鄱陽湖濕地灰化苔草種群生產力特征及其水文響應
2018-08-29 11:08:04周云凱白秀玲寧立新
生態學報 2018年14期
周云凱,白秀玲 ,*,寧立新
1 河南大學環境與規劃學院,開封 475004 2 北京師范大學地理科學學部,北京 100875
植物是濕地生態系統中重要的組成部分之一,也是主要的初級生產者,其空間分布及生長發育受氣候、水文、土壤以及地形等多因素影響[1- 2]。大量研究表明,濕地植物往往沿某個環境梯度(如水分、鹽分、高程、溫度等)呈較為明顯的帶狀或環帶狀分布[3- 6]。對于淡水湖泊濕地而言,水分條件是影響濕地植物的關鍵因素[7- 9],濕地水分條件(淹沒深度、淹沒頻率等)從湖岸到湖心方向隨高程降低表現出一定的帶狀變化特點,形成不同植物的適生環境,從而引起植物的空間分布也呈現出相應的帶狀分異,這是濕地植物對水分條件差異性適應和響應的結果[10-12],每種濕地植物都占據著一定的水分生態位[13]。而在同一植物分布帶中,受水分梯度和水位波動的影響,植物的生長發育過程不盡相同,植物會對不同的水分條件產生趨異反應,表現出不同的生態和生理特征[14-17]。
長期以來,濕地植物與水之間的作用關系一直都是濕地生態學研究的重要領域。其中,濕地水分條件如何影響植物生長與生存、濕地水情變化如何影響植物的空間分布與群落演替又是濕地生態學研究的熱點問題[18- 21]。針對濕地植物生長發育與水分條件之間關系,許多學者已開展較為廣泛的研究。如王麗等[17]研究了三江平原小葉章(Calamagrostisangustifolia)生物量積累及其分配對不同水分梯度的響應;欒金花等[22]研究了三江平原不同水分梯度下漂筏苔草(Carexpseudocuraica)株高的生長變化規律;李亞芳等[23]則研究了洞庭湖濕地不同高程短尖苔草(Carexbrevicuspis)生長繁殖策略與水位變化之間的關系。然而當前相關研究更多側重于場地模擬控制試驗,而對野外實地監測研究相對較少,通過人為設定或控制的水分梯度,往往難以反映濕地復雜的水文條件和特定植物的水文經歷,而開展自然狀態下濕地植物生長發育過程及其與水分條件之間關系研究,對于深入了解濕地植被格局形成、植物群落演替及進行濕地生態調控與恢復具有更為重要的理論和現實意義。
