大豆GmCYS20基因在百脈根共生 結瘤過程中的功能研究
2018-09-18 11:09:12柯丹霞彭昆鵬張孟珂賈妍
草業學報 2018年9期
柯丹霞,彭昆鵬,張孟珂,賈妍
(1.信陽師范學院生命科學學院,河南 信陽 464000;2.大別山農業生物資源保護與利用研究院,河南 信陽 464000)
半胱氨酸蛋白酶抑制劑(cystatin)在生物體內廣泛存在[1],是一類可以與半胱氨酸蛋白酶特異性結合,并抑制其蛋白水解酶活性的蛋白質[2]。Cystatin蛋白首先被報道參與調控植物發育過程,包括嚴格控制植物體內蛋白質的降解[3],參與貯藏蛋白的沉積和調運等生理過程[4-6]。此外,cystatin蛋白通過調控半胱氨酸蛋白酶的活性,從而延緩葉片衰老過程。如康乃馨(Dianthuscaryophyllus)cystatin 基因Dc-CPIn可以抑制花瓣的萎蔫[7]。水稻(Oryzasativa)cystatin 基因OC-I在煙草(Nicotianatabacum)中的異源表達延長了煙草的營養生長階段,開花時間和葉片衰老過程被推遲[8]。蘋果(Malusprunifolia)cystatin 基因MpCYS4也能夠延緩葉片的衰老過程[9]。
近年來,研究發現cystatin基因在提高植物對非生物脅迫的抗性方面具有重要作用[10-11]。水稻cystatin 基因OC-I通過調控半胱氨酸蛋白酶活性影響植株對脅迫的抗性[12]。擬南芥(Arabidopsisthaliana)AtCYS4和AtCYS5基因受高溫誘導表達,基因的過表達增強擬南芥的耐高溫能力[13-14]。蘋果MpCYS2[15]、MpCYS4[16]和MpCYS5[17]基因分別提高轉基因擬南芥對干旱、脫落酸(abscisic acid,ABA)和鹽脅迫的抗性。野生大豆(Glycinesoja) cystatin蛋白GsCPI14通過與鈣結合類受體蛋白激酶GsCBRLK相互作用,參與堿脅迫反應[18]。另外,植物cystatin蛋白對鱗翅目、鞘翅目等有害昆蟲的生長抑制作用[19-20]以及cystatin參與植物對病原菌入侵的防御過程[21-22]等方面也得到了較為深入的研究。
綜上,大量證據表明植物cystatin基因在調控植株發育,提高植物對非生物脅迫和病蟲害的抗性等方面具有重要作用。但關于該類基因在豆科植物共生結瘤過程中的功能研究報道較少。目前已經在大豆(Glycinemax)中分離并鑒定了20個cystatin基因,它們在大豆的14個不同組織中均有表達[23],其中7個基因在根瘤中具有轉錄活性[24]。Yuan等[25]細致描述了20個大豆cystatin基因的全基因組特征,并對該類基因在接種根瘤菌后不同時期的根及根瘤中的表達特征進行了檢測。其中,GmCYS20基因在接種后0.5 h的根中表達量迅速升高,在隨后的7~24 h、5、16以及21 d根中表達量持續上升,維持較高的轉錄水平。推測GmCYS20可能在大豆共生結瘤過程中發揮功能。為了明確該基因的生物學功能,本研究在前人工作基礎上克隆了大豆GmCYS20基因,在豆科模式植物百脈根中異源表達該基因,通過對復合體百脈根共生結瘤表型以及根瘤菌侵染表型的鑒定,揭示GmCYS20基因在共生結瘤過程中的功能,為進一步闡明該類基因在豆科植物共生結瘤過程中的分子機制奠定基礎。
