、P等底物和養分的數量和質量,影響土壤酶活性和生態酶化學計量比。
4 結論
本研究發現中亞熱帶地區存在著一定的磷養分限制,花崗巖發育的土壤磷養分限制的強度較砂巖發育的土壤嚴重,并且砂巖發育的米櫧林土壤磷養分限制比杉木林嚴重,而花崗巖發育的米櫧林和杉木林土壤則表現出相反的趨勢,因而磷養分限制強度因森林類型和土壤母質的不同而不同。統計分析表明土壤母質可能主要通過影響土壤pH、土壤全磷含量,調控土壤酶活性和生態酶化學計量比,而森林類型可能主要通過改變土壤C∶P和N∶P影響土壤酶活性和生態酶化學計量比,因而本研究建議,未來森林經營及管理應考慮土壤母質和森林類型差異對區域森林土壤養分循環的影響。
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
科技知識動漫(2017年7期)2017-08-09 19:52:45
科技知識動漫(2017年5期)2017-05-11 21:34:16
科技知識動漫(2017年4期)2017-04-15 22:24:55
科技知識動漫(2017年2期)2017-02-06 20:59:46
科技知識動漫(2016年10期)2016-10-18 20:35:00