鄱陽湖濕地是我國重要的淡水湖泊濕地之一,受長江及“五河”(贛江、撫河、饒河、信江和修水)來水的共同作用,水位季節變化明顯,呈現豐水期和枯水期交替出現的獨特水文節律[24],濕地洲灘各高程帶“淹沒-出露”過程不一,這種不同的水文經歷使洲灘植被沿高程梯度呈現出較為明顯的帶狀分布格局[13]。近年來,隨著氣候變化和三峽水利工程興建運行等人類活動的干擾,鄱陽湖水文情勢發生顯著改變,主要表現為豐水期水位降低、枯水期提前和低水位持續時間延長[25- 26],這勢必會影響到濕地植物的生長發育、分布格局與群落結構,進而影響濕地生態系統功能的發揮[12,27]。但目前針對鄱陽湖濕地植物與水文變化之間關系研究還十分薄弱,對于濕地典型植物在生長發育過程中對不同水文條件采取的適應策略以及植物定植生長所需的水文條件閾值尚不清晰,為此,本文選取鄱陽湖濕地洲灘優勢植物——灰化苔草作為研究對象,基于野外實地觀測,研究自然狀態下鄱陽湖濕地不同高程帶灰化苔草種群發育過程及其與水文條件之間的響應關系,以認清鄱陽湖濕地節律性水文變化背景下灰化苔草的生態適應策略、響應方式和水文生態閾值,為進一步揭示鄱陽湖濕地植被帶狀分布格局形成及植物群落演替機制提供基礎理論支持,同時也為鄱陽湖濕地水文調控、生態恢復與生物多樣性保護提供重要科學依據。
1 材料與方法
1.1 研究區概況
鄱陽湖位于江西省北部,長江中下游南岸,地理位置24°29′14″—30°04′41″N、113°34′36″—118°28′58″E。上承“五河”(贛江、撫河、饒河、修水、信江)及區間來水,經由湖口調蓄后匯入長江,與長江存在著復雜的水文、水動力交互,是一個典型的吞吐型湖泊。由于受上游來水和長江的頂托、拉空和倒灌等作用的綜合影響,鄱陽湖水位年內變化劇烈,年內最大變幅可達9.7—15.79 m,呈現出“洪水一片,枯水一線;高水是湖,低水似河”的獨特自然景觀[6,28]。湖區地處丘陵平原區,四周群山環繞,整個湖盆自東南向西北傾斜。以松門山為界,可將鄱陽湖分為南北兩部分,南部寬淺,為主湖區;北部狹深,為入江水道。湖泊地貌主要有洲灘、島嶼、內湖、水道和汊港等類型[29]。該地區氣候上屬于亞熱帶季風氣候,雨熱同期,四季分明,年均氣溫16.5—17.8℃,無霜期250—280 d,年均日照時數約為1970 h,年均太陽總輻射量可達4500×106J/m2,而多年平均降水量為1350—1700 mm[30]。湖區洲灘土壤主要有草甸土、草甸沼澤土和沼澤土等類型[31]。