1 材料與方法
1.1 實驗材料
大豆測序品種Williams82(W82)種子由中國科學院生態與地理研究所孔凡江研究員提供;百脈根(Lotusjaponicus)MG-20種子、改造的植物表達載體p1302G(攜帶GUS標簽)、百脈根根瘤菌PN28(攜帶LacZ標簽的MAFF303099菌株)由華中農業大學農業微生物國家重點實驗室張忠明教授提供。
1.2 GmCYS20基因的克隆和植物表達載體的構建
W82大豆種子表面滅菌后,種臍朝下平鋪于無菌潤濕濾紙上,28 ℃暗培養待萌發。收集新鮮大豆根尖組織,液氮速凍。參照RNA提取試劑盒(Invitrogen公司,USA)說明書,提取大豆根尖組織總RNA。按照TIANGEN公司反轉錄試劑盒操作說明獲得cDNA 第一鏈。根據https://www.soybase.org/search/網站公布的大豆GmCYS20基因(Glyma.20g045500)序列設計引物F-GmCYS20和R-GmCYS20(表1),PCR擴增GmCYS20目的基因。回收目的片段,連接T載體,送南京金斯瑞公司測序驗證。設計引入酶切位點EcoRI和SmaI的引物F-OX和R-OX(表1),將測序正確的目的基因插入植物表達載體p1302G(Gus基因作為篩選標記基因)中,構建p1302G-GmCYS20重組質粒,經凍融法轉入發根農桿菌LBA1334中備用。
1.3 序列分析
利用NCBI網站的Blastp工具尋找栽培大豆(Glycinemax)GmCYS20蛋白(NP_001239817.1)的同源蛋白,包括野生大豆(Glycinesoja,KHN35444.1),綠豆(Vignaradiata,XP_014516943.1),木豆(Cajanuscajan,XP_020236689.1),赤豆(Vignaangularis,XP_017411678.1),鷹嘴豆(Cicerarietinum,XP_012574834.1),蒺藜苜蓿(Medicagotruncatula,XP_003599710.2),狹葉羽扇豆(Lupinusangustifolius,XP_019419326.1),紫花苜蓿(Medicagosativa,AAZ98791.1),百脈根(AFK41117.10),栽培花生(Arachishypogaea,CBX19819.1),蔓花生(Arachisduranensis,XP_020995318.1)和落花生(Arachisipaensis,XP_016197069.1)共12種不同豆科植物的cystatin同源蛋白。利用DNAMAN軟件進行同源蛋白的多序列比對和進化樹分析。
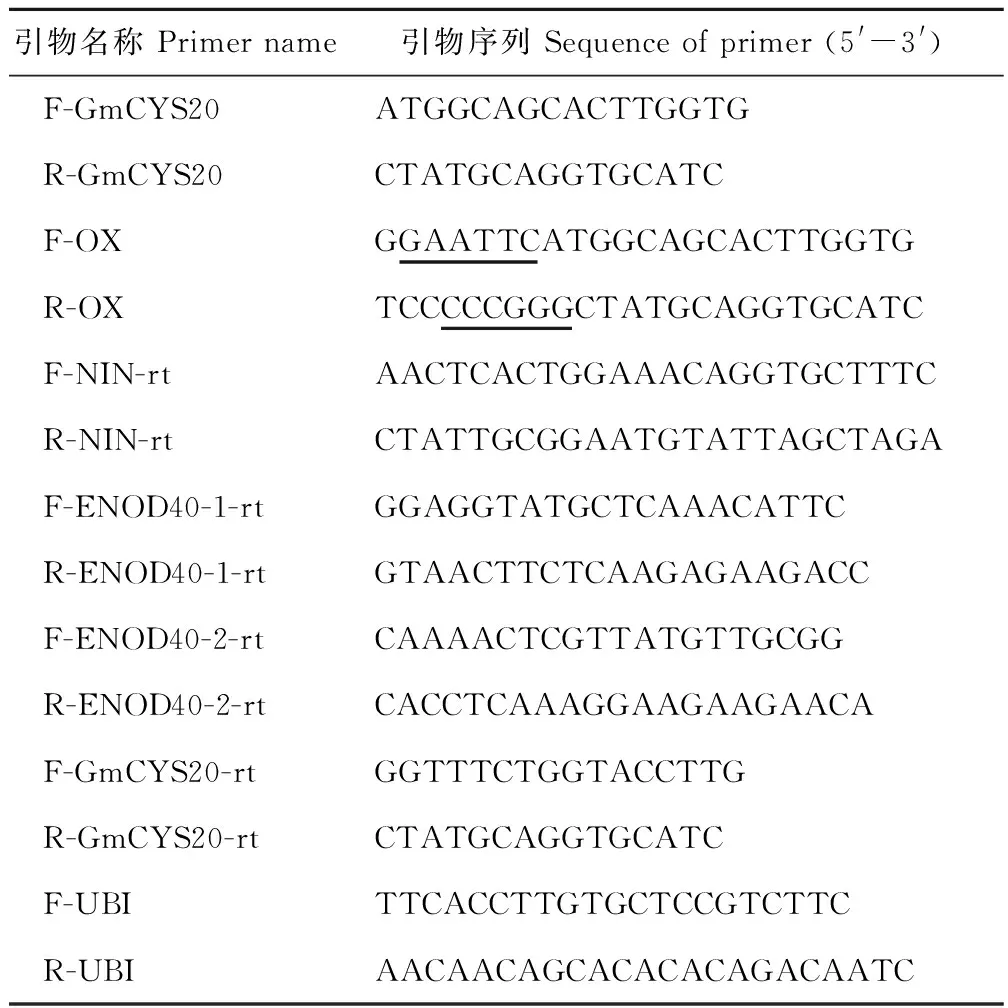
表1 本研究中所使用的引物Table 1 The primers used in this study
注:下劃線區域為酶切位點序列。
Note:The underline region indicates the restriction enzyme cutting sites.
