黑土區水稻生長生理特性與產量對耗水過程的響應
2018-09-17 06:53:42魏永霞楊軍明侯景翔
農業機械學報 2018年9期
魏永霞 汝 晨 吳 昱 劉 慧 楊軍明 侯景翔
(1.東北農業大學水利與土木工程學院, 哈爾濱 150030; 2.東北農業大學農業部農業水資源高效利用重點實驗室, 哈爾濱 150030; 3.黑龍江農墾勘測設計研究院, 哈爾濱 150090; 4.東北林業大學林學院, 哈爾濱 150040; 5.東北農業大學理學院, 哈爾濱 150030)
0 引言
我國是世界上最大的水稻生產國,水稻種植面積占糧食總面積的28%,耗水量占全國總用水量的54%左右,占農業總用水量的70%左右[1]。東北地區水資源在年際間、地區間以及年內分布極不均衡,嚴重制約了東北地區水稻的生產發展。因此,發展高效的節水灌溉技術對于東北黑土區農業生產應對水資源短缺及農業可持續發展具有重要意義。
節水的有效途徑就是根據水稻不同時期對水分敏感性進行針對性的水分調控,從而使得水資源得到有效利用[2]。水稻耗水過程是指在全生育期內水稻維持自身生長發育、生理活動所需水量的動態變化[3],水稻生長生理對水分的響應和適應規律一直是生物逆境學研究的重點。近年來有關研究表明,適度干旱脅迫有助于構建適宜株型結構,提高葉片活力和凈光合速率,為水分高效利用和水稻優質高產奠定了基礎。邵璽文等[4]認為,在反應敏感階段保持淺水層能夠增強后期根系活力,延緩葉片衰老時間,增加干物質積累量。大量研究表明,水分脅迫對水稻干物質積累[5-6]、根系活力[7]、光合生理特性[8-9]也存在顯著影響,良好的土壤水分條件是水稻正常生長與高產的基礎。段素梅等[6]研究則認為,在某一階段適度虧水可使復水后短期作物生長生理特性得以改善,不同生育期水稻受水分脅迫的影響機理不同,這體現了水稻對環境水分變化具有一定的適應性。以往研究中,多關于水分脅迫對水稻產量形成的影響,主要集中在單一階段、同等水分脅迫程度下的研究,缺乏科學系統的比較分析,針對精準調控條件下水稻生長生理特性對耗水過程響應的研究較少。因此,本試驗開展水稻耗水過程對水稻生長、生理特性及產量影響的研究,分析水稻生長生理特性及產量對水分條件變化的響應特征與規律,旨在探索不同階段控水帶來的正負效應及緩解這種影響的措施,有助于深入了解水稻對水分逆境的抗性機理,充分挖掘水稻生產潛能,以期為黑土區水稻高產、節水栽培提供依據。
1 材料與方法
1.1 研究區概況
試驗區位于黑龍江省慶安縣和平灌區水稻灌溉試驗站基地(北緯46°41′~47°4′,東經127°20′~127°49′),屬于北溫帶半干旱、半濕潤的大陸性氣候。多年平均降水量550 mm,多年平均溫度2.5℃,全年無霜期128 d。試區土壤種類為白漿型水稻土,土壤基礎理化指標:容重為1.02 g/cm3、孔隙度61.6%、0~30 cm體積飽和含水率平均為55.5%、pH值為6.51、有機質質量比41.5 g/kg、全氮質量比15.10 g/kg、全磷質量比15.21 g/kg、全鉀質量比20.09 g/kg、堿解氮質量比153.98 mg/kg、有效磷質量比25.21 mg/kg、速效鉀質量比157.19 mg/kg。
1.2 試驗設計
試驗在HSY.XHZ-1型自動稱重式蒸滲儀(測筒高1.5 m,直徑1.13 m,表面積1 m2,測量精度100 g)內進行,供試水稻品種為龍慶稻3號,插秧規格為行株距22.5 cm×12.5 cm,共4行24穴,每穴定5株,并配備移動式遮雨棚,防止降雨的干擾。蒸滲儀四周種植同一品種水稻作為保護田。根據作物各生育階段長度接近等原則,以水稻6個生育時期耗水量為試驗因素,采用U7(76)均勻試驗設計,在試驗范圍內挑選出具有代表性的試驗點,這些點在試驗范圍內充分均衡分散,并能夠反映體系的主要特征。