由于水熱資源較為充足和水位周期性漲落變化,鄱陽湖區發育了大片的天然濕地,成為我國生物多樣性最為豐富的地區之一[32]。從濕地邊緣向中心,植物呈較為明顯的帶狀或環帶狀分布特征,依次分布著以狗牙根(Cynodondactylon)、蒿(Artemisiaspp.)等為主的中生植物群落,以蘆葦(Phragmitesaustralis)、南荻(Triarrhenalutarioriparia)等為主的挺水植物群落,以苔草(Carex)、虉草(Phalarisarundinacea)等為主的濕生植物群落和以馬來眼子菜(Potamogetonmalaianus)、苦草(Vallisnerianatans)、菱(Trapabispinosa)、荇菜(Nymphoidespeltata)等為主的沉水/浮葉植物群落[6,33]。其中灰化苔草是鄱陽湖濕地分布最廣的洲灘植物種,主要分布在洲灘的中低高程帶上。
1.2 樣地設置
本研究選取灰化苔草分布集中的蚌湖東側修水入湖口洲灘作為野外觀測樣地,該洲灘高程在15 m左右,洪水期被湖水淹沒,枯水期出露,洲灘上植物沿高程呈現出較為明顯的帶狀分布,隨著高程由低到高依次分布著虉草、灰化苔草、南荻等植物群落,其中灰化苔草面積最大,生長也較為茂盛,分布在12—14.5 m的高程帶上。洲灘未經開墾、放牧、火燒等人為干擾,自然環境保存良好。分別在洲灘四周按與湖岸垂直方向(沿高程或水分梯度)各布設1條樣帶,樣帶寬度為20 m,樣帶長度以整個洲灘上灰化苔草出現的最低和最高高程來進行確定;在每個樣帶內按0.5 m高程距離布設采樣點,在同一高程帶上,按等距離設立3個面積為1 m×1 m的樣方,用GPS-RTK(南方測繪,靈銳S86)確定各樣方的高程和地理位置。
1.3 研究方法
2013年3—5月在設置的樣地內進行灰化苔草種群生態觀測和樣品采集(灰化苔草在2月下旬萌發,3—5月為其生長季,5月下旬洲灘被湖水淹沒,灰化苔草死亡或沉入水底休眠,完成其一個生長周期),采樣時間間隔15天左右。對每個樣方分別測量灰化苔草種群的高度、密度、地上生物量和地下生物量等生態指標。其中,用標尺測定灰化苔草株高;用計數法測定種群密度;地上生物量采用收獲法,地下生物量采用挖掘法。采集10株植物樣品帶回實驗室處理,先用清水洗凈地下根系,用吸水紙吸干后,分別對地上和地下部分進行濕重稱量,之后在鼓風干燥箱中先于105℃下殺青10分鐘,再在80℃下烘干至恒重,稱其干重,最后換算成單位面積的地上及地下生物量,兩者相加計算出總生物量。