1.4 百脈根的發狀根轉化
首先對百脈根種子進行表面滅菌,置于潤濕濾紙上22 ℃黑暗培養,待其兩片子葉展開,下胚軸長至1 cm長時,剪去根部,獲得用于侵染的外植體。用攜帶有重組質粒p1302G-GmCYS20的LBA1334農桿菌菌懸液(OD600=0.6)浸泡外植體30 min,置于MS培養基上,22 ℃暗培養3~5 d。隨后將外植體移入含300 mg·L-1頭孢霉素的MS培養基上,22 ℃培養2~3周,待發狀根長出后,標記每一條發狀根并剪下發狀根根尖部分,浸泡于GUS染液[26]中37 ℃過夜反應。觀察根尖顯色情況,與平板上的發狀根一一對應,將不變藍的陰性發狀根徹底剪掉,留下變藍的陽性發狀根繼續培養。隨機挑選部分陽性發狀根,進行RT-PCR檢測。提取陽性發狀根總RNA,反轉錄后用表1中F-GmCYS20和R-GmCYS20引物擴增GmCYS20基因,以百脈根多聚泛素(Polyubiquitin,UBI)為內參基因,引物為F-UBI和R-UBI。
1.5 復合體百脈根的結瘤試驗
將上述復合體植株種入無菌沙盆,煉苗2~3 d,第4天接種根瘤菌PN28,每天澆灌無氮營養液1次。接種4周后,分別將超表達GmCYS20復合體植株(GmCYS20-OX)和空載體對照p1302G復合體植株從沙盆中取出,用蒸餾水清洗根系,對結瘤表型進行統計拍照。根據單株結瘤數和樣本量(n),計算單株平均結瘤數,試驗重復兩次,取平均值,于2017年8月完成。
1.6 復合體百脈根的基因表達檢測
利用表1中GmCYS20和結瘤相關基因NIN、ENOD40-1和ENOD40-2的熒光定量引物檢測復合體植株陽性發狀根中各基因的轉錄水平,百脈根UBI基因作為內參。按照Takara公司PrimeScript RT reagent Kit操作說明進行熒光定量PCR(Real-time PCR)檢測,根據相對定量法(2-△△Ct:用于比較不同樣品之間的變化比率)公式計算結果。試驗重復3次,采用Excel 2007和SPSS 13.0軟件進行數據分析,并繪制圖表。
1.7 復合體植株的根瘤菌侵染表型鑒定
同1.5結瘤試驗,復合體百脈根植株接種PN28根瘤菌10 d后,取出植株,將發狀根清洗干凈,每棵植株剪下2~4條4~6 cm左右的發狀根,放入2%戊二醛溶液中固定2~3 h,使內源β-gal失活,然后用pH 7.4的PBS緩沖液清洗發狀根2~3次,去掉殘留的戊二醛溶液。最后將發狀根放入LacZ染液[27]中,30 ℃染色過夜。將染色處理后的發狀根轉移到PBS緩沖液中,制作水浸片,光學顯微鏡下觀察發狀根中根瘤菌的侵染情況。按照根瘤菌吸附到根毛頂端、侵入線進入根表皮、根瘤菌進入根瘤原基和成熟根瘤形成這4個階段統計復合體植株發狀根中根瘤菌的侵染表型。試驗于2017年9月完成。
2 結果與分析
2.1 GmCYS20基因的序列分析
