酵母與芽孢桿菌在小麥粉基質中共培養的生長規律
2018-09-04 01:52:04黃實寬李志軍陳向東
釀酒科技 2018年8期
關鍵詞:生長
李 欣,黃實寬,常 旭,姚 娟,李志軍,陳向東
(1.武漢大學生命科學學院,湖北武漢430072;2.湖北工業大學教育部發酵工程重點實驗室,工業微生物湖北省重點實驗室,湖北省工業發酵協同創新中心,湖北武漢430068;3.湖北安琪酵母股份有限公司,湖北宜昌443003)
我國白酒發酵屬于典型的開放型自然發酵過程,多種不同微生物參與發酵過程。因此,研究微生物之間的相互作用對于白酒釀造機制的認識和釀造技術的發展均具有重要作用,特別是對白酒主體成分酒精和重要風味物質的形成發揮重要作用的酵母與其他微生物之間的互作關系[1-3]。唐潔等研究了釀酒酵母(Saccharomyces cerevisiae)和異常畢赤酵母(Pichia anomala)以順序接種混菌發酵方式的代謝特征,結果表明,混合發酵能夠形成更多的酯類物質,總酸和高級醇含量卻相對較低,有效改善了發酵液的風味特性[4]。凌杰等[5]發現醬香型白酒發酵中的釀酒酵母能夠抑制地衣芽孢桿菌的生長。吳軒德等[6]在研究釀酒酵母與巴氏醋桿菌(Acetobacter pasteurianus)混合發酵時發現酵母在發酵早期對醋酸菌的生長有抑制作用,而在發酵后期受到醋酸菌的影響而加快衰亡。醬香型白酒釀造優勢菌株拜耳接合酵母(Zygosaccharomyces bailii)與地衣芽孢桿菌共培養時受到溫度依賴型的抑制作用,即37℃下地衣芽孢桿菌抑制拜耳接合酵母生長[7]。熊小毛等[8]將東方伊薩酵母菌Z1、釀酒酵母XG-1和解淀粉芽孢桿菌WB-1混合發酵時,2種酵母菌在發酵過程中始終占優勢,而芽孢桿菌在發酵中后期明顯受到抑制。可見,不同酵母菌株之間以及酵母與細菌在白酒液態共酵過程中存在生長互作關系。
為改善和提高白酒固態發酵品質,在發酵過程中添加純種外源微生物已經成為一種廣泛采用的豐富微生物群系和維持特定微生物細胞濃度的手段[9-13]。傳統白酒釀造過程中有大量的微生物參與[14-16],這些微生物是否能用于白酒液態發酵生產工藝中,其生長適配性是關鍵。本研究以白酒釀造中常見的地衣芽孢桿菌、枯草芽孢桿菌、釀酒酵母、漢遜酵母、畢赤酵母和魯氏接合酵母為研究對象,分析共酵條件下的細菌和酵母的生長規律,為芽孢桿菌與酵母多菌種純種發酵應用提供理論和應用指導。