用總生物量干鮮比(DW/FW)反映植物體含水量的多少(干鮮比=干重(Dry Weight)/濕重(Fresh Weight) ×100%);用生物量增長速率分析方法中的絕對增長速率(Absolute growth rate, AGR)和相對增長速率(Relative growth rate, RGR)反映灰化苔草種群生物量的積累狀況,計算公式分別為:
(1)
(2)
式中Bi、Bi+1分別是ti、ti+1時刻的灰化苔草種群的生物量(g/m2)。
1.4 數據處理與分析
借助Excel 2007進行數據處理和制圖。利用SPSS 19.0對測定數據進行統計分析,采用相關分析確定灰化苔草種群生態指標與水文參數之間的相關性;通過單因素方差分析(one-way ANOVA)和LSD多重比較法進行不同高程帶生態指標差異顯著性檢驗。
2 結果與分析
2.1 灰化苔草種群特征及其變化
由于13 m以下高程帶的灰化苔草在3月下旬即被湖水淹沒,因此,在進行灰化苔草種群發育過程研究時,主要針對13 m及以上高程帶的灰化苔草種群進行分析。
2.1.1 密度與株高
初步研究表明,在生長季內,各高程帶灰化苔草的種群密度變化趨勢基本一致,表現為隨時間推移而逐漸增加,但變化過程略有差異(圖1)。14.5 m高程帶灰化苔草種群密度在3月下旬之前增加較快,而之后基本保持穩定,維持在1875—1900株/m2;13 m和13.5 m高程帶的種群密度在3月上旬至4月中旬增加較為緩慢,4月下旬之后增加速度較前期有所加快;而14 m高程帶種群密度在3月下旬之前增加較慢,之后開始加快,到4月下旬增加速度再次放緩。各高程帶灰化苔草種群的平均株高與密度變化趨勢基本一致(圖1),整體表現為隨時間推移而逐漸增高,但不同生長階段增長幅度存在明顯差異。在4月下旬之前,株高的增長速度隨高程的增加而增大,而4月下旬之后,14 m和14.5 m高程帶增長速度明顯減慢,而13 m和13.5 m高程帶增長速度則快于前期。
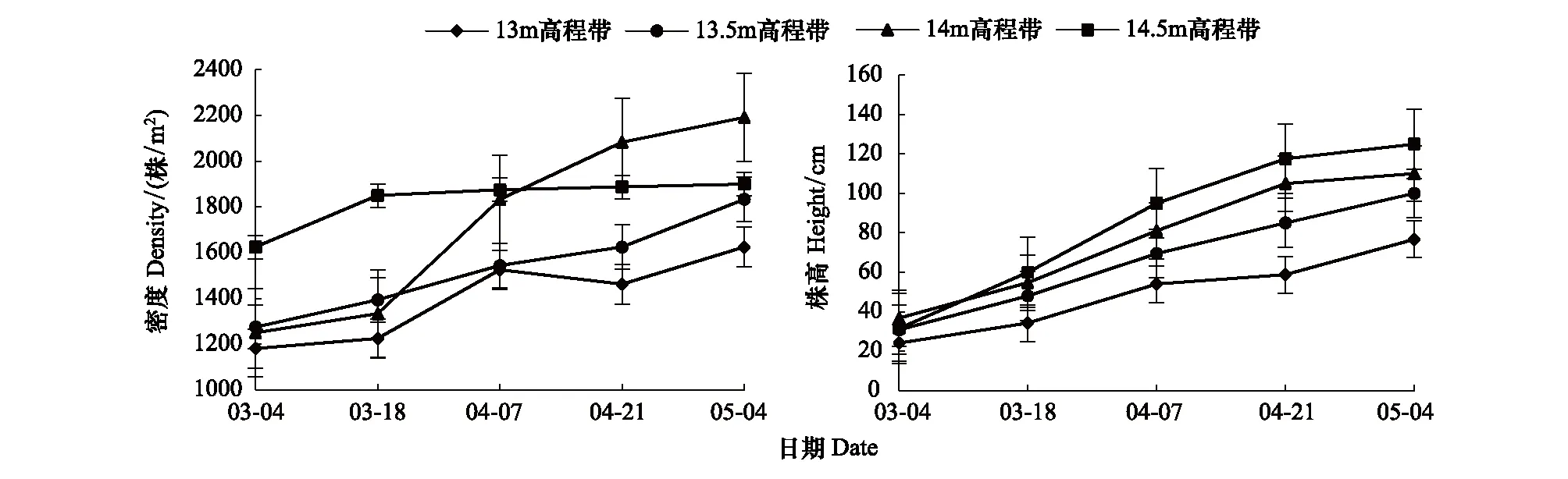
圖1 灰化苔草種群密度與株高變化Fig.1 Changes of height and density of Carex cinerascens population in different elevation zones