60%土壤飽和含水率是水稻生長發育相對較低的水分控制標準,設置60%~100%的水分控制標準為重度調虧,依次較均勻地選取重度調虧與適宜灌水下限間的不同田面水層深度和土壤濕度,直至設置20~30 mm的田面水層深度為水分控制的最高標準,即完成6個土壤水分控制標準的選取,而對照處理(CK)是參照當地大田的水分控制標準進行設計,同時也保證了均勻設計中各時期不同處理間控水標準存在一定差異,從而可探究各因素對指標的影響及指標的變化規律。共7個處理,3次重復,21個蒸滲儀。各處理水稻各生育時期水分調控標準見表1。各處理均采用相同田間管理方法,施肥、除草等均保持一致,全生育期總施氮110 kg/hm2,基肥∶分蘗肥∶促花肥∶保花肥比例為4.5∶2∶1.5∶2;施P2O545 kg/hm2,全部用于基肥一次性施入;施K2O 80 kg/hm2,分基肥和水稻8.5葉齡時期(幼穗分化期)兩次施入,比例為1∶1。于2017年5月18日插秧,9月20日收獲。
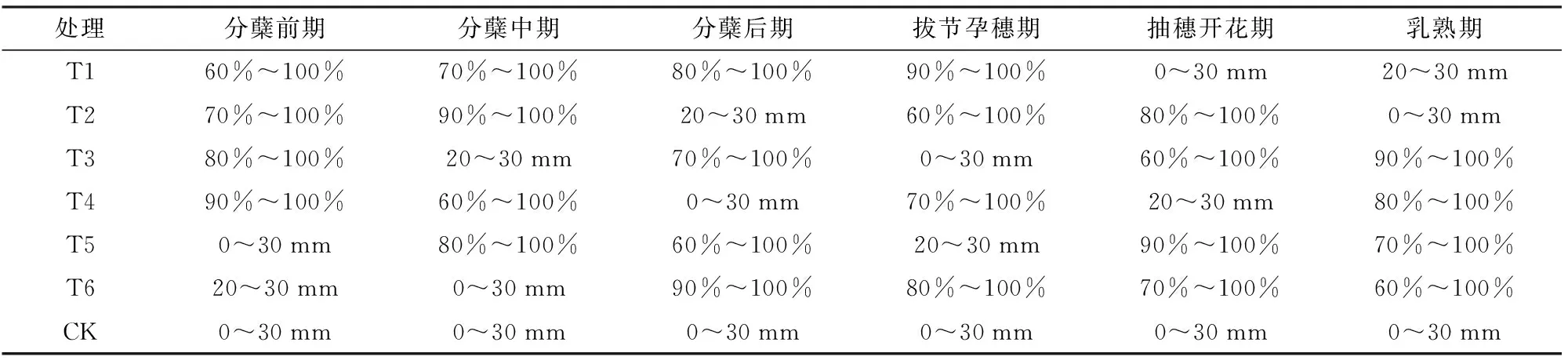
表1 水稻不同生育期水分控制標準Tab.1 Water control standards at different growth stages of rice
注:“%”為占土壤飽和含水率的百分比,“mm”為水層深度。
1.3 觀測指標與方法
1.3.1作物耗水量
每天08:00采用便攜式土壤水分速測儀觀測蒸滲儀內土表的水分變化(無水層),采用專用水尺測定水層深度的變化(存在水層);土壤水分到達控制下限即灌至設計上限,灌水前后需加測,灌水量由水表讀出。并于每天08:00使用稱量式蒸滲儀獲取總體質量,根據前后的質量差(W1)計算水稻每天實際蒸散量(耗水量),由于試驗在自動感應式遮雨棚內進行,且蒸滲儀底部有底,故不考慮生長季內降雨量、地表徑流量、地下水補給量的影響。當在各個生育時期對植株干物質積累量進行取樣時,需記錄取樣前、后蒸滲儀的總體質量,兩次質量差值(W2)為取樣引起蒸滲儀的質量變化,則W1-W2的差值即為各生育時期取樣當天的耗水量。作物階段耗水量、耗水強度、耗水模系數計算公式為
ETi=ΔWi+Ii
(1)
CRi=ETi/di
(2)
Ri=ETi/ET×100%
(3)
式中ETi——階段耗水量,mm
ΔWi——階段儲水量差值,mm
Ii——階段灌水量,mm
CRi——階段耗水強度,mm/d
di——階段持續時間,d
Ri——階段耗水模系數,%
ET——全生育期耗水總量,mm
i——生育階段,i為1、2、3、4、5、6分別代表分蘗前期、中期、后期,拔節孕穗期、抽穗開花期和乳熟期
1.3.2水稻各器官干物質積累量
分別于分蘗前期至成熟期,每處理選取代表性植株3穴,將樣品地上各部分(葉片、莖鞘、穗)擦拭表面灰塵后分別裝袋,同時進行根系取樣,挖取長等于行距、寬等于株距、深40 cm帶根土塊,用流水緩慢清洗,直至泥土沖洗干凈為止,洗凈后用無氮吸水紙吸干裝入袋中,把各時期樣品裝袋后于105℃殺青30 min,調至80℃干燥至恒質量,冷卻后用精度0.01 g電子秤測定。
1.3.3根系傷流量
分別于分蘗前期至乳熟期每個處理選取代表性植株3穴,剪去土面10 cm以上的地上部植株,套上已稱量裝有脫脂棉的自封袋密封(每蘗1袋),并用橡皮筋將其扎緊。當天18:00開始套袋,第2天8:00將其取下稱量,收集后與收集前質量差值即為單莖傷流量。
1.3.4葉片光合指標
每個蒸滲儀選取3個測試樣點進行觀測,在各生育時期對已標記的代表性單株功能葉進行測定,采用美國LI-COR公司生產的LI-6400型光合儀在每個生育期的10:00—11:00測定功能葉片凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)、胞間CO2濃度(Ci),數據由儀器自動讀出。
1.3.5測產
成熟期每個蒸滲儀選取6穴代表性水稻單打單收計算每穴實際產量,根據每公頃穴數計算每公頃產量。
1.4 數據處理方法
以分蘗前期、分蘗中期、分蘗后期、拔節孕穗期、抽穗開花期、乳熟期的耗水量為變量,采用多元回歸分析與各因變量建立水分響應模型,并對模型及各因素進行顯著性檢驗;運用Excel進行數據處理,Origin 9.10進行作圖,SPSS 22.0作回歸統計分析;多重比較采用LSD法,顯著水平取0.05。