1 材料與方法
1.1 材料、試劑及儀器
菌株:地衣芽孢桿菌(Bacillus licheniformis)、枯草芽孢桿菌(Bacillus subtilis)。釀酒酵母(Saccharomyce cerevisiae)、漢遜酵母(Hansenula)、弗比恩畢赤酵母(Pichia fabianii)、魯氏接合酵母(Zygosaccharomyces rouxii)均由湖北宜昌安琪酵母股份有限公司提供。
小麥粉,武漢市太陽行食品有限責任公司;蛋白胨(生化試劑),北京雙旋微生物培養基制品廠;葡萄糖(食品級),山東祥瑞藥業有限公司;體積分數95%酒精(醫用級),武漢興和達商貿有限公司。
YEPD培養基:酵母粉10 g,蛋白胨20 g,葡萄糖20 g,蒸餾水1000 mL,自然pH值,115℃滅菌20 min。
LB培養基:酵母粉5 g,蛋白胨10 g,氯化鈉10 g,蒸餾水1000 mL,pH7.2~7.4,121 ℃滅菌20 min。
小麥粉培養基:5 g小麥粉中加入100 mL水,振蕩混勻,自然pH值,121℃滅菌20 min。
儀器設備:CJ-2D無菌操作臺,天津泰斯特儀器有限公司;BL-75A高壓滅菌鍋,上海博迅實業有限公司公司;HNY-211B恒溫搖床,天津歐諾儀器儀表有限公司;ZSD-A1160A恒溫培養箱,上海智城分析儀器制造有限公司;CT15RE離心機,日本日立公司;V-1100D可見分光光度計,上海美譜達儀器有限公司。
1.2 實驗方法
1.2.1 芽孢桿菌發酵液的制備
從菌種保藏斜面挑取適量芽孢桿菌菌苔于LB培養基中,37℃、180 r/min振蕩培養。待OD600達到2.5(枯草芽孢桿菌)或6.0(地衣芽孢桿菌)后結束活化。接種1%芽孢桿菌種子液至小麥粉培養基中,于30℃、180 r/min條件下發酵72 h。發酵液在10000 r/min條件下離心10 min,收集上清液,用于靜態法分析中的培養基的配制。
1.2.2 種子的制備
