酸棗根系結構可塑性對自然梯度干旱生境的適應機制
2018-09-28 08:45:02朱廣龍陳許兵郭小倩焦秀榮周桂生
生態學報 2018年16期
關鍵詞:結構
朱廣龍,陳許兵,郭小倩,焦秀榮,周桂生
揚州大學農業科技發展研究院,教育部農業與農產品安全國際合作聯合實驗室,揚州 225009
干旱是影響植物生長、發育和分布的主要限制因素,在干旱環境中,植物的生理代謝及結構特征必定發生改變,以適應或緩解逆境而得以生存[1]。根系是植物吸收水分和養分的主要器官,直接接觸土壤,是最先感受土壤逆境脅迫的主要部位,也是土壤逆境對植物的危害最主要的部位[2- 3]。在干旱等逆境脅迫下,根系首先感應并發出信號,同時其形態結構、生理化學、生物量等發生相應變化,進而抑制植物生長或做出應激反應[4]。在長期進化的過程中,植物能夠感應外界逆境脅迫,形成了一系列抵御不利環境的機制和策略,如植物體外部形態和細胞結構的改變,生理生化代謝途徑的適應等[2]。有研究表明根系感知與響應脅迫的能力在很大程度上依賴于其對不斷變化的土壤環境的適應[5]。根系解剖結構是根系發育水平的直接體現,與其生理功能密切相關[6],其木質化程度、輸導組織、表皮的附屬結構等都會影響植物抵抗逆境的能力[7]。然而,目前對根系解剖結構適應逆境的研究大多集中在某一解剖組織響應某一特定的脅迫,植物根系整體解剖結構對不同生境的適應機制方面的研究較少。
酸棗(Ziziphusjujubavar.spinosas)為鼠李科(Rhamnaceae)棗屬(ZizyphusMill.)植物,主要生長于向陽山坡、荒蕪丘陵和平原,從濕潤的海濱到干旱的荒漠均有分布[8]。酸棗抗旱耐貧瘠能力強,具有重要的水土保持作用,是綠化荒山的先鋒樹種[9]。目前,對酸棗的研究主要集中在生物活性物質提取[10]、藥理作用探索[11]、抗旱生理及耐旱形態特征等方面[12- 14]。研究表明酸棗地上部分器官具有典型的旱生結構,葉片表面有表皮毛分布、氣孔下陷、角質層較厚,葉片和莖的結構中晶體和分泌細胞較多,增強了其抗旱性[8, 15- 16]。