2 結果與分析
2.1 水稻各生育時期耗水特征
表2為不同水分處理下水稻各生育時期的耗水特征。從整個生育期來看,水稻各處理生育期內耗水強度總體呈現先穩定上升后逐步下降的趨勢,6月中旬以前,日平均氣溫較低致使分蘗前期歷時稍長,植株蒸騰量較小,但水稻耗水強度仍然較高,在2.87~3.74 mm/d之間;分蘗中期水稻進入旺盛的營養生長階段,田間耗水由棵間蒸發為主轉向植株蒸騰為主,至分蘗后期平均耗水強度逐漸增加至4.56 mm/d。隨著生育進程推進和高溫天氣的持續,抽穗開花期平均耗水強度增至5.66 mm/d,此時期是水稻營養生長和生殖生長并進階段,植株生長旺盛且冠層覆蓋率較高,水稻需水量大。水稻灌漿后,各處理耗水強度開始迅速回落,至乳熟期耗水強度降低為3.56 mm/d。各處理耗水模系數在整個分蘗期最大,平均值為43.98%,抽穗期后耗水模系數基本趨于穩定,乳熟期各處理耗水模系數均值為16.02%,此時期是籽粒灌漿、產量形成的重要階段,故耗水模系數同樣較高。
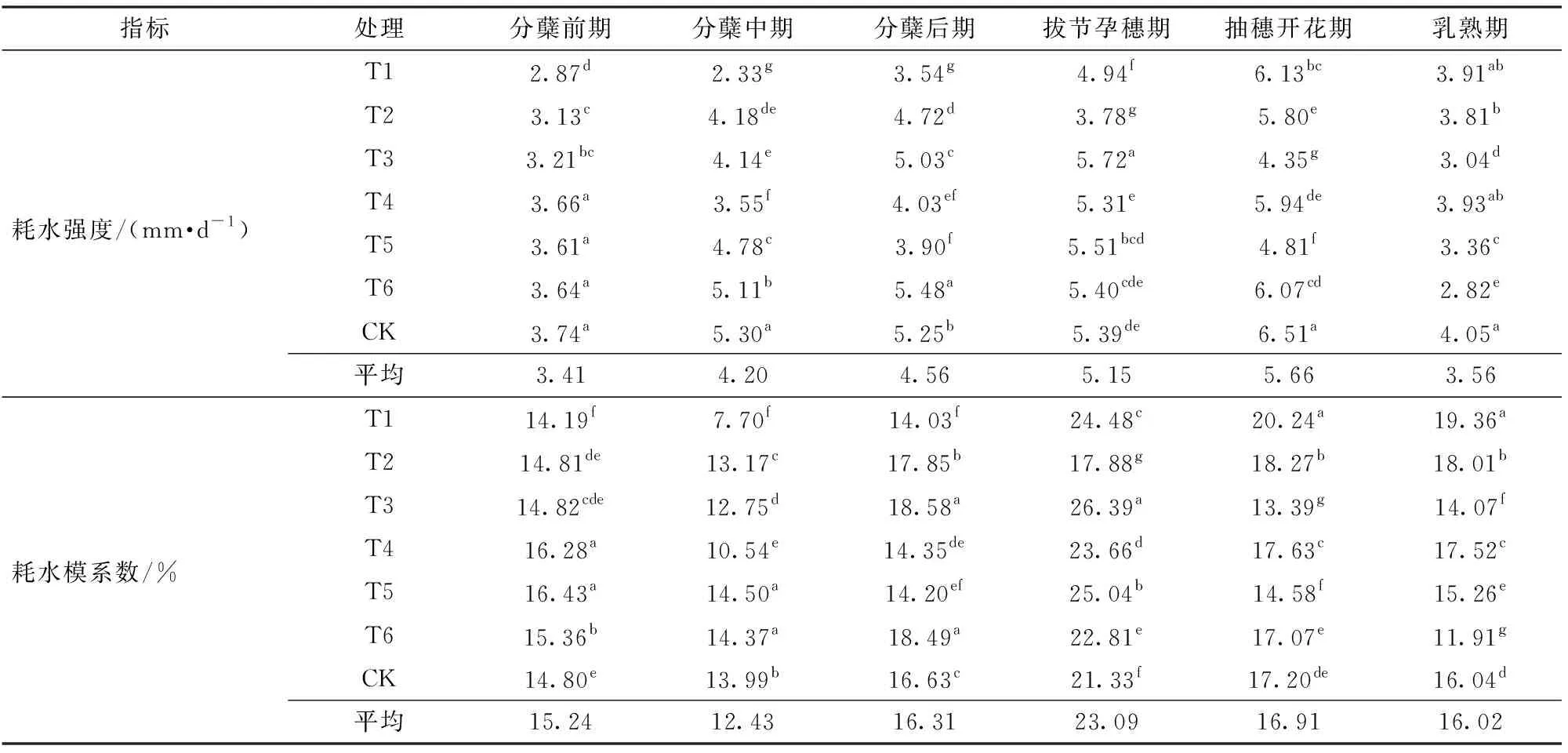
表2 各生育期耗水特征Tab.2 Water consumption characteristics of each growth stage
注:同列不同小寫字母表示在0.05水平上差異顯著。
2.2 耗水過程對水稻各器官干物質積累的影響
圖1為不同處理各生育時期水稻各器官干物質積累的變化曲線。從圖1可知,不同處理水稻各器官干物質呈現出相同器官具有相同的變化規律。水稻葉片干物質量隨著生育進程的延續均呈現單峰曲線變化趨勢,葉片干物質量在拔節孕穗期達到最大。莖鞘干物質量在抽穗開花期達到最大,此時期莖鞘干物質最大積累量可為后期光合產物向籽粒轉移提供有力保障。而水稻葉片、莖鞘干物質積累量在達到最大值后開始集中流向穗部,使穗干物質量在抽穗開花期后迅速增加。各處理根部干物質量的變化與葉片、莖鞘呈現相似的規律,且呈倒“V”型曲線,其在抽穗開花期達到最大。

圖1 不同生育期水稻各器官干物質積累動態曲線Fig.1 Dynamic curves of dry matter accumulation of rice various organs at different growth stages
由于各處理在不同生育階段控水標準不同,導致水稻各器官干物質積累過程呈現出一定差異。分蘗中期開始葉片干物質量差距逐漸顯現,至分蘗后期各處理葉片干物質量極差達到最大,為3.4 g/穴;T3、T5處理在分蘗后期至拔節孕穗期葉片干物質量增長幅度較CK處理分別提高40.3%、51.8%,可能是由于旱后復水給拔節孕穗期葉片生長帶來一定的超補償作用;T1、CK處理在拔節孕穗期后葉片干物質量降低趨勢相對平緩。分蘗中期各處理莖鞘干物質量增加幅度為全生育期最大,為5.08~7.7 g/穴。分蘗后期T2、T4處理的莖鞘干物質量增加幅度分別為5.45、6.02 g/穴,均高于CK處理;抽穗開花期進行水分調控的各處理莖鞘干物質量增加幅度均低于CK處理。拔節孕穗期穗干物質量差異不明顯,T1處理在抽穗開花期進行復水,但其穗干物質量仍然最低,為6.44 g/穴。說明抽穗開花期前的連續虧水使水稻營養器官發育受阻,進而對生殖生長階段的籽粒灌漿過程帶來不利影響。分蘗前期、分蘗中期以CK處理的根干物質量最大,分別為1.67、4.24 g/穴,分蘗后期開始,進行水分調控的個別處理根干物質量高于CK處理,T3處理的抽穗開花期根干物質量增加幅度最低,為2.17 g/穴。
水稻各器官干物質積累量是反映作物生產能力的一個重要指標,且產量的高低取決于光合物質積累量及其分配和轉化是否協調合理。水稻籽粒的灌漿物質大部分來自于抽穗期前后葉片、莖鞘、根部的貯積和抽穗后轉運到穗部的非結構性碳水化合物[10]。因此,探究不同時期耗水量對各器官干物質最大積累量的影響,對于提高各器官干物質量以將更多營養物質轉向穗部籽粒從而提高產量具有重要意義。分別建立葉片最大干物質量(ya)、莖鞘(yb)及根最大干物質量(yd)、穗最大干物質量(yc)與耗水量(分蘗前期至對應時期)的水分響應方程,如表3所示。各方程均達到顯著或極顯著水平,決定系數R2均較高,表明各方程擬合效果良好。