2.1.2 生物量
各高程帶的灰化苔草生物量變化及累積過程有所不同(圖2)。在生長季內地下生物量與總生物量一直處于增加狀態;而地上生物量除13 m高程帶呈不斷增加外,其他高程帶上均表現為先增加后減少的變化過程。從絕對增長速率來看,各高程帶灰化苔草地下生物量最大絕對增長速率出現在4月下旬—5月上旬(圖3),而地上生物量與總生物量的最大絕對增長速率出現時段不同,整體表現為13.5 m及以上高程帶出現在4月中旬,13 m高程帶則出現在4月下旬—5月上旬;這表明灰化苔草快速生長期出現時段隨高程而變化,高程越高,則出現越早。從相對增長速率來看,各高程帶地下生物量與總生物量一直處于累積過程,地上生物量在13 m高程帶上也一直處于累積過程,而13 m高程帶以上則在4月下旬之前處于累積過程,之后開始衰退,生物量出現降低。
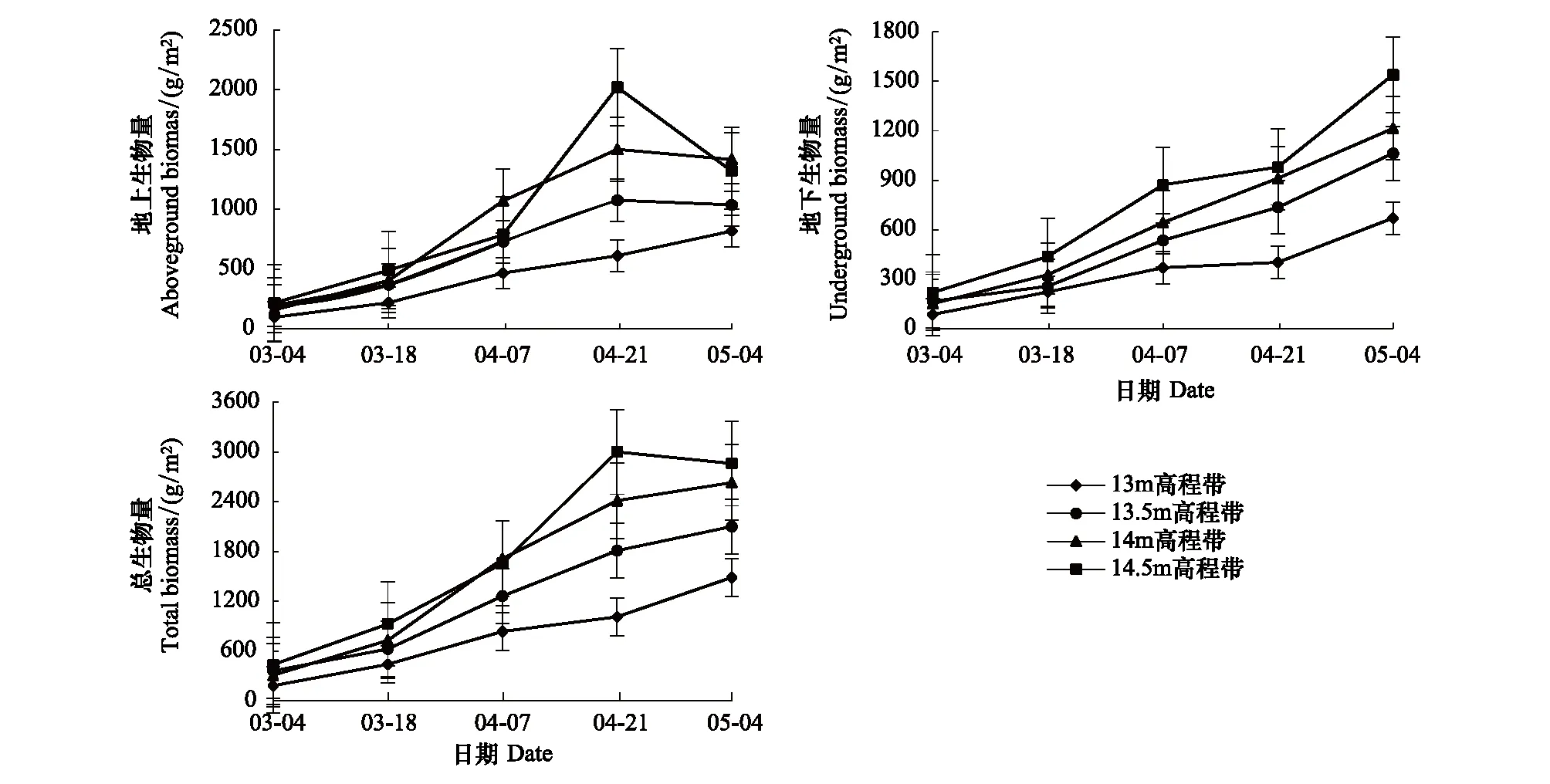
圖2 灰化苔草種群生物量變化Fig.2 Changes of aboveground, underground and total biomasses of Carex cinerascens population in different elevation zones
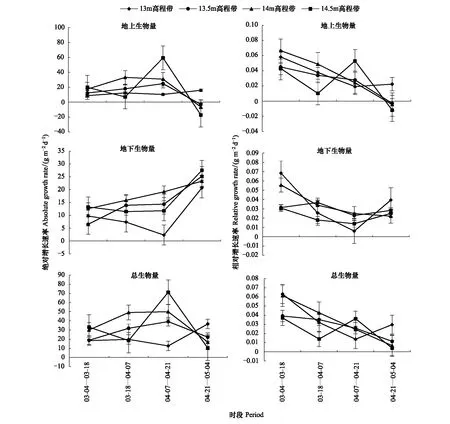
圖3 不同生長階段灰化苔草種群生物量增長速率變化Fig.3 Changes of absolute growth rate (AGR) and relative growth rate (RGR) of aboveground, underground and total biomasses of Carex cinerascens population in different elevation zones
2.1.3 干鮮比
各高程帶灰化苔草在生長季內的干鮮比隨時間變化過程基本一致(圖4),整體均呈增加趨勢,在3月上旬灰化苔草生長的初期階段,干鮮比較低,在20.8%—23.2%之間;進入4月中旬的快速生長期階段,灰化苔草的干鮮比有所增大,為23.3%—26.1%;到5月上旬的成熟期,灰化苔草干鮮比達到整個生長季的最大值,為26.8%—29.1%。從灰化苔草干鮮比的變化過程可以看出,生長初期的干鮮比要小于生長后期,這表明隨著時間的推移,灰化苔草植物體內的水分含量逐漸減少,而積累的干物質含量逐漸增多。