然而,關于酸棗地下部分根系的解剖結構特征方面的研究較少,關于酸棗根系結構特征對不同梯度干旱生境的適應機制方面的研究尚未見報道。為此,本研究以生長于梯度干旱生境中的酸棗植株為試驗材料,探究植物根系結構對不同干旱生境的適應機理,以期了解在全球氣候變遷的背景下植物對不同生態環境的適應機質。
1 材料與方法
1.1 采樣地概況
實驗材料分別采自煙臺萊山區大莊村、石家莊贊皇縣北溝村、寧夏銀川市賀蘭山附近、新疆吐魯番市紅星農場三隊。同時采集原生境土樣,測定土壤含水量。煙臺和石家莊樣地土壤類型為棕壤,土壤質地為壤土;銀川和吐魯番樣地土壤類型為灰漠土,土壤質地為沙土。樣地從東到西(煙臺-石家莊-銀川-吐魯番)構成一個自然的梯度干旱環境(各生境土壤含水量、年降水量及濕潤系數見表1)。
1.2 采樣方法
分別選取上述樣地樹齡5年(依據芽鱗痕判斷樹齡)的酸棗植株5株,以植株基莖為中心,半徑為0.5 m的圓形區域垂直向下挖取根樣。挖至細根時帶土取出,用水沖洗掉泥土,以免破壞根系幼嫩的根尖組織。
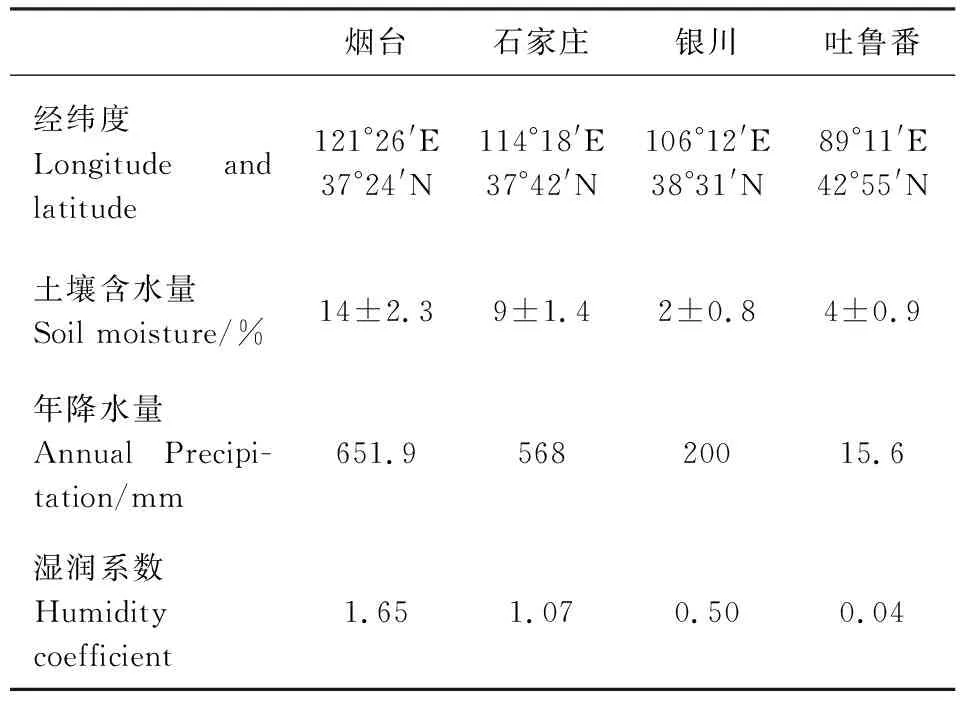
表1 采樣地區地理環境及氣候特征
濕潤系數是指一地降水量與蒸發量的對比關系,濕潤系數越小,則該地干燥程度越明顯。各個樣地的濕潤系數根據當地氣象部門提供的氣象資料計算而得。
根系初生結構的研究選取根尖處的幼嫩組織為材料,根系次生結構的研究選取植株主根地面下10 cm處的根段為材料。4個樣地中材料的選取部位和截取長度一致。將選取的材料橫切成0.5 cm長的小段,立即裝入FAA固定液中保存,帶回實驗室研究。根系結構的研究采用常規石蠟切片法。具體過程為:將采集的樣品固定24 h以上,取出經軟化劑軟化處理4—5 d(因酸棗根系次生結構硬度大,需軟化后方能切片,初生結構無需軟化;軟化劑配方:甘油10 mL+濃硝酸10 mL+蒸餾水80 mL),全自動組織脫水機(LEICA TP 1020,德國)脫水,二甲苯透明,放入50恒溫烘箱浸蠟3—5 d,然后用石蠟包埋機(LEICA EG 1150H,德國)包埋,石蠟切片機(LEICA RM 2265,德國)切片,切片厚度10 μm。