從新活化的YEPD平皿上挑取酵母菌落至YEPD培養基中,于30℃、180 r/min條件下培養,待OD600至達到15(釀酒酵母、漢遜酵母和畢赤酵母)或3.0(魯氏酵母)后,結束培養并作為種子液用于后續靜態法和動態法分析實驗。
從新活化的LB平皿上挑取芽孢桿菌菌落至LB培養基中,于37℃、180 r/min條件下培養,待OD600至達到1.0(枯草芽孢桿菌)或3.0(地衣芽孢桿菌)后,結束培養并作為種子液用于后續動態法分析實驗。
1.2.3 靜態法分析細菌對酵母細胞生長影響
靜態法是在酵母細胞培養的初期提供一定濃度的營養物質,而在整個細胞培養周期中不再額外添加任何營養物質。選取葡萄糖、蛋白胨和芽孢桿菌發酵液作為組分,設計了6個培養基,如表1所示。取0.5 mL酵母種子液加入2 mL無菌離心管中,以8000 r/min離心10 min,去除上清液,并在沉淀中加入0.2 mL無菌水,混勻后全部接入50 mL酵母生長培養基中(接種量1%),于30℃、180 r/min條件下培養24 h,并測定生物量。
1.2.4 動態法分析細菌對酵母細胞生長影響
動態法利用芽孢桿菌水解淀粉的能力為單一芽孢桿菌和單一酵母混合培養提供持續性碳源。取1.0 mL酵母或0.5 mL芽孢桿菌種子液加入2 mL無菌離心管中,以8000 r/min離心10 min,去除上清液,并在沉淀中加入0.2 mL無菌水,混勻后全部接入50 mL小麥粉培養基中(酵母的接種量2%,芽孢桿菌接種量1%),于30℃、180 r/min條件下培養,每24 h取樣用于測定菌數。
1.2.5 微生物菌數的檢測
微生物菌數的檢測采用經典的稀釋涂布法。新鮮的發酵樣品經過適當倍數的稀釋后涂布在固體培養基上,于30℃(酵母)或37℃(細菌)培養至菌落可見。LB固體培養基用于芽孢桿菌的計數。含有終濃度為25μg/mL氯霉素的YEPD固體培養基用于酵母的計數。總微生物數和酵母比例的計算如下:
總微生物數(CFU/mL)=酵母細胞數(CFU/mL)+芽孢桿菌菌數(CFU/mL);
酵母比例(%)=(酵母細胞數(CFU/mL))/(總微生物數(CFU/mL))×100。
1.2.6 數據分析
每組實驗3個平行。運用Excel計算平均值和方差,并運用Origin 8.6軟件對數據作圖。