圖4 灰化苔草干鮮比變化 Fig.4 Change of DW/FW of Carex cinerascens population in different elevation zones
2.2 鄱陽湖濕地水文條件空間分異
淡水濕地植物的分布和生長發育是對周期性水文節律長期適應的結果。在灰化苔草相鄰的兩個同期生長季間(2012年5月4日—2013年5月4日),鄱陽湖水位(星子站,吳淞高程,下同)變化十分顯著(圖5),2月份湖水位達到最低,為8.91 m(2013年2月6日),8月份湖水位最高,為19.64 m(2012年8月13日),最大水位差達10.73 m。在最高與最低水位之間,湖泊水位呈現出明顯的漲水期和退水期。對于鄱陽湖洲灘不同高程帶而言,由于水文經歷的差異,在各高程帶上形成了不同的水文條件,表現出一定的水分梯度變化。在此將各高程上的水文條件以淹沒期平均水深(淹沒的總水深與淹沒天數的比值)、淹沒頻率(淹沒天數與計算總天數的比值)和平均距湖水面高度(高程與日水位差值之和與計算總天數的比值,水面以下為-,水面以上為+)來進行反映。在研究期間,洲灘上灰化苔草分布的下限12 m高程帶大部分時間被湖水所淹沒,淹沒期水深在0.02—7.64 m之間,平均值為3.89 m,相鄰同期生長季內淹沒頻率為60.38%,平均距湖水面高度為-1.73 m;隨著高程的增加,洲灘淹沒天數逐漸減少,而出露天數逐漸增多,至洲灘灰化苔草分布上限14.5 m高程帶上,淹沒期平均水深減小到2.78 m,該高程帶上淹沒頻率降低到42.08%,平均距湖水面高度增大為0.77 m(表1)。總體來看,洲灘上灰化苔草定植與生長的水文條件閾值為淹沒期平均水深2.78—3.93 m,淹沒頻率42.10%—60.38%,而平均距湖水面高度則為-1.73—0.77 m。
2.3 不同高程灰化苔草種群特征差異及對水文條件響應
選取灰化苔草接近成熟期的4月下旬進行不同高程帶灰化苔草種群特征的差異分析。由表2可知,灰化苔草種群密度隨高程呈波動變化,差異性分析顯示不同高程帶灰化苔草種群密度差異不顯著(F=2.171,P>0.05),這說明高程形成的水分梯度對種群密度無明顯影響。