將切片所得蠟帶粘片后經復水、透明,番紅-固綠雙重對染法染色(全自動染色機 LEICA AUTOSTAINER XL,德國),中性樹膠封片,自動蓋片機(LEICA CV 5030,德國) 蓋片后制成永久制片,Nikon E200 光學顯微鏡下觀察照相[15]。
1.3 統計分析
每個樣地隨機選取20 張切片拍照,FSX100 OLYMPS 觀察分析圖像,根系次生結構各參數采用 Motic Images Plus 2.0 圖像分析軟件測定,每個樣地統計50個數據,然后用SPSS 13.0統計軟件對數據進行ANOVA方差分析和最小差異法比較(LSD0.05)分析,EXCEL軟件作圖,Photoshop 軟件圖像處理,排版。
2 結果
2.1 酸棗根系初生結構
酸棗根的初生結構包括表皮、皮層和微管柱(圖1A)。表皮位于幼根的最外層,由單層體積較小、緊密排列的細胞組成,稱為表皮細胞。表皮細胞為長方形或近似球形,表皮向外突出可形成根毛。表皮的內側是皮層,占根初生結構的大部分比例,由體積較大的多層薄壁細胞組成,薄壁細胞近似圓球形,數目眾多,呈環形分布(圖1A)。微管柱位于最內層,細胞小而密集,由中柱鞘、初生木質部、初生韌皮部及薄壁細胞組成。中柱鞘是微管柱的最外層,有1—2層球形細胞組成,細胞壁薄,排列整齊而緊密,呈環形分布。中柱鞘內層為初生木質部和初生韌皮部,細胞小而緊密排列,圖中較大的細胞為正在分化的初生木質部,小細胞為初生韌皮部。初生結構發育完成后,初生木質部與初生韌皮部相間分布(圖1B)。
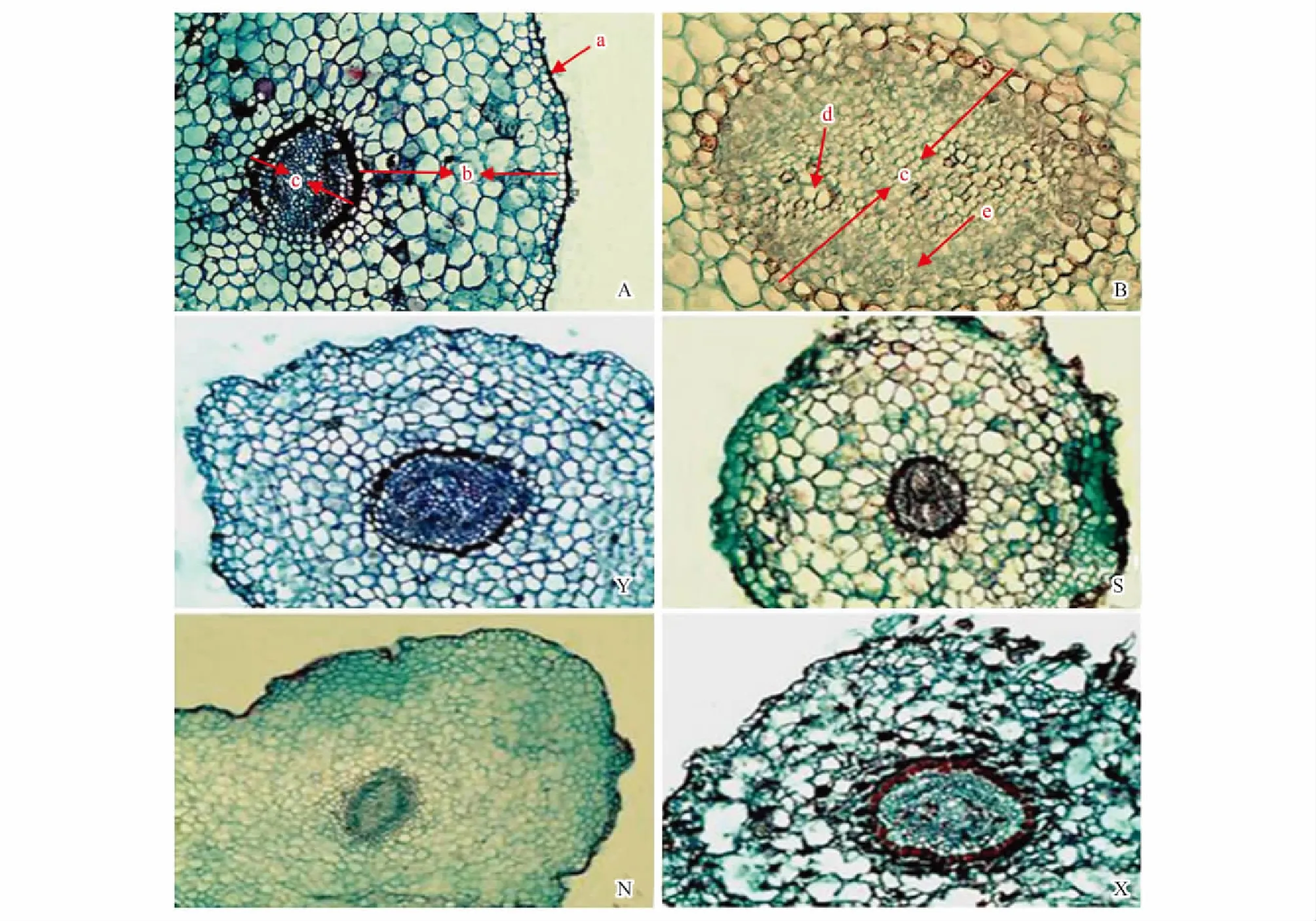
圖1 不同地區酸棗根初生結構
從煙臺至新疆吐魯番樣地,酸棗根初生結構表皮細胞的厚度和寬度逐漸增加,依次為20.65×21.12、22.12×24.06、35.13×34.34、37.40×36.44 μm,煙臺和石家莊根初生結構表皮細胞的寬度大于厚度,呈長方形橫向排列;寧夏銀川和吐魯番根系的表皮細胞厚度大于寬度,呈長方形縱向排列(圖1Y、1S,圖2)。