2 結果與分析
2.1 芽孢桿菌發酵液組分分析
B.licheniformis離心發酵上清液中的還原糖濃度為(3.29±0.11)g/L,pH5.82±0.05。B.subtils離心發酵上清液中的還原糖濃度為(10.68±1.81)g/L,pH7.06±0.14。
為了清晰地顯示芽孢桿菌發酵液對酵母生長的影響,后續研究設置2個對照樣。由于芽孢桿菌在小麥粉發酵培養基中會水解淀粉形成一定濃度的游離葡萄糖,將只含有2%葡萄糖的培養基設置為對照1,用于判斷酵母生長缺失是否是由于營養碳源不足所導致。將含有2%葡萄糖和0.2%蛋白胨的培養基設置為對照2,用于分析是否是氮源缺乏導致酵母無法生長。2個對照樣有助于解析發酵液營養條件與酵母生長的關系。
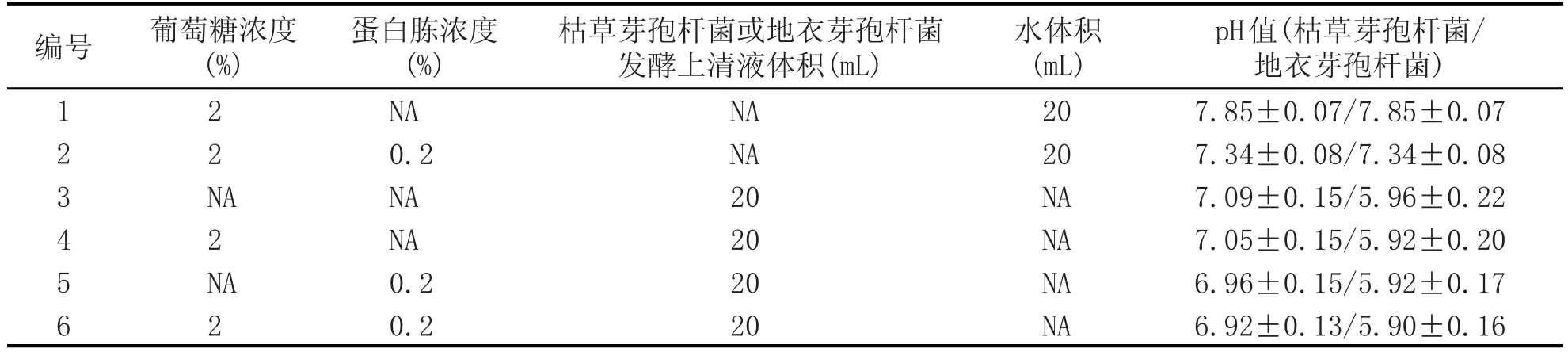
表1 6種酵母生長培養基
2.2 枯草芽孢桿菌發酵液對酵母生長的影響
圖1展示了枯草芽孢桿菌發酵液對漢遜酵母、畢赤酵母、釀酒酵母和魯氏結合酵母生長的影響。漢遜酵母在只含有葡萄糖的培養基中只有微弱的生長能力,但加入微量酵母浸粉后,其生長能力獲得極大的促進,OD600值從0.342±0.08增加到4.852±0.44。枯草芽孢桿菌發酵液中的漢遜酵母生長水平最低,僅有0.088±0.02,添加葡萄糖和酵母浸粉后,其生長能力雖有所提升,但依然低于單一葡萄糖條件。可見,漢遜酵母的生長能力受到營養不足和芽孢桿菌發酵液抑制作用的雙重影響。與漢遜酵母相似,畢赤酵母在只含有葡萄糖的培養基中也只有微弱的生長能力,但加入微量酵母浸粉后,其生長能力獲得一定的促進,OD值增大了2.96倍。然而,只要存在枯草芽孢桿菌發酵液,無論是否加入葡萄糖和酵母浸粉,畢赤酵母生長能力都沒有顯著的變化,OD600均不超過0.2。可見,枯草芽孢桿菌發酵液不足以促進其的生長。與漢遜酵母和畢赤酵母相似,釀酒酵母在只含有葡萄糖的培養基中也只有微弱的生長能力,其OD600為0.411±0.20,但加入微量酵母浸粉后,其生長能力獲得顯著的增大,其OD600為3.198±0.38。釀酒酵母在枯草芽孢桿菌發酵液中的生長能力最差,其OD600為0.232±0.06,但葡萄糖或酵母浸粉對釀酒酵母的生長能力有微弱的回補作用。