根據大豆GmCYS20基因的已知序列設計引物,以大豆根尖組織cDNA為模板,PCR擴增目的條帶。經測序驗證,克隆片段長度為291 bp,堿基數及序列與網站公布序列完全一致,說明已經成功克隆了GmCYS20基因(圖1A),該基因在大豆中的ID號是Glyma.20G045500,位于大豆基因組20號染色體的8475315~8478532位,含有2個外顯子和1個內含子。序列分析表明(圖1B),GmCYS20基因的CDS區編碼97個氨基酸殘基,蛋白質的等電點(PI)為5.83,N端無信號肽,三維結構包含2個α螺旋和6個β折疊。GmCYS20屬于第Ⅲ類cystatin蛋白,含有該家族共有的4個保守結構域GG、LARFAVE、QVVSG和SW。
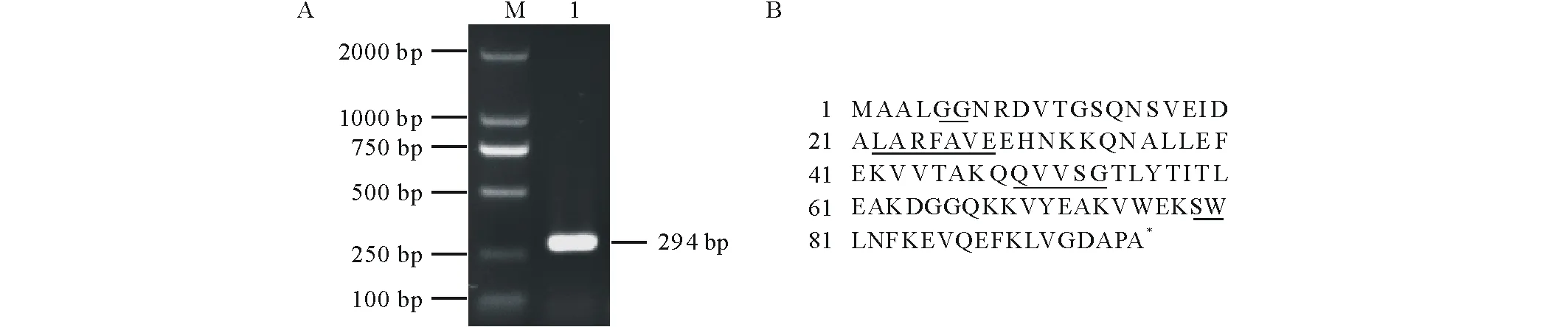
圖1 GmCYS20基因的克隆(A)和編碼氨基酸序列分析(B)Fig.1 Cloning of GmCYS20 gene (A) and analysis of encoded amino acid sequence (B) M: DL2000 DNA marker; 1: 目的片段 Target fragment; 下劃線氨基酸代表4個保守結構域GG、LARFAVE、QVVSG和SW Underlined amino acids represent 4 conserved domains GG, LARFAVE, QVVSG and SW.
在NCBI網站上對GmCYS20基因的編碼序列進行Blastp比對,尋找其他豆科植物中GmCYS20的同源蛋白。結果發現,GmCYS20與野生大豆、綠豆、木豆、赤豆、鷹嘴豆、 蒺藜苜蓿、狹葉羽扇豆、紫花苜蓿、百脈根、栽培花生、蔓花生和落花生等12種豆科植物的CYS蛋白相似性較高(圖2)。通過DNAMAN軟件獲得大豆GmCYS20蛋白與上述12種豆科植物CYS蛋白的進化樹,由圖3可知,大豆GmCYS20蛋白與野生大豆CYS蛋白處在同一進化分支上,親緣關系最近。
2.2 轉GmCYS20基因復合體百脈根的獲得
為了明確GmCYS20基因在共生結瘤過程中的生物學功能,在模式豆科植物百脈根中對GmCYS20基因進行超量表達的研究。將克隆的GmCYS20基因插入改造過的植物表達載體p1302G中,重組質粒經EcoRI和SmaI雙酶切后獲得大小正確的載體片段和目的片段(圖4A),將質粒送交公司測序。利用凍融法將測序結果正確的質粒轉入發根農桿菌LBA1334,鑒定后保存于-80 ℃,用于百脈根的遺傳轉化。
參照1.4中的百脈根發狀根轉化流程,制備轉GmCYS20基因的復合體百脈根。首先對切口處新長出的發狀根進行GUS染色。標記每一條發狀根并剪下其根尖部分,浸泡于GUS染液中37 ℃過夜反應。結果如圖4B所示,變藍的為陽性發狀根,箭頭所指為不變色的陰性發狀根。根據標記,將平板上對應的陰性發狀根徹底剪掉,留下陽性發狀根進行盆栽試驗。復合體百脈根接種根瘤菌4周后,隨機挑選部分發狀根進行GUS染色,結果發現發狀根均呈藍色(圖4C)。與此同時,隨機挑選部分發狀根,進行RT-PCR檢測。結果顯示,檢測的轉GmCYS20基因發狀根中均出現較強的轉錄信號,而空載體對照中沒有出現目的條帶(圖5),說明外源基因GmCYS20已經成功整合到GmCYS20-OX百脈根發狀根基因組中。