表3 水稻各器官干物質積累水分響應方程Tab.3 Moisture response equation of dry matter accumulation in various organs of rice
通過方程ya可知,ET1未通過顯著性檢驗(P1=0.089),其余變量達顯著或極顯著水平(P2=0.017,P3=0.002,P4=0.031)。分蘗前期作物需水量低,ET1不會對ya造成顯著影響。其中ET3系數最大,說明此時期供水對ya的影響最大。由方程yb可知,ET1、ET3未通過顯著性檢驗(P1=0.063,P3=0.067),其余變量達顯著或極顯著水平(P2=0.038,P4=0.025,P5=0.004)。其中ET2系數與ET4相差不大,說明兩個生育時期耗水量對水稻莖鞘干物質量影響程度較為接近;ET5系數最大,說明抽穗期耗水量對yb影響最大。從方程yc可知,各變量均通過顯著性檢驗,且均對yc呈顯著或極顯著正影響(P1=0.035,P2=0.004,P3=0.037,P4<0.001,P5<0.001,P6=0.029)。ET2系數為0.095,在0.01下達極顯著水平,表明分蘗中期耗水量對yc存在極顯著影響。由方程yd可知,ET1、ET5未通過顯著性檢驗(P1=0.081,P5=0.056),其余變量均達顯著水平(P2=0.034,P3=0.016,P4=0.024)。ET2系數為0.04,表明分蘗中期耗水量對yd存在顯著正影響。ET3、ET4的系數為負,表明分蘗后期、拔節孕穗期耗水量均對yd存在顯著負影響。
2.3 耗水過程對水稻根系傷流量的影響
圖2為不同處理各生育時期水稻根系傷流量的動態變化曲線。從圖2可以看出,水稻各處理根系傷流量在生育期內呈現單峰曲線的變化趨勢,根系傷流量在抽穗開花期達到最大值,乳熟期下降幅度明顯。建立抽穗開花期根系傷流量(y)與該指標達到最大值前各生育階段耗水量的水分響應方程
y=1.401+0.001ET1+0.005ET2-0.004ET3-
0.003ET4-0.003ET5(R2=0.921,P<0.01)
(4)
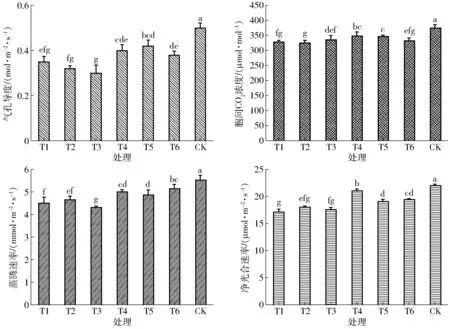
圖3 抽穗開花期各處理葉片氣孔導度、胞間CO2濃度、蒸騰速率及凈光合速率Fig.3 Leaf Gs, Ci, Tr and Pn of different treatments at heading-flowering stage
方程在0.01條件下達極顯著水平,決定系數R2較高,表明該方程能夠良好地反映根系傷流量與階段耗水量之間的關系。由方程y可知,ET1未通過顯著性檢驗(P1=0.067),ET2、ET3達極顯著水平(P2=0.005,P3=0.007),ET3、ET4達顯著水平(P4=0.018,P5=0.019),其中ET3、ET4、ET5對y為負效應。ET2系數最大,表明分蘗中期耗水量對y的調控效應最大;而ET3、ET4、ET5均對y起到一定負作用,這是因為分蘗中期根部器官已獲得良好發育,分蘗后期至抽穗期的水分脅迫雖然會導致土壤含水率低,水稻生長對惡劣環境產生的“抗性”和根系生長的趨水性促進受旱時稻根向深處延伸,特別是促進了吸收根的生長,增強根系的吸收能力[11]。抽穗開花期后水稻各處理根傷流量呈下降趨勢,這可能由于根系衰老和部分根系死亡的緣故。
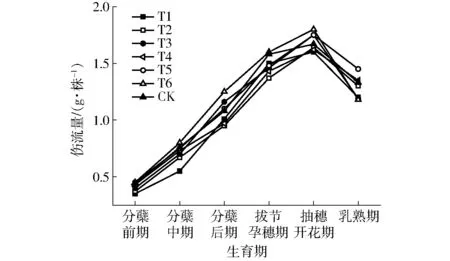
圖2 不同生育期水稻根系傷流量動態曲線Fig.2 Dynamic curves of bleeding amount of root at different growth stages
2.4 水稻葉片光合特性對耗水過程的響應及其機制分析