然而,同時補加葡萄糖和酵母浸粉后的效果依然無法與對照2等效。可見,碳源的不足是釀酒酵母在芽孢桿菌發酵液中生長差的原因之一。枯草芽孢桿菌發酵液中還存在抑制釀酒酵母生長的因子。魯氏結合酵母在所有培養基中的生長水平都不高,且6種培養基之間沒有巨大差異的出現,OD600分布在0.15~0.28之間,因此,無法就枯草芽孢桿菌對魯氏結合酵母是否存在抑制作用得出明確的結論。這可能是魯氏結合酵母自身的營養需求特性所決定。
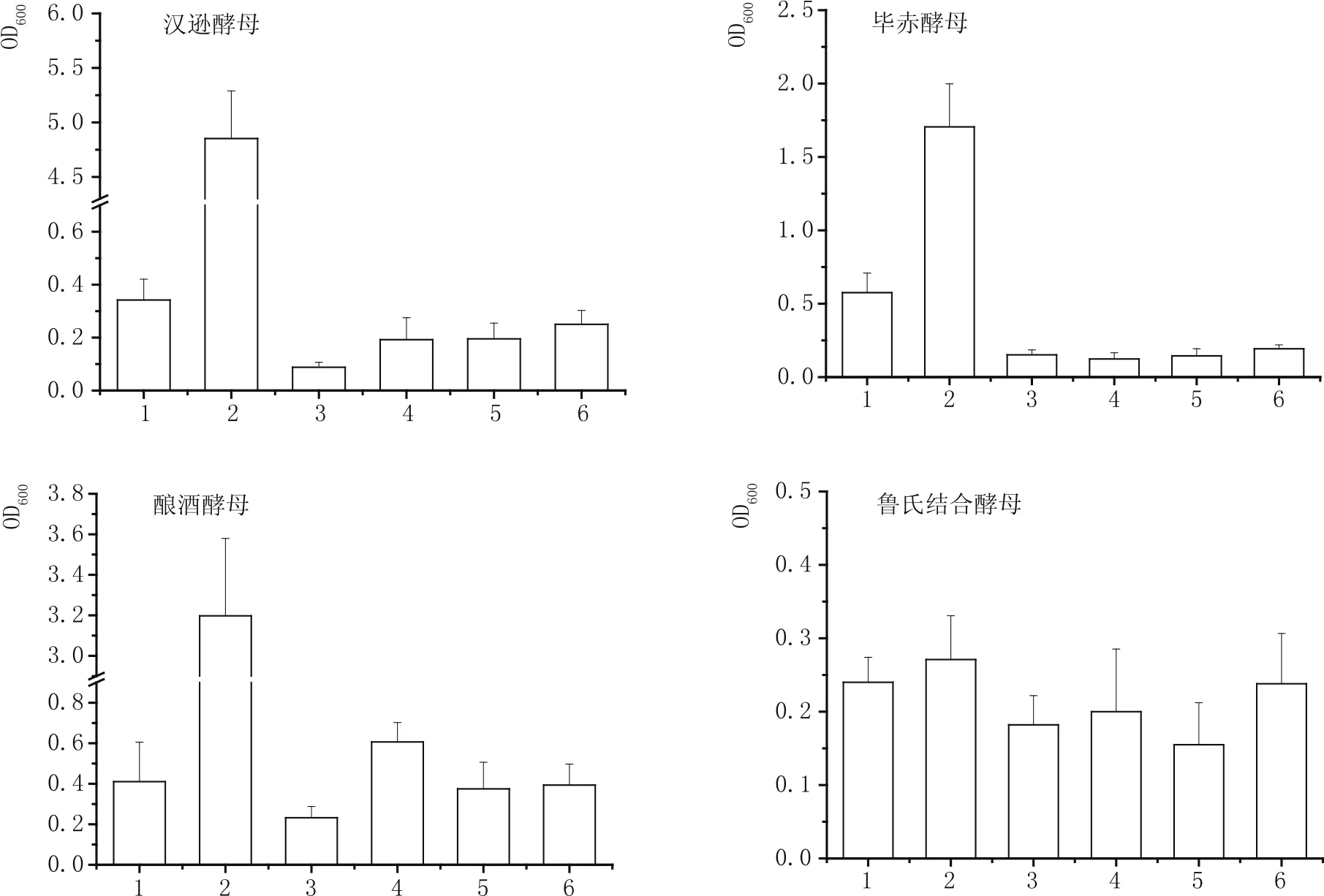
圖1 4種酵母在含枯草芽孢桿菌發酵液的培養基中的生長比較
綜上所述,枯草芽孢桿菌發酵液對4種酵母都有抑制效應,除魯氏結合酵母外。一般而言,酵母的最適生長pH值在4~5之間。是否是由于pH值過高導致酵母無法生長呢?含有枯草芽孢桿菌發酵液的培養基的pH值均在7.0附近,而含有葡萄糖和酵母浸粉的培養基的pH值在7.3左右。除魯氏結合酵母外,其余3種酵母在后一種培養基中的生長是所有條件中做好的,因此,pH值不應該被認為是酵母生長差的因素。
2.3 地衣芽孢桿菌發酵液對酵母生長的影響
圖2展示了地衣芽孢桿菌發酵液對漢遜酵母、畢赤酵母、釀酒酵母和魯氏結合酵母生長的影響。漢遜酵母在單一葡萄糖培養基中的生長是6種培養基中最弱的,其OD值只有0.34±0.08。在地衣芽孢桿菌發酵液中,漢遜酵母生長幾乎不受影響,其OD值接近3.0。無論是單一葡萄糖培養基還是地衣芽孢桿菌發酵培養基,只要加入葡萄糖,漢遜酵母的生長都有顯著的增長,分別比對照樣提高了約16倍和1.9倍。當在地衣芽孢桿菌發酵液中添加酵母浸粉時,其促酵母生長效應比添加葡萄糖的更明顯。最適培養基是在地衣芽孢桿菌發酵液中同時補加葡萄糖和酵母浸粉,其OD值為8.21±0.52。可見,地衣芽孢桿菌發酵液對漢遜酵母生長沒有抑制作用,而且添加碳源和有機氮源對漢遜酵母均有促生長效果,但有機氮源對促進漢遜酵母的生長效果更強。畢赤酵母在單一葡萄糖培養基和單一地衣芽孢桿菌發酵液培養基中具有相似的OD值,分別為0.58±0.13和0.72±0.23。當葡萄糖中加入酵母浸粉后,畢赤酵母的生長得到顯著提升,OD值超過1.5。