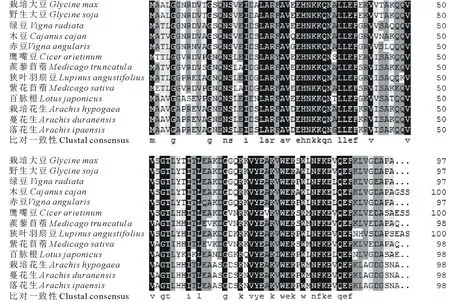
圖2 大豆GmCYS20與其他豆科植物同源蛋白的多序列比對Fig.2 Multiple sequence alignment of GmCYS20 with its homologs in other leguminous plants
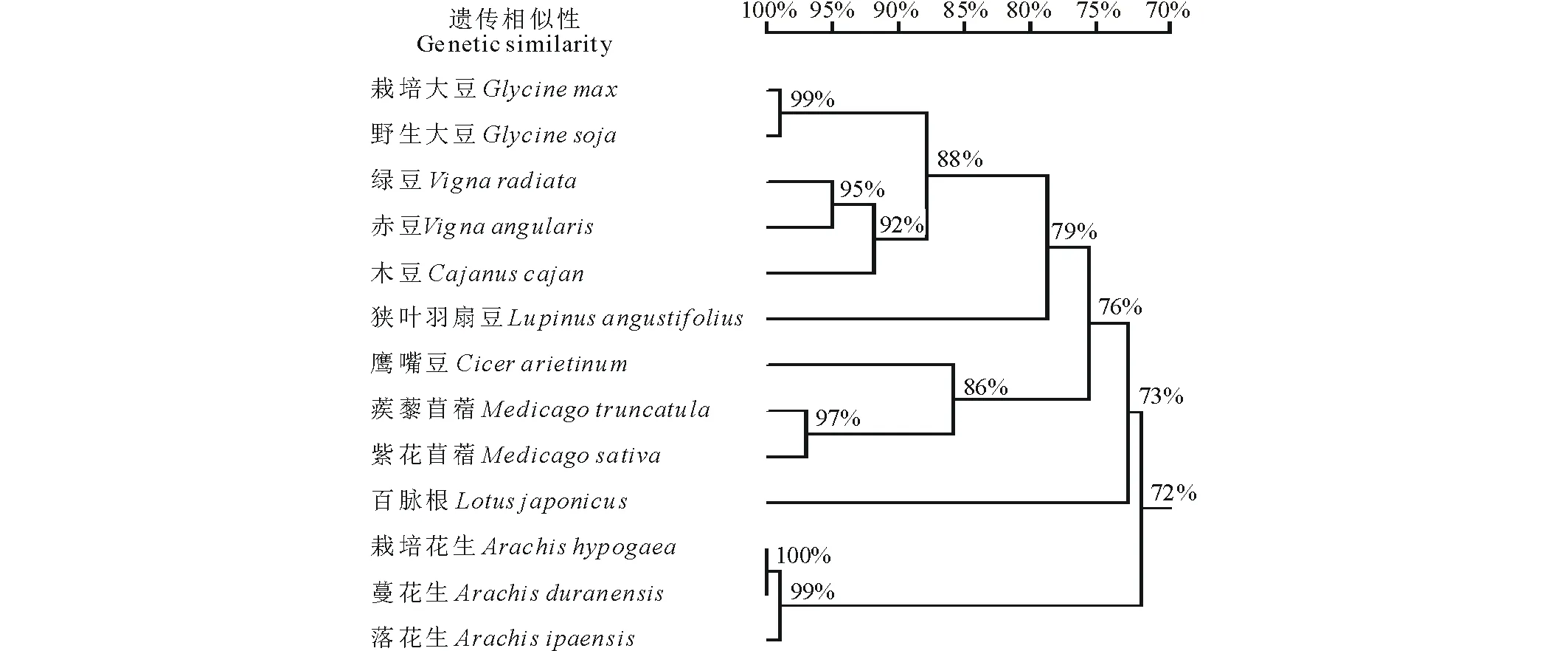
圖3 大豆GmCYS20與其他豆科植物同源蛋白的進化樹分析Fig.3 The phylogenetic tree analysis of GmCYS20 with its homologs in other Leguminous plants 標尺代表遺傳相似性,指不同植物間同源蛋白進化關系的遠近。The scale represents genetic similarity, indicates the proximity relationships among species.
2.3 復合體百脈根的結瘤表型分析
對上述鑒定為陽性的復合體百脈根植株進行結瘤試驗。接種根瘤菌4周后,統計GmCYS20-OX組和空載體對照組的結瘤表型發現,GmCYS20-OX組的單株平均結瘤數明顯高于對照組。GmCYS20-OX組的單株平均結瘤數為8.8個,而空載體對照組的單株平均結瘤數為16.1個(圖6)。說明GmCYS20基因在百脈根中的異源表達,能夠顯著增加百脈根根瘤數目,GmCYS20基因在百脈根的結瘤過程中起著積極的促進作用。
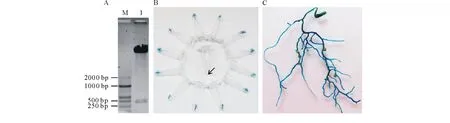
圖4 植物表達載體的構建及陽性毛根的鑒定Fig.4 Construction of plant expression vector and identification of positive hairy roots A: p1302G-GmCYS20的酶切鑒定 Identification of p1302G-GmCYS20 by enzyme digestion; M: DL2000 DNA marker; 1: p1302G-GmCYS20的EcoR I 和 Sma I 雙酶切圖p1302G-GmCYS20 digested with EcoR I and Sma I; B:毛根根尖的GUS染色圖,箭頭所示為不顯藍色的陰性毛根 GUS staining of the hairy root apex. The arrowhead is shown as a negative hairy root apex that does not show blue; C:陽性毛根接種根瘤菌4周后的GUS染色圖 GUS staining of the positive hairy roots after inoculation with rhizobium for 4 weeks.