2.4.1耗水過程對水稻葉片光合特性指標的影響
光合作用是光合物質生產的源泉,氣孔導度、凈光合速率、蒸騰速率和胞間CO2濃度等都是表征群體光合生產能力的重要指標。不同處理水稻葉片的氣孔導度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr)及凈光合速率(Pn)均在抽穗開花期達到最大,探究耗水過程對抽穗開花期葉片光合特性的影響,對于增強抽穗期后光合生產物質的積累及更多地向穗部轉運提供重要依據[12]。圖3為抽穗開花期葉片的各光合特性指標,從圖3可知,T3處理Gs最低,為0.30 mol/(m2·s),與T1、T2處理均未達到顯著性差異(P>0.05)。T2處理Ci最低,為325.32 μmol/mol;T4、T5處理間Ci差異不顯著,但均顯著高于其他虧水處理。T6處理Tr高于T4處理,差異不顯著,但顯著低于CK處理(P<0.05);各處理與T3處理之間Tr均達到顯著性差異(P<0.05)。T1、T2、T3處理之間Pn差異不顯著,且均處于較低水平;而T4處理Pn均顯著高于其他虧水處理(P<0.05)。整體來看,CK處理的4種光合指標均顯著高于其他處理(P<0.05)。分別將抽穗開花期Pn、Gs、Ci及Tr與相應各時期耗水量建立水分響應方程,如表4所示。對各方程進行顯著性檢驗,均在P<0.01條件達極顯著水平,其決定系數R2均在0.80以上,表明各方程的擬合效果良好。

表4 各光合特性指標水分響應方程Tab.4 Moisture response equation of different photosynthetic characteristics indexes
氣孔導度反映了氣孔對外界氣體及水蒸氣的傳導程度,是與光合作用密切相關的一個指標。由方程Gs可知,ET1、ET3未通過顯著性檢驗(P1=0.075,P3=0.052),其余變量均達到顯著或極顯著水平(P2=0.037,P4=0.016,P5<0.001)。ET2相比ET1、ET3系數大,表明分蘗中期耗水量比分蘗前期、分蘗后期對Gs影響更為顯著;ET4、ET5對Gs均存在顯著正影響,但Gs對ET5的響應最為敏感。
胞間二氧化碳濃度的變化反映了葉肉細胞光合作用能力,可以作為判斷影響光合速率變化主要原因的依據。由方程Ci可知,ET1未通過顯著性檢驗(P1=0.089),其余變量均達到顯著或極顯著水平(P2<0.001,P3=0.021,P4=0.009,P5<0.001),其中ET3對Ci存在顯著負效應。ET2系數高于ET4,表明分蘗中期耗水量比拔節孕穗期對Ci影響更大;ET3系數為負值,表明分蘗后期耗水量對Ci有顯著負影響。ET5系數最大,這說明植株從分蘗前期保持良好的生長發育至抽穗開花期時,ET5才是對Ci影響最顯著的變量。
蒸騰速率不僅受外界環境條件的影響,而且還受植物本身的控制和調節,是一種復雜的生理過程。由方程Tr可知,ET3未通過顯著性檢驗(P3=0.072),其余變量均達到顯著或極顯著水平(P1=0.028,P2=0.019,P4<0.001,P5<0.001),ET1對Tr有顯著負效應。隨著生育時期推進,作物耗水量逐漸成為制約葉片蒸騰速率的主要因素,ET2、ET4、ET5對Tr均達到顯著正效應,其中ET4、ET5系數較高分別為0.061、0.073,表明拔節孕穗期、抽穗開花期耗水量對Tr的影響較其他時期更為顯著。
凈光合速率是表征植物光合作用能力的一個重要指標,同時也是物質積累和產量形成的基礎[13]。由方程Pn可知,ET1未通過顯著性檢驗(P1=0.092),其余變量對Pn的影響均達到顯著或極顯著水平(P2=0.001,P3=0.038,P4=0.009,P5=0.004)。分蘗前期水稻營養器官尚未發育成熟,葉片光合能力較弱,分蘗前期耗水量對Pn的影響不顯著。ET2系數較ET4、ET5較大,表明分蘗中期耗水量對Pn的正效應最大,也說明了分蘗中期供水充足條件下葉片良好生長對Pn的提升具有重要保證。而ET3系數為負,表明分蘗后期耗水量對Pn有顯著負效應。