而地衣芽孢桿菌發酵液中添加葡萄糖后,其OD值只增大了約0.2,但添加酵母浸粉后,其OD值增大到1.90±0.16。最佳的營養環境是在地衣芽孢桿菌發酵液中同時添加葡萄糖和酵母浸粉,其OD值為2.39±0.28。可見,畢赤酵母能夠在地衣芽孢桿菌發酵液中生長,其生長能力不強的主要原因是缺乏氮源。在所有的營養條件下,釀酒酵母在只存在葡萄糖的培養基中的生長能力是最弱的,OD值為0.41±0.19,但添加少量的酵母浸粉后,其生長OD值超過3.0。在地衣芽孢桿菌發酵液中的生長OD值超過1.0,而且單一添加葡萄糖并不會促進酵母生長能力的提高,但是加入酵母浸粉能極大的提升酵母生長能力,OD值為3.30±0.40。可見,釀酒酵母在地衣芽孢桿菌中具有一定的生長能力,其生長能力不強的主要原因是氮源的缺乏,而非碳源。與前述的3種酵母明顯不同,魯氏結合酵母在單一葡萄糖培養基和地衣芽孢桿菌發酵液培養基中的生長能力都很弱,其OD值均未超過0.3。同時,在這兩種基礎培養基中單一添加葡萄糖或酵母浸粉,魯氏結合酵母的生長也沒有顯著提升,OD值不超過0.4。最適的營養條件是在地衣芽孢桿菌發酵液中同時添加葡萄糖和酵母浸粉,其OD值是其他營養條件的3~4倍。可見,魯氏結合酵母的生長同時需要足量的碳源和有機氮源,且碳源比有機氮源對魯氏結合酵母更重要。綜上所述,4種酵母在地衣芽孢桿菌發酵液中生長能力不強的主要原因在于發酵液中營養物組分和濃度不足。
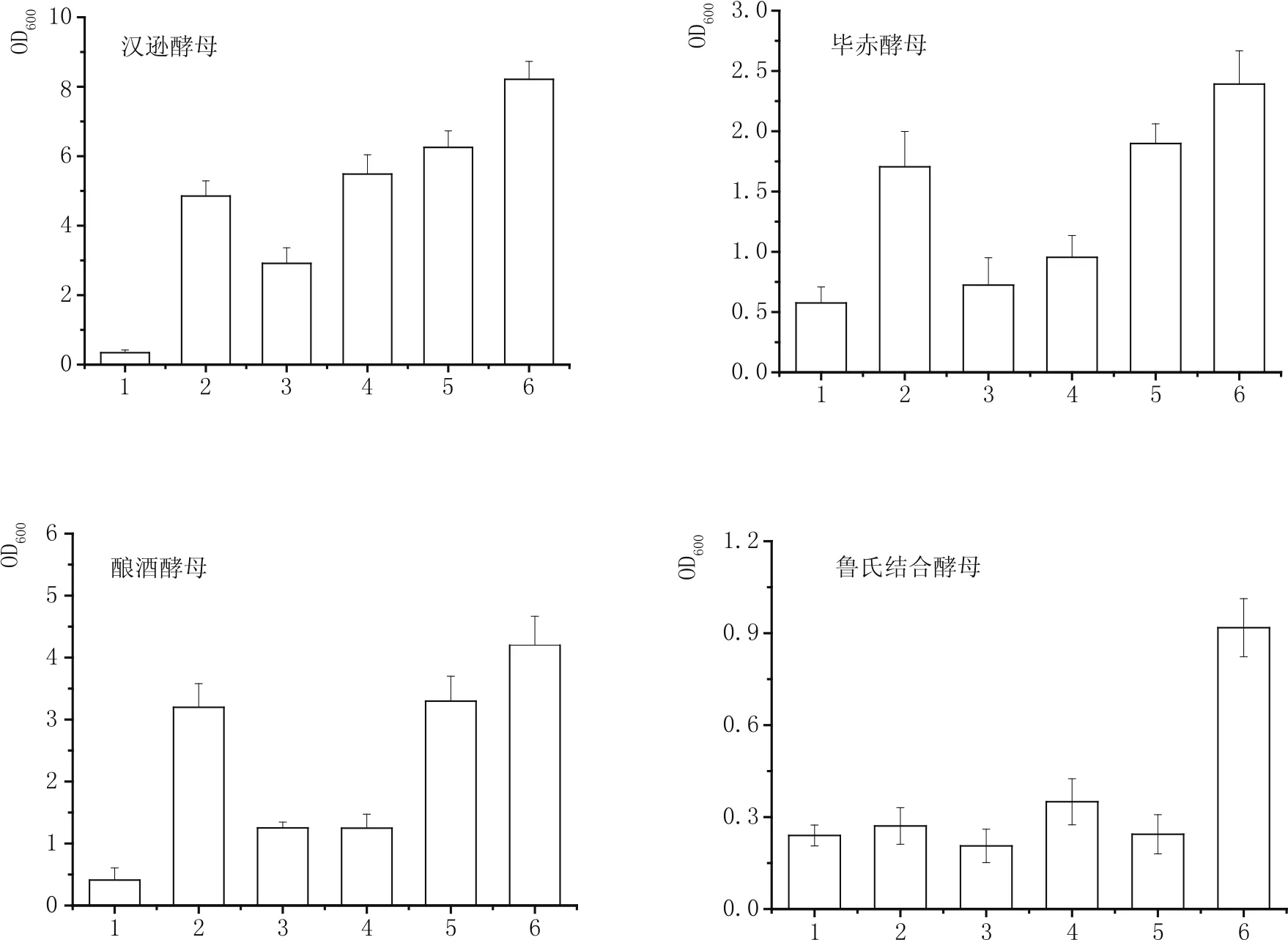
圖2 4種酵母在含地衣芽孢桿菌發酵液的培養基中的生長比較
2.4 枯草芽孢桿菌與酵母混培過程中的酵母細胞數的變化
枯草芽孢桿菌分別與4種酵母混培過程中的總菌數和酵母細胞數的變化如圖3所示。4種酵母中,漢遜酵母和釀酒酵母與枯草芽孢桿菌混培時,發酵液中的總菌數都是持續性增加,在發酵終點達到最大,分別為2.39×109±2.76×109CFU/mL(漢遜酵母)和4.58×109±0.53××109CFU/mL(釀酒酵母)。