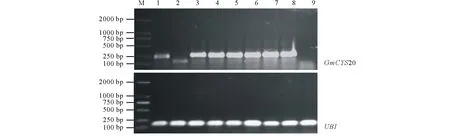
圖5 轉基因陽性毛根的分子生物學檢測Fig.5 Molecular detection of transgenic positive hairy roots 多聚泛素作為內參基因 UBI was used as reference gene. M:DL2000 DNA marker;1:陽性對照(模板為質粒) Plasmid as positive control;2:空載體對照 Empty vector control;3~8:轉基因陽性毛根 Transgenic positive hairy roots; 9:陰性對照(模板為水)ddH2O as positive control.
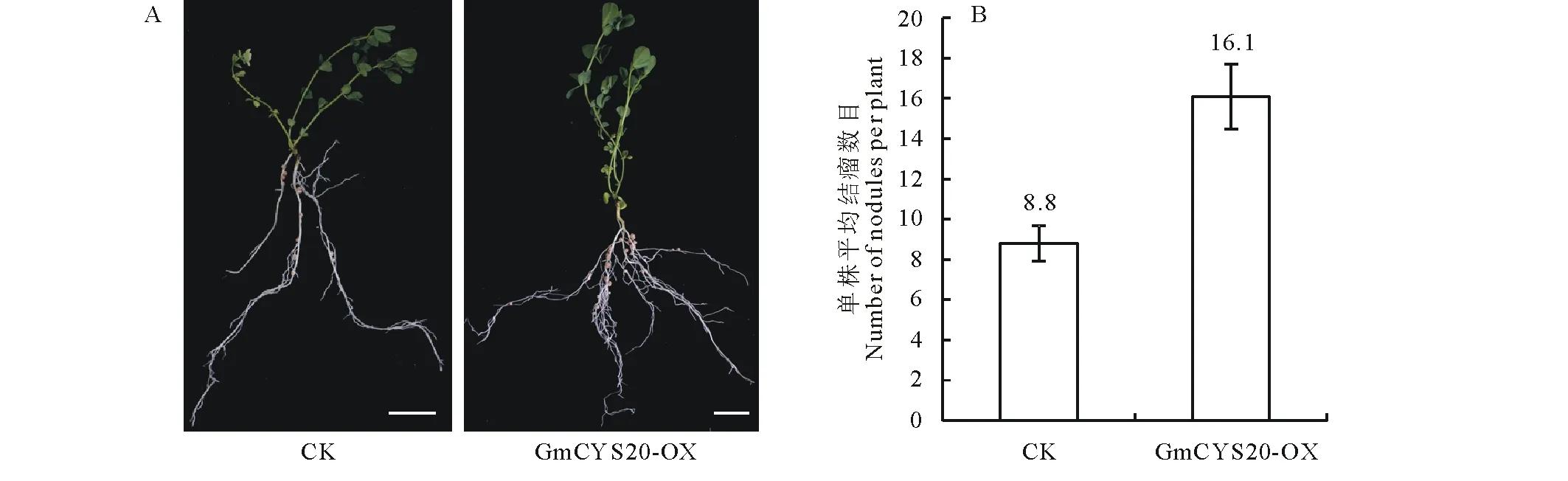
圖6 過量表達GmCYS20基因對百脈根結瘤數目的影響Fig.6 Effect of overexpression GmCYS20 gene on nodule number in L. japonicus A:接種根瘤菌4周后復合體植株的結瘤表型 Nodulation of composite plants at 4 weeks of post inoculation with MAFF303099; GmCYS20-OX:超表達GmCYS20復合體植株 Composite plants overexpressing GmCYS20; CK:空載體p1302G復合體植株Composite plants expressing empty vector p1302G; Bars=10 mm.