2.4.2凈光合速率對氣孔導度、胞間CO2濃度及蒸騰速率的響應關系及其機理分析
抽穗開花期各處理的Gs、Ci、Tr、Pn之間的擬合效果如圖4所示,各擬合方程的R2均較高,表明方程能夠較好反映各光合特性指標之間的關系。由圖4可知,葉片氣孔導度能直接或間接對胞間CO2濃度及蒸騰速率產生影響,葉片Ci隨著Gs的增加呈二次函數關系,而當葉片Gs降低時Ci也隨之降低,而后趨于平緩。Tr與Gs呈二次函數拋物線關系,Tr隨著Gs增加而增加,但其增加幅度不斷降低;由圖4c、4d可以發現,胞間CO2濃度、蒸騰速率與凈光合速率也存在函數關系,Tr與Pn呈線性正相關關系,葉片Pn隨著Tr的增加呈直線增加趨勢;而葉片Pn隨著Ci增加呈拋物線趨勢。
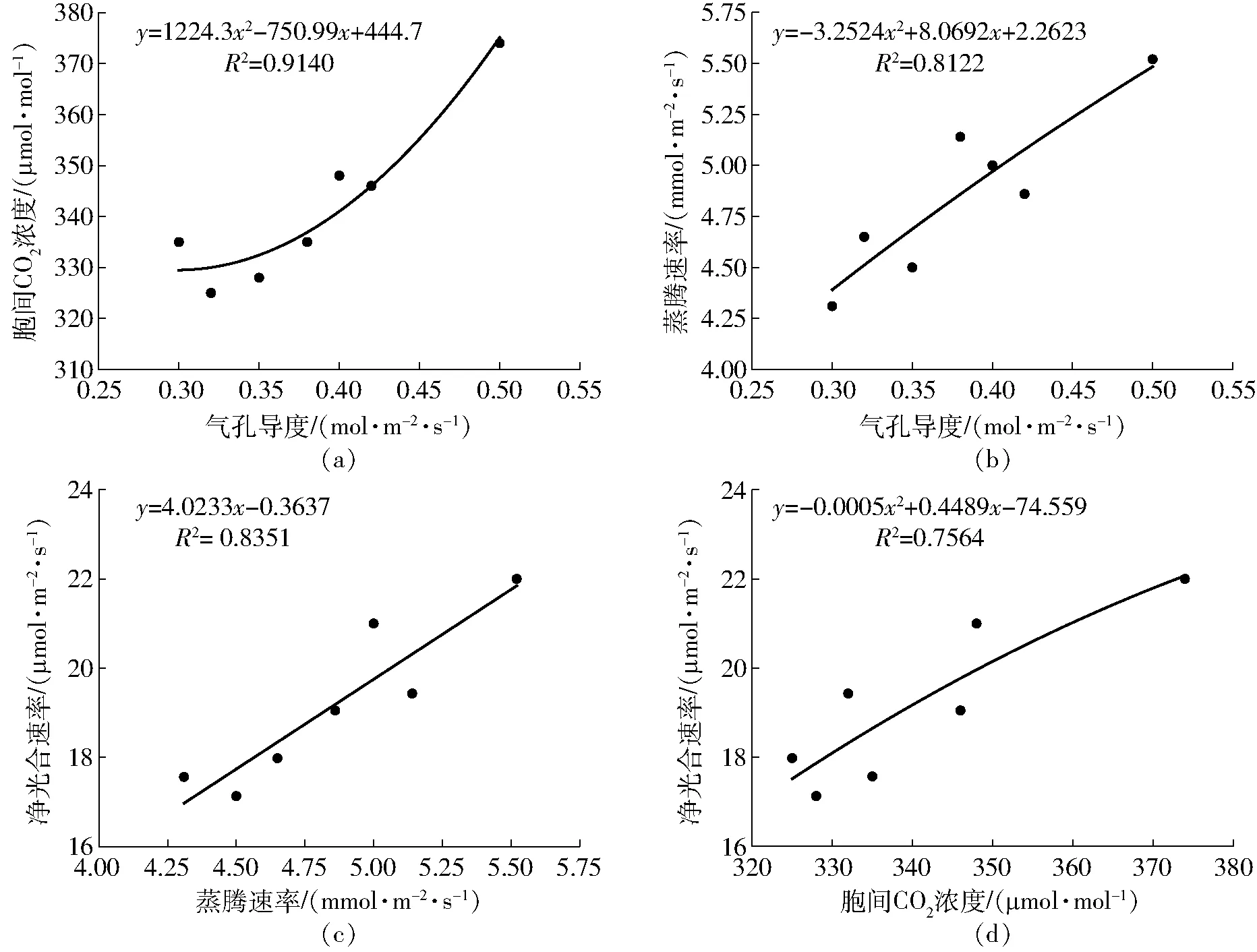
圖4 抽穗開花期光合特性指標間擬合效果Fig.4 Fitting charts of photosynthetic characteristics indexes at heading-flowering stage
分析Gs、Ci、Tr與水稻耗水過程響應關系可以進一步明確Pn對耗水過程的影響機理,結合表4和圖4分析可知,分蘗前期植株生理活動較弱,ET1對Gs影響未達顯著水平(P>0.05),不會引起外界CO2供應受阻,進而對Pn無顯著影響。ET2對Pn調控效應最大,此時期耗水量會顯著影響抽穗開花期Gs,阻礙了CO2的供應進而導致Ci減小,會引起抽穗開花期Pn降低。ET3對Pn存在一定負影響,這是由于水稻對環境CO2擴散至細胞阻力很小,這種低氣體擴散抗阻特性使得葉片在部分氣孔關閉同時,依然能保持較高胞間CO2濃度供給,保證光合作用進行[14],另外分蘗后期虧水會對生育后期復水產生一定超補償作用,一定程度引起抽穗開花期Ci增大,對Pn增加奠定良好基礎[15]。ET4增加會顯著提升Pn,結合圖4a、4d分析可知,Gs增加會促進葉片細胞吸收更多外界CO2而增加Ci,從而提高Pn。ET5增加能夠顯著提升Pn,由于抽穗開花期耗水量增加會引起葉片Gs、Tr增大的同時也利于吸收更多環境中CO2,滿足光合作用所需,保持較高胞間CO2濃度以提高葉片凈光合速率。不難發現,抽穗開花期耗水量對Pn影響最大,因為此時期處于需水關鍵期及籽粒形成關鍵階段,耗水量增加會極大提升葉片光合能力以促進光合物質生產及向穗部轉運與分配。
2.5 耗水過程對水稻產量的影響
作物水分生產函數反映了作物產量與水分之間的函數關系。圖5顯示了不同處理的水稻產量,由圖可知,CK處理的產量最高為7 168.7 kg/hm2,與其他處理間均達到顯著性差異(P<0.05),其次為T6處理,T2處理的產量最低,為5 612.2 kg/hm2,雖低于T1處理,但差異不顯著(P>0.05)。根據試驗實測的各生育時期蒸發蒸騰量,采用具有代表性的Jensen模型[16]建立水稻水分生產函數,進而得到各生育階段的水分敏感指數,Jensen模型的表達式為
(5)
式中ya——各處理實測產量
ym——充分供水條件下的最大產量
ETmi——充分供水條件下第i階段最大耗水量
方程擬合優度R2為0.984,且在0.01條件下達極顯著水平,表明模型擬合效果較好。水分敏感指數(λ)反映了作物不同生育時期因缺水而減產的程度,水分敏感指數越高,因缺水導致的減產幅度越大。對于黑土區水稻而言,各生育時期耗水量對水稻產量影響由大到小順序為:抽穗開花期、拔節孕穗期、分蘗中期、乳熟期、分蘗前期、分蘗后期。其中分蘗中期、拔節孕穗期、抽穗開花期λ值較高,分別為0.120、0.244、0.252,為水稻對水分虧缺的敏感時期,優先滿足這些時期的供水對于水稻獲得較高產量具有重要保證。
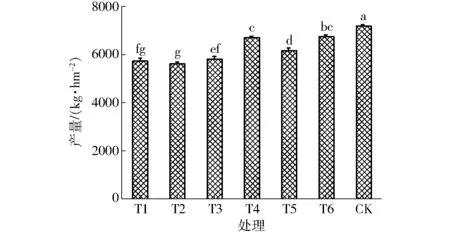
圖5 各處理水稻產量Fig.5 Rice yield of each treatment
3 討論