畢赤酵母和魯氏接合酵母分別與枯草芽孢桿菌混培時,總菌數分別在發酵第72 h(10.23×107±0.50×107CFU/mL)和144 h(34.20×108±0.62×108CFU/mL)積累至最大值,隨后分別下降至 2.44×107±0.13×107CFU/mL(畢赤酵母)和30.40×108±0.85×108CFU/mL(魯氏接合酵母)。除漢遜酵母和畢赤酵母外,其他2種混培條件下的酵母細胞數都呈現出持續下降過程,發酵結束時的釀酒酵母細胞數為2.33×102CFU/mL,而魯氏接合酵母檢測不到。與此對應的是,酵母比例隨著發酵進行而逐漸下降。可見,枯草芽孢桿菌分別與釀酒酵母和魯氏酵母混培過程中總菌數的變化主要反映的是枯草芽孢桿菌的菌數變化,也說明釀酒酵母和魯氏接合酵母與枯草芽孢桿菌的適配性差。在混培體系中,漢遜酵母和畢赤酵母都能保持一段生長增殖階段,最大酵母細胞數分別為5.30×108±0.54×108CFU/mL(漢遜酵母,72 h)和6.73×107±0.70×107CFU/mL(畢赤酵母,144 h),但酵母細胞數都無法維持。同時,漢遜酵母的細胞比例持續下降,只有畢赤酵母的細胞比例在發酵中后期上升并維持。
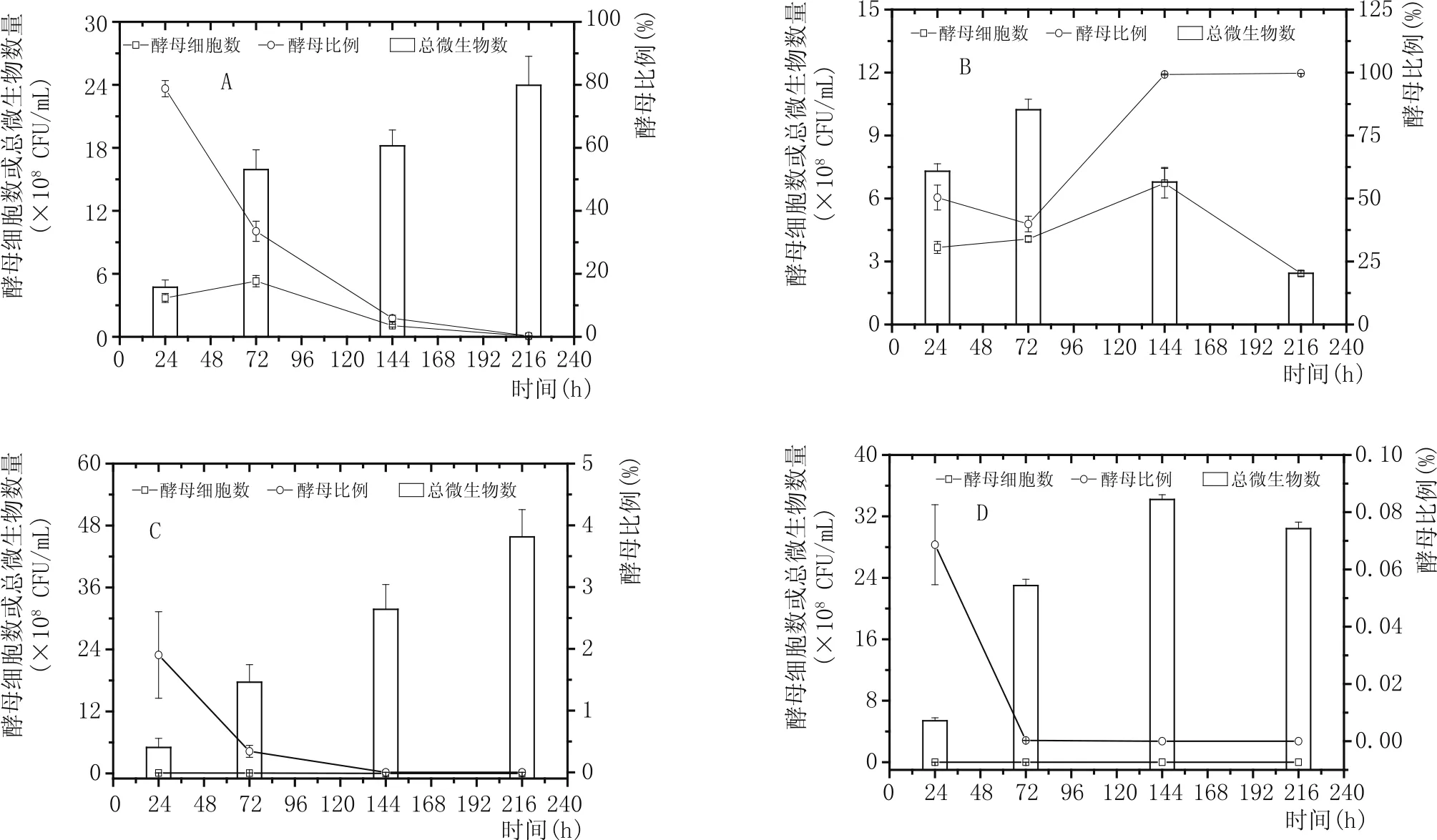
圖3 4種酵母與枯草芽孢桿菌混培過程中酵母細胞數的變化
2.5 地衣芽孢桿菌與酵母混培過程中的酵母細胞數的變化
地衣芽孢桿菌分別與4種酵母混培過程中的總菌數和酵母細胞數的變化如圖4所示。當地衣芽孢桿菌與酵母混培時,最高總微生物數均在發酵中期,分別為17.12×108±2.84×108CFU/mL(漢遜酵母,圖4A)、8.33×108±0.26×108CFU/mL(畢赤酵母,圖4B)、21.36×108±4.58××108CFU/mL(釀酒酵母,圖4C)和31.27×108±1.68×108CFU/mL(魯氏接合酵母,圖4D)。在混培體系中,除漢遜酵母有明顯的生長增殖過程(最大值為14.70×108±3.00×108CFU/mL),畢赤酵母和釀酒酵母的細胞數分別在0.22×108CFU/mL和0.70×108CFU/mL左右波動,而魯氏接合酵母細胞數呈持續下降,發酵終點的細胞數只有3.67×102±0.47×102CFU/mL。相對應的,漢遜酵母細胞比例持續增大,而另外3種酵母的細胞比例持續下降。可見,這4種混酵體系中,除漢遜酵母與地衣芽孢桿菌的混酵過程以酵母細胞增殖為主,其他3種情況皆以地衣芽孢桿菌增殖為主。同時,漢遜酵母與地衣芽孢桿菌的適配性在這4種酵母中是最佳的。