2.4 轉基因發狀根中GmCYS20及結瘤指示基因的轉錄水平檢測
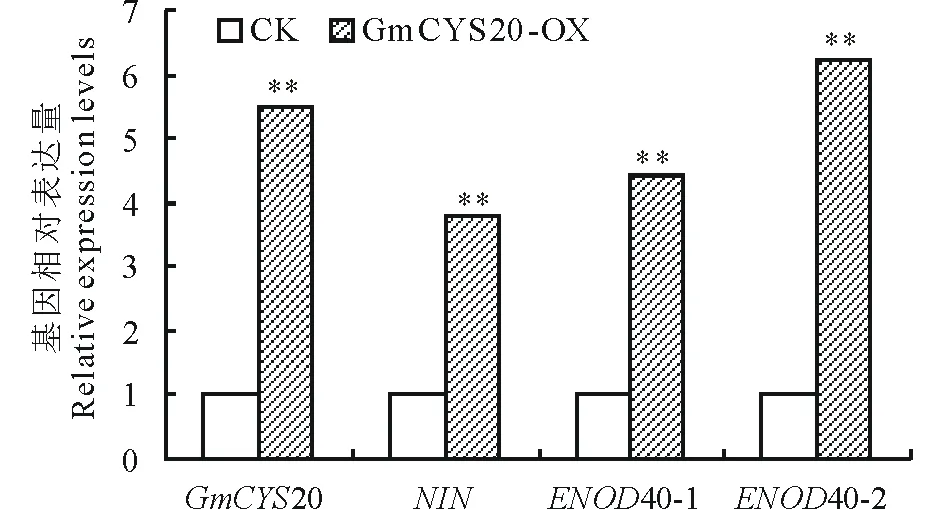
圖7 熒光定量PCR檢測復合體植株中GmCYS20及 共生相關基因的表達Fig.7 Real-time PCR analysis of the transcription levels of GmCYS20 and symbiotic related genes in composite L. japonicus NIN:結瘤起始基因Nodulation initiation gene;ENOD40-1, ENOD40-2:結瘤素基因Nodulin genes;** 表示差異極顯著(P<0.01) ** represents extremely significant difference (P<0.01).
利用Real-time PCR方法檢測轉基因發狀根中GmCYS20基因的表達,結果顯示GmCYS20基因在GmCYS20-OX發狀根中的表達水平為空載體對照(CK)的5.5倍(圖7),說明植物超表達載體的構建有效,GmCYS20基因在GmCYS20-OX發狀根中確實是過量表達的。接著檢測結瘤指示基因NIN、ENOD40-1和ENOD40-2在轉基因發狀根中的轉錄水平,進一步分析GmCYS20基因的過量表達對結瘤指示基因表達水平的影響。結瘤起始基因NIN是侵入線和根瘤原基形成的關鍵基因,結瘤素基因ENOD40-1和ENOD40-2是根瘤起始和發育的關鍵基因[28]。由圖7可見,3個結瘤指示基因在GmCYS20-OX發狀根中的表達水平均顯著增加,特別是ENOD40-2基因,為對照的6.2倍。結果表明GmCYS20基因的過量表達上調結瘤指示基因的表達。
2.5 轉基因發狀根中根瘤菌的侵染表型分析
為了進一步明確GmCYS20基因在根瘤形成的哪個階段發揮功能,對接種根瘤菌10 d的轉基因發狀根進行染色處理,光學顯微鏡下觀察發狀根中根瘤菌侵染宿主植物及根瘤的形成過程。按照根瘤菌吸附到根毛頂端、侵入線進入根表皮、根瘤菌進入根瘤原基和成熟根瘤形成這4個階段統計復合體植株發狀根中根瘤菌的侵染表型(圖8A~D)。結果表明GmCYS20-OX復合體植株在根瘤菌進入根瘤原基以及成熟根瘤形成這兩個階段的統計數目較對照明顯增多,而在根瘤菌吸附到根毛頂端和侵入線進入根表皮這兩個階段與對照組并無明顯差異(表2)。說明GmCYS20-OX復合體植株根瘤數目的增多是由根瘤原基增多導致的,與根瘤菌的侵染無關。以上數據證實GmCYS20基因與根瘤的起始和發育相關,是一個重要的共生信號調節因子。

圖8 轉GmCYS20基因毛根中根瘤菌的侵染表型分析Fig.8 Rhizobial infection assay of transgenic hairy roots overexpressing GmCYS20 轉基因毛根接種攜帶LacZ標簽的根瘤菌PN28 The transgenic hairy roots were inoculated with M. loti strain PN28 that constitutively expresses a lacZ marker.根瘤菌侵染表型分為根瘤菌吸附到根毛頂端(A),侵入線進入根表皮(B),根瘤菌進入根瘤原基(C)以及成熟根瘤的形成(D)4個類別 Rhizobial infection phenotypes included 4 categories: the position of the IT tips at the root hairs (A), at epidermis (B), at the nodule primordia (C) and at the mature nodule (D). 箭頭表示根瘤菌到達的部位,用于不同表型的統計Arrows indicate the characteristic features used for scoring the phenotypes. Bar=25 μm.