水稻全生育期主要階段內的耗水強度和耗水量均大致呈現出“低-高-低”的變化趨勢,在抽穗期達到最大值后逐次衰減,與石建初等[17]研究結果一致,本研究結果表明,分蘗前期耗水量及耗水強度仍然較高,其大小主要取決于插秧前土壤墑情和灌水量,因為該時段水稻幼苗葉片少且覆蓋率低,田間耗水量又以棵間土壤蒸發為主,特別是表層土壤含水率的高低密切相關。研究結果還表明,分蘗期耗水模系數占全生育期43.98%,可能是由于分蘗期階段歷時最長,地上部光合產物覆蓋率較低,土壤無益蒸發強度增大所致。不難發現,水稻的耗水強度在抽穗開花期達到最大,但有學者研究結果表明水稻在乳熟期蒸騰耗水強度最大[18],這可能是由于不同水稻品種、氣候條件及灌溉方式所引起的。
水稻干物質的構成是光合產物在不同器官中積累與分配的結果。本研究結果表明:分蘗后期耗水量對最大葉片干物質量影響大于拔節孕穗期,這是因為分蘗后期莖蘗數基本達到高峰,耗水量增加會促進葉片的快速發育,拔節孕穗期雖然處于葉片生長的關鍵階段,但其是基于分蘗后期葉片良好發育為前提條件。抽穗開花期若發生水分虧缺對莖鞘和穗干質量積累抑制作用較大,同時葉片衰老的速率加快,抑制了后期干物質的合成,導致“庫源”比例失調[19],使營養物質向籽粒的轉運與分配受到約束。而ET3、ET4對最大根干物質量均產生一定負效應,因為此時期適度的土壤水分調控,有利于改善土體環境的通透性,促進好氣性細菌的繁殖,限制根層土壤中有毒有害物質的產生,加快有機質的分解而提高肥力、促進根系發育[20],同時對稻株生理起到先抑后促,對碳氮代謝起到削氮促碳的作用,利于氮代謝旺盛逐漸轉向碳代謝,達到抑氮增糖,強化根系的作用[21],為根干物質量的積累打下重要基礎。然而過于追求水稻根部干物質積累,在一定程度也抑制葉片和莖鞘營養物質向穗部轉化,對于穗部籽粒的生長可能是一種負影響,這與吳岳軒等[22]研究相吻合。抽穗開花期水稻根部活性能夠促進土壤水分及有機物從根系轉運至地上部冠層,利于延緩葉片衰老,對于激發作物生產潛能提供必要保證[23],對最大根系傷流量研究結果表明,由于分蘗前期根系吸收水分及養分的能力較弱,耗水量對抽穗開花期根傷流量的影響并不顯著;ET2對其調控效應最大,此時期供水充足會使植株幼根的伸長生長區和根毛區發育良好,有助于改善水稻根系后期的抗逆能力進而提高根傷流量[24];分蘗后期至抽穗開花期適當水分虧缺均利于提高根系活力,進而增加根系傷流量,這與汪妮娜等[23]研究結果相似。表明水分脅迫并非完全是負效應,特定發育階段、一定水分脅迫對作物生長生理特性是有益的。本研究還發現,ET5減少對最大根干物質積累無顯著影響,但卻會對最大根傷流量提高起到一定促進作用,因為在水分虧缺的條件下,營養物質優先供應給地下部根系,因此對根系的生長和干物質積累影響不顯著[25],而此時期適當水分脅迫能促進根系對水分及營養的吸收,提高根系活力,這可能與水稻根尖細胞對水分脅迫敏感性和生長過程中離子的積累有關[26]。而尋求適宜的灌水量以平衡根干物質積累、根系活力和地上部營養器官干物質積累之間的關系,提高穗干物質積累,是今后仍需探究的問題。
水分虧缺會引起氣孔導度下降、CO2供應受阻,導致葉片光合能力降低,但嚴重虧水時葉片的光合作用會呈現不可逆性[27]。本研究結果表明,ET2增加會使水稻各項生理活動均處于旺盛水平,葉片氣孔導度增大以加快與外界氣體的交換,為作物葉片良好發育打下基礎,進而對Gs產生一定正影響;ET4、ET5對Gs均存在顯著正影響,但Gs對ET5的響應程度最為強烈,這是由于抽穗開花期為作物籽粒形成的最重要階段,為了激發植株體內碳水化合物的合成,需要提高葉片氣孔開度以增強自身的生理及代謝活動,以提高光合物質生產潛能并將光合產物及時向穗部轉運與分配[28]。研究還表明,分蘗中期遭受水分脅迫,葉片的各項生理活動將會受到極大影響,進而對葉片細胞吸收、同化環境中CO2能力造成難以恢復的影響[29],嚴重影響Ci;ET3降低均對Ci和Pn起到一定促進作用,這是由于ET3減少會使葉片氣孔導度變小來減少蒸騰損失,一方面體現了葉片為適應水分虧缺的自身調節機制,另一方面也是有效促進分蘗后期復水之后反彈補償的產生,引起氣孔增大,利于吸收環境CO2,使Ci增大并提供更多光合作用所需原料,對抽穗期Pn提升起到正向調控效果[30]。