皮層薄壁細胞的厚度和寬度變化先增后減(圖3),寧夏銀川植株根系的薄壁細胞最大,其厚度和寬度達到70.49×57.48 μm。新疆吐魯番的植株根系的薄壁細胞厚度為59.89 μm,小于寧夏銀川但高于煙臺和石家莊樣地的;煙臺植株根系的薄壁細胞厚度最小為42.95 μm(圖3)。根系初生結構的皮層厚度也隨干旱加劇而遞增,其中以寧夏銀川樣地的最高為594.13 μm,其次是新疆吐魯番402.49 μm,煙臺和石家莊的最低,分別為343.38 μm和356.08 μm(圖4)。
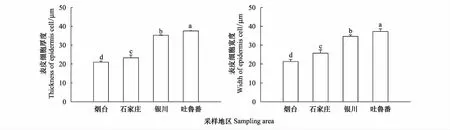
圖2 不同地區酸棗根初生結構表皮細胞厚度和寬度的變化
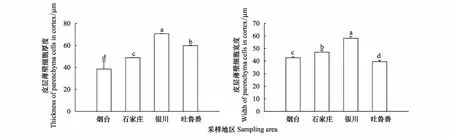
圖3 不同地區酸棗根初生結構皮層薄壁細胞厚度和寬度的變化
皮層薄壁細胞層數的變化如圖4,寧夏銀川樣地的皮層薄壁細胞層數最多為15層,其次是煙臺樣地的8層,石家莊7層,新疆吐魯番樣地的最少為6層排列。中柱鞘細胞的厚度和寬度依次呈增大的趨勢,新疆吐魯番樣地的厚度和寬度最大為26.61×27.93 μm,煙臺樣地的最小為18.01×17.86 μm,細胞近似球形排列。維管柱面積以寧夏銀川樣地的最大,是煙臺的1.20倍,石家莊的0.94倍,新疆吐魯番的1.17倍(P<0.05)(圖5)。
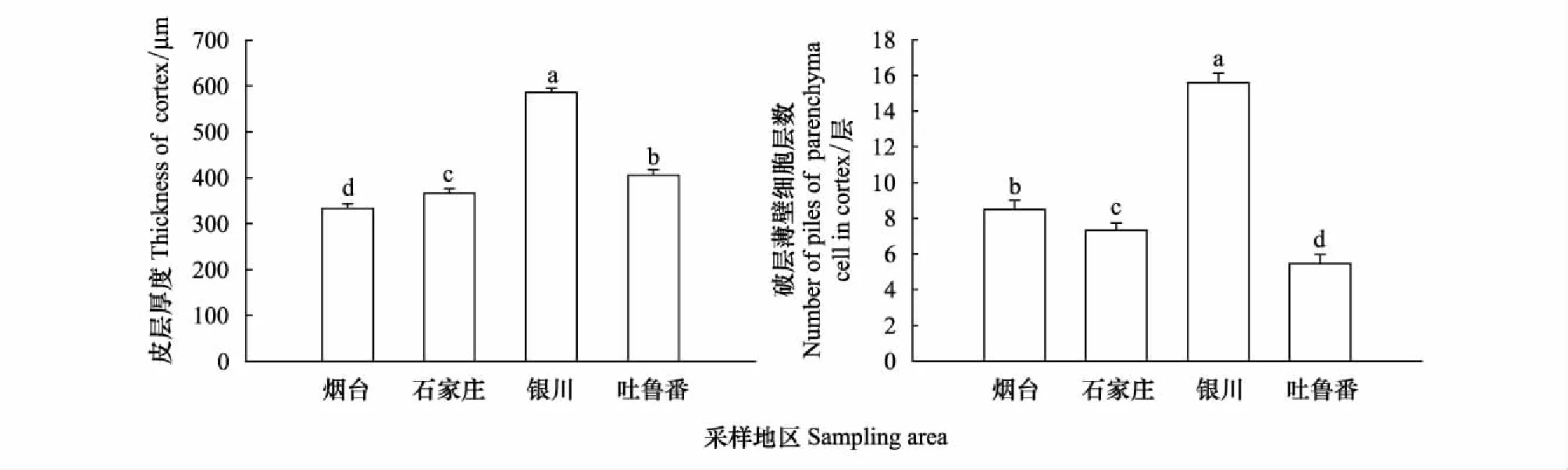
圖4 不同地區酸棗根初生結構皮層厚度和皮層薄壁細胞層數變化
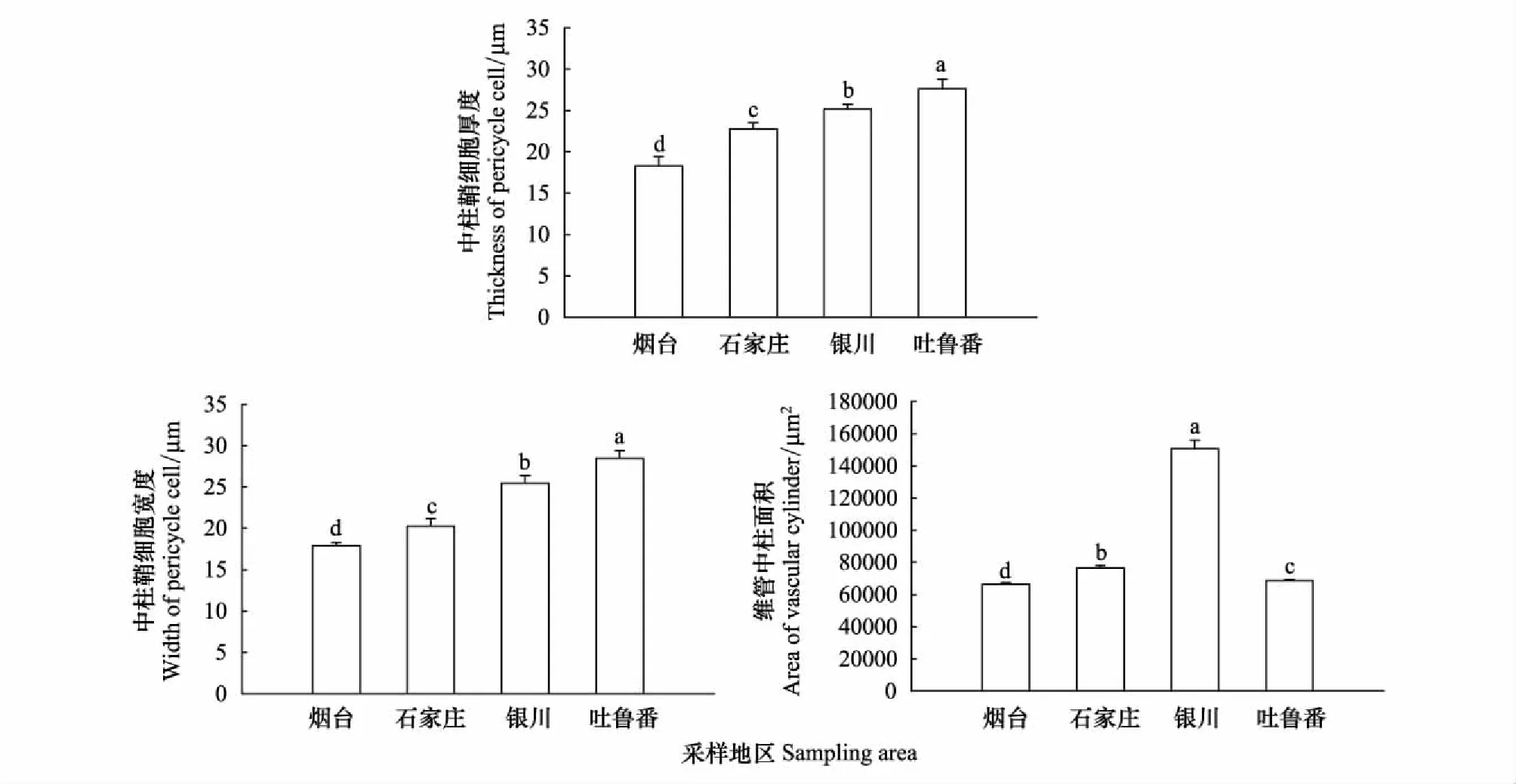
圖5 不同地區酸棗初生根中柱鞘細胞厚度和寬度及維管柱面積變化
2.2 酸棗根系次生結構
酸棗根的次生結構包括周皮(木栓層、木栓形成層、栓內層)和次生維管組織(次生韌皮部、維管形成層和次生木質部)。木栓層位于根的最外層,被染液染成紅色,細胞已老化死亡,分布松散,以后脫落(圖6A)。木栓形成層位于木栓層下方,細胞也已不同程度的木質化,細胞狹長扁平,2—3層排列緊密,呈深紫色(圖6A)。