3 結論與討論
相比單菌種純種發酵,多菌種純種發酵有著更復雜的互作關系。不同種屬和性狀的微生物在生長和代謝之間的影響將對發酵產物產生巨大的影響。本研究采用的靜態法(恒定的初始碳源)和動態法(持續供給碳源)相互補充,共同揭示了2種芽孢桿菌和4種酵母在生長上的相互影響。營養物濃度對酵母的生長繁殖能力起主要作用。同時,枯草芽孢桿菌對酵母有生長抑制效應,這種作用來自枯草芽孢桿菌合成的某種或某些代謝產物,但地衣芽孢桿菌對酵母的生長抑制作用微弱。與此相匹配,當有持續性的碳源提供時,漢遜酵母顯著生長,而畢赤酵母或明顯增殖(枯草芽孢桿菌中)或維持生長(地衣芽孢桿菌中),釀酒酵母或持續減少(枯草芽孢桿菌中)或維持生長(地衣芽孢桿菌中)。除漢遜酵母與地衣芽孢桿菌的混酵過程以酵母細胞增殖為主,其他培養過程均以芽孢桿菌增殖為主。同時,酵母與芽孢桿菌的適配性順序為漢遜酵母>畢赤酵母>釀酒酵母>魯氏酵母。
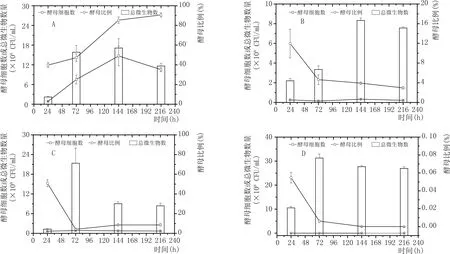
圖4 4種酵母與枯草芽孢桿菌混培過程中菌數的變化
魯氏酵母是4種酵母中受影響最大的。在恒定的初始碳源條件下,魯氏酵母幾乎不生長;持續供給碳源時,魯氏酵母細胞數在兩種芽孢桿菌中均持續性下降。這既與魯氏酵母對營養需求的特殊性相聯系,也與芽孢桿菌抑制效應有關。畢赤酵母也表現出一些有趣的現象。當枯草芽孢桿菌持續提供碳源時,畢赤酵母細胞比例持續升高,甚至出現抑制枯草芽孢桿菌的效應,而在靜態法中,畢赤酵母幾乎不生長;當地衣芽孢桿菌持續提供碳源時,畢赤酵母細胞比例下降,而在靜態法中,畢赤酵母生長能力隨營養豐度增加而提高。前者很可能與碳源不足有關,而后者可能是由于地衣芽孢桿菌在混培特定階段產生抑制性代謝產物而引起。可見,多菌種純種混培過程的微生物間生長關系復雜,它們之間的關系受到微生物菌種特性、營養環境和代謝產物等共同影響。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