ET4、ET5對各光合特性指標均有顯著和極顯著正影響,表明此時期耗水量的增加對于葉片光合潛能發揮具有重要作用,但葉片Tr過大也會對植株產生一定的負面影響,作物在進行光合作用過程中,勢必與周圍環境發生氣體交換,因此葉片水分不可避免地要順著水勢梯度丟失,但植株蒸騰作用會產生一定蒸騰拉力,其引起的吸收過程可為植株較高部分提供水分,給作物生長帶來一定優勢[31]。因此,尋求該時期作物最佳的需水量,不僅可以減少水分消耗,而且對作物生長有利。對于葉片凈光合速率下降的原因,除了考慮氣孔導度的大小,還需考慮胞間CO2濃度的變化;由圖4a可知,隨著氣孔導度降低,胞間CO2濃度趨于平緩,這說明植物光合速率的下降,是氣孔導度和胞間CO2濃度共同作用的結果,但結果和其他學者[32]研究結論不一致,在某種情況下,認為Ci降低是葉肉細胞光合活性增大乃至光合速率增高的結果,可能是由于光合指標測定時間、氣候因素及作物品種差異造成的。
采用Jensen模型描述各處理產量對耗水過程的響應關系,結果顯示,模型中水分敏感指數在整個生育期內呈兩頭小、中間大趨勢,表明拔節孕穗期、抽穗開花期虧水對產量的影響最為顯著,這與王克全等[33]的結論一致。但本研究中拔節孕穗期和抽穗開花期的水分敏感指數較程衛國等[34]的研究結果略小,由于本研究按照生育階段接近等時段原則,將分蘗期細化為分蘗前、中、后期,生育階段劃分越細,水分敏感指數越小。水分敏感指數變化規律與水稻耗水規律相吻合,分蘗中期、拔節孕穗期、抽穗開花期耗水量對產量影響最大,分蘗中期是莖蘗生長的旺盛階段,供水充足會促進有效分蘗的發生,易形成合理的高產群體。拔節孕穗期、抽穗開花期水稻地上部冠層覆蓋率較高,營養物質逐漸從莖稈及葉片轉移至穗部籽粒中,此兩時期是產量形成的最關鍵時期,也是整個生育期的需水關鍵期,保證充足的供水對水稻產量的提升具有重要作用,同時也證明了Jensen模型的有效性和試驗結果的可靠性。
4 結論
(1)在水稻各處理生育期內,耗水強度總體呈現先穩定上升、后逐步下降的趨勢,分蘗前期各處理耗水強度在2.87~3.74 mm/d之間,至分蘗后期平均耗水強度逐漸增加至4.56 mm/d,各處理水稻耗水強度在抽穗開花期達到最大,為5.66 mm/d。水稻耗水模系數在整個分蘗期達到最大,平均值為43.98%,其次為拔節孕穗期,乳熟期耗水模系數最低,均值為16.02%。
(2)耗水過程對水稻各器官干物質積累的影響表明,各時期耗水量對葉片最大干物質量的影響由大到小順序為:ET3、ET2、ET4、ET1;對莖鞘最大干物質量的影響由大到小順序為:ET5、ET4、ET2、ET1、ET3;對穗最大干物質量的影響由大到小順序為:ET5、ET4、ET2、ET6、ET3、ET1;對根最大干物質量的影響由大到小順序為:ET3、ET2、ET4、ET5、ET1。
(3)耗水過程對水稻根系傷流量和葉片光合指標的影響表明,抽穗開花期傷流量對ET2的正響應最為顯著;ET3、ET4、ET5均對其達顯著負影響。ET2、ET4、ET5對葉片的Gs、Ci、Tr、Pn的正影響相比其他時期更為顯著;ET1對Tr存在顯著負效應;ET3均對Ci、Pn存在顯著負效應。Gs與Ci呈開口向上二次函數關系;Tr與Gs、Pn與Ci均呈二次函數拋物線關系;Tr與Pn呈線性正相關關系。
(4)耗水過程對水稻產量的影響表明,分蘗中期、拔節孕穗期、抽穗開花期λ值較高,分別為0.120、0.244、0.252,為水稻對水分虧缺的敏感階段,在黑土區有限灌溉水量條件下,水量優先分配到這3個階段,是水稻獲得高產的重要保證。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
當代陜西(2021年2期)2021-03-29 07:41:24
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