栓內層位于木栓形成層下方,細胞長方形或圓柱形,木質化程度低,著色較淺,細胞輪廓清晰,排列整齊(圖6A)。栓內層下方為次生韌皮部,細胞體積小,細胞壁薄,細胞呈小長方形或圓球形,細胞層數多,緊密整齊的堆疊在一起;包括篩管、伴胞和韌皮薄壁細胞,在次生韌皮部中,夾雜有少量的韌皮纖維,有縱向排列的韌皮射線,起著橫向運輸的作用(圖6B)。次生韌皮部下為形成層,與次生韌皮部細胞相比,細胞體積較大,3—5層緊密排列,形成層向外形成韌皮部、向內形成木質部(圖6B)。次生木質部位于形成層的下方,包括導管、管胞、木射線和木薄壁細胞。次生木質部發達,占整個次生結構的2/3。導管數量多,管徑大,管壁厚。導管分布方式多樣,有單個分布的,兩個分布的及多個導管聯合分布的。木薄壁細胞大小一致,縱向垛疊,排列整齊,各導管之間有木射線縱向發出,木射線有體積較小的長方形細胞首位相連而成;木射線與韌皮射線常是相通的,木射線和韌皮射線合成維管射線(圖6B、6C、6D)。

圖6 酸棗根次生結構 A、B、C:×10,D:×20
各生境酸棗根的次生結構如圖7所示,木栓層均已老化即將脫落,染色較深。木栓形成層和栓內層細胞也相繼老化,細胞輪廓不清晰,著色較淺(圖7Y1、7S1、7N1、7X1)。木栓層、木栓形成層、栓內層合稱周皮,能防御機械損傷,對植物其保護作用。次生韌皮部細胞數目眾多,細胞輪廓清晰,排列緊密,形成層明顯(圖7Y2、7S2、7N2、7X2)。次生木質部發達,結構清晰,輸導系統發達,導管數目眾多(圖7Y3、7S3、7N3、7X3)。各導管間有木射線發出,數目多,細胞排列整齊(圖7Y4、7S4、7N4、7X4)。從煙臺至新疆吐魯番隨生境干旱加劇,酸棗植株根系周皮逐漸加厚、致密度提高。次生木質部中,單個視野導管的數量增加,新疆吐魯番樣地的最多為101.6個,比煙臺、石家莊、寧夏銀川的分別增加了73.68%、30.93%和9.25%(P<0.05)(圖8)。隨干旱加劇,導管的直徑也依次增大。4個樣地的平均導管直徑分別為58.64、63.61、69.43 μm和72.76 μm。其中以新疆吐魯番的最大,分別高于其他地區24.08%、14.38%和4.8%(P<0.05)(圖8),說明在梯度干旱環境下,酸棗的根部通過分化出更多導管,增加管徑來提高水分利用效率,增強對干旱環境的適應性。