根瘤菌侵染表型The rhizobial infection phenotypeCK (n=30)GmCYS20-OX (n=30)根瘤菌吸附到根毛頂端The position of the IT tips at the root hairs4852侵入線進入根表皮The position of the IT tips at the epidermis5849根瘤菌進入根瘤原基The position of the IT tips at the nodule primordia4677成熟根瘤形成The position of the IT tips at the mature nodule4382
3 討論
百脈根是一種優良的豆科牧草,其固氮作用能夠增加土壤肥力,因此被廣泛應用于補播改良草場、建立人工放牧地和人工草地。同時,百脈根的細胞再生性能好,遺傳轉化效率高,目前已經成為實驗室研究生物固氮機理、外源基因轉化和牧草品質改良的模式豆科植物。Hansen等[29]利用發根農桿菌(Agrobacteriumrhizogenes)最早建立了“復合體植株”系統,此系統主要由非轉化的地上部分和轉化的發狀根組成。發根農桿菌介導的百脈根發狀根轉化系統,具有遺傳操作簡便、轉化效率高和穩定性好等突出優點。每個轉基因發狀根代表一個獨立的轉化事件[30]。該系統目前已經被廣泛應用于生物固氮領域基因功能的研究[31-34]。由于大豆遺傳轉化效率相對較低,研究候選基因功能費時耗力[35],因此本研究利用發根農桿菌介導的百脈根發狀根轉化系統來驗證大豆候選基因GmCYS20在共生結瘤途徑中的生物學功能,快速高效評價該候選基因在目標作物——大豆中的育種利用價值。
半胱氨酸蛋白酶數量眾多且廣泛存在于豆科植物中,幾乎參與植物生長發育的各個方面,包括萌發、晝夜節律、植物衰老和程序性細胞死亡等,在根瘤發育、脅迫調節以及根瘤衰老控制過程中也有許多半胱氨酸蛋白酶參與其中[36]。研究表明,基因沉默紫云英(Astragalussinicus)半胱氨酸蛋白酶基因AsNODF32,根瘤發育和類菌體衰老被延緩,根瘤壽命被延長[37]。抑制蒺藜苜蓿半胱氨酸蛋白酶CYP15A也可以延緩根瘤的衰老[38]。苜蓿根瘤衰老的過程中伴隨著一個保守的半胱氨酸蛋白酶亞家族MtCP1~MtCP6的表達,此亞家族蛋白可能涉及共生體結構的降解[39],這些半胱氨酸蛋白酶的表達或活性受抑制因子的調控。豌豆半胱氨酸蛋白酶PsCyp15A基因的表達在根瘤開始衰老時被激活[40]。
1983年最早報道了大豆中18個半胱氨酸蛋白酶在根瘤發育和衰老過程中維持較高的轉錄水平[41]。綜合最新的大豆共生相關數字基因表達譜[42-43]以及根瘤樣品轉錄組數據[24]發現,有28個Papain-like半胱氨酸蛋白酶可能參與結瘤、根瘤的發育及衰老,但是否存在特異的半胱氨酸蛋白酶參與調控根瘤的發育目前并不明確。半胱氨酸蛋白酶抑制蛋白是半胱氨酸蛋白酶的天然抑制劑,明確其在根瘤發育中的作用,可有效揭示其調控的半胱氨酸蛋白酶參與根瘤發育的機理。本研究中獲得的大豆cystatin蛋白GmCYS20可能也是通過調控特異的半胱氨酸蛋白酶活性,從而參與根瘤的起始和發育過程。因此,進一步點對點尋找GmCYS20蛋白對應的半胱氨酸蛋白酶底物,驗證二者之間的相互作用及酶活調節機制對于闡明cystatin蛋白在共生結瘤過程中的分子調控機理具有重要意義。
4 結論
本研究克隆了大豆的1個半胱氨酸蛋白酶抑制劑基因GmCYS20,分析發現GmCYS20蛋白是一個典型的半胱氨酸蛋白酶抑制劑。通過對轉GmCYS20基因復合體百脈根植株的共生表型鑒定發現,GmCYS20基因正調控根瘤的起始與發育。研究結果揭示了GmCYS20基因在共生結瘤過程中的生物學功能,為進一步闡明該類基因在豆科植物共生結瘤過程中的分子機制奠定了理論基礎,同時,也為揭示豆科植物與根瘤菌的共生互作機制提供了新的分子生物學證據。