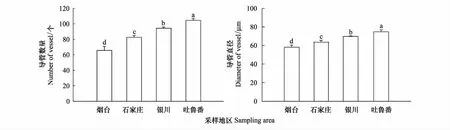
圖8 不同地區酸棗次生根中導管數量和導管直徑的變化
3 討論
本研究表明,酸棗根初生結構包括表皮、皮層和微管柱。皮層位于表皮的內部,占根初生結構的大部分比例,由大量的薄壁細胞組成,具有光合、呼吸、貯藏及各類代謝物的合成和轉化功能。從煙臺至新疆吐魯番隨梯度干旱增加,酸棗根初生結構表皮細胞的厚度和寬度逐漸增大。煙臺和石家莊根初生結構表皮細胞寬度大于厚度,呈長方形橫向排列;寧夏銀川和新疆吐魯番的表皮細胞厚度大于寬度,呈長方形縱向排列。這些表皮細胞排列方式的變化,增加了其厚度和致密性。尤其在寧夏銀川和新疆吐魯番的沙質生境中,較厚的表皮細胞有利于根的伸展。皮層細胞的厚度、寬度增大,層數增多,細胞中儲藏大量的營養物質。微管柱包括中柱鞘和初生維管組織,微管柱的面積能反應初生木質部和初生韌皮部的發育程度。在梯度干旱生境中,微管柱的面積依次增大,以寧夏銀川的最大。這與其土壤含水量有關,寧夏地區的土壤含水量是四個生境中最低的。在極度干旱的土壤環境中,植株根系以加強輸導系統的分化,提高對水分的吸收與傳輸效率。
酸棗根的次生結構包括周皮和次生維管組織,次生木質部發達,占整個次生結構的2/3。從煙臺至新疆吐魯番,次生木質部中導管數量逐漸增多,管徑依次增大。這種輸導系統的變化有利于提高輸水效率,增強抗旱性。有研究表明,水分脅迫時,達烏里胡枝子的導管比正常供水發達,其導管直徑變大對水分的輸導更有利,增強了抵抗水分的能力[17],這與本研究的結果一致。干旱條件下酸棗根部周皮和次生維管組織發達,周皮的木栓層具有保護作用,根皮層層數減少,縮短了水分吸收的距離[18];木質部發達,輸導組織所占比例有增大趨勢,能更有效地輸送水分,韌皮部的薄壁組織細胞的細胞壁強烈木質化可進一步保證輸導的安全性[19]。此外周皮的存在既能防止高溫對根的灼傷及根部向土壤反滲透失水,又可防止冬季嚴寒對根的凍傷[20]。同時可對土壤中的有害物質進行選擇隔離,阻止其進入根中,保證了植物體生理代謝的正常進行[21]。維管束是根系內主要的輸導組織,是連接根系與地上部分的通道,維管束直徑的變化可明顯反映出根系內輸導組織(導管和篩管)的發育狀況[22]。
有研究證實,諸如酸棗植株這類根部維管組織發達、韌皮部外側由數層薄壁細胞組成且胞間隙發達的結構,可使根的滲透吸水能力進一步提高;另一方面部分表皮細胞突起可形成根毛,使根的吸收面積增大[20]。木質部占有較大的比例,導管分子數量多、口徑大,根的這種結構特點有利于對水分的吸收和橫向、縱向運輸,增強了酸棗植株的抗旱能力[23]。有研究表明,在干旱加劇的條件下,小葉章根中導管的數量增加、管腔增大,維管柱厚度增加,這與本研究的結果一致,這種結構的變化有利于提高輸水效率,增強根的輸導功能,有利于適應干旱環境[24]。旱生植物狹葉柴胡的根系結構在干旱逆境中也表現出相似的變化[25],其根次生結構中周皮發達,能保護根部抵御干旱環境;導管管徑增大、數量增多有利于水分的運輸,從而提高了植株的抗旱能力[26]。
在逆境條件下,植物體內的水分和鹽分變化直接影響根系的細胞結構,并嚴重影響植物對水分和營養物質的吸收,最終表現在植物的表型上[27]。植物體內的水分狀況和水分代謝直接受土壤環境和氣候的影響。在取樣調查中,我們發現隨梯度干旱加劇,酸棗植株節間縮短、植株矮小、葉面積減小[8,15]。植物在生長過程中地上部與地下部具有協調效應,即地上部生長旺盛則地下部生長受抑,地上部生長受抑則地下部生長過快。在梯度干旱生境中,寧夏銀川和新疆吐魯番生境的酸棗植株地上部分生長減緩,而地下部根系較發達,其縱深扎根深度可達數米,以吸取土壤深層水分而維持生命。因此,在干旱逆境中,酸棗植物除了根系結構的可塑性改變外,地上部與地下部的相互協調生長也是其適應極端干旱逆境的策略之一。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