不同高程帶灰化苔草平均株高隨著高程的升高而逐漸增加,高程間存在顯著差異(F=5.879,P<0.05),14.5 m高程的平均株高達到117.5 cm,是13 m高程的兩倍。多重差異性比較發現,13 m高程帶灰化苔草株高與13.5 m高程帶差異不明顯,而與14 m及14.5 m高程帶形成顯著差異,其他高程帶間差異則不明顯,這說明13.5以下高程的水文條件對灰化苔草株高的增加起到抑制作用,而13.5 m及以上高程帶的水文條件更有利于灰化苔草的伸長生長。
不同高程帶灰化苔草生物量均隨高程的升高而增大,且高低灘位間相差一倍以上,高程間亦存在顯著差異(地上生物量:F=5.022,P<0.05;地下生物量:F=5.485,P<0.05;總生物量:F=6.599,P<0.05)。多重差異性分析顯示,13 m高程帶的地下生物量與總生物量與其他高程帶存在顯著差異,其他高程帶之間則差異不顯著;而地上生物量13 m高程與13.5 m高程無明顯差異,但與14、14.5 m高程帶差異顯著,而其他高程帶間的地上生物量無明顯差異。可以看出,13.5 m及以上高程帶的水文條件更有利于灰化苔草生物量的形成與積累,而13 m高程帶的水文條件對生物量積累相對不利。
將4月下旬灰化苔草各生態指標與淹沒期平均水深、淹沒頻率、平均距湖水面高度3個水文參數分別做相關分析,結果顯示,除密度以外,其他各指標與水文參數均呈顯著負相關(P<0.01),這進一步說明水文條件對灰化苔草生長發育產生重要影響。
由上述分析可知,灰化苔草生長發育對高程形成的不同水分梯度產生差異性響應。在13.5 m以下的低灘位,由于頻繁的過度淹沒而使灰化苔草生長受到水分條件的抑制,植株矮小,生物量積累相對較少;而在13.5 m及以上中高灘位,形成灰化苔草生存有利的水文環境,更利于灰化苔草的伸長生長和生物量的形成與累積。結合前面水文參數的分析,可對春季生長季內灰化苔草生長發育的適宜水文條件做出初步判斷,有利于灰化苔草生長所需的淹沒期平均水深閾值為 3.70—2.78 m,淹沒頻率閾值為42.08%—43.17%,平均距湖水面高度閾值為-0.23—0.77 m。
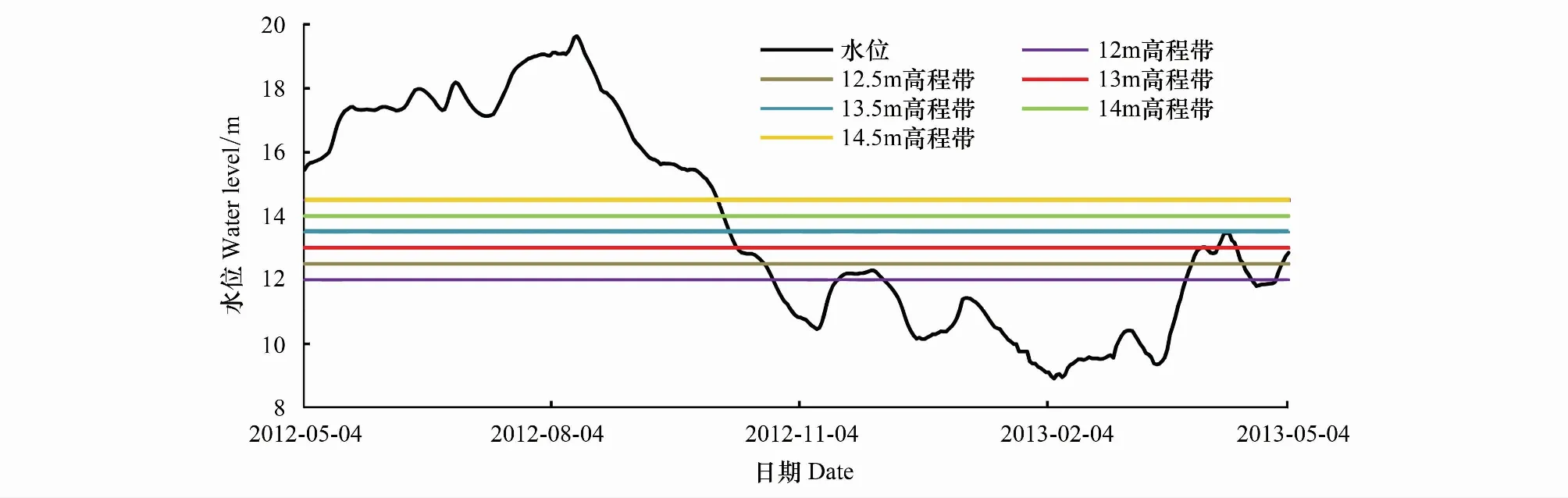
圖5 鄱陽湖濕地星子站水位變化Fig.5 Water-level variation of Xingzi hydrological station in Poyang Lake wetland during 2012/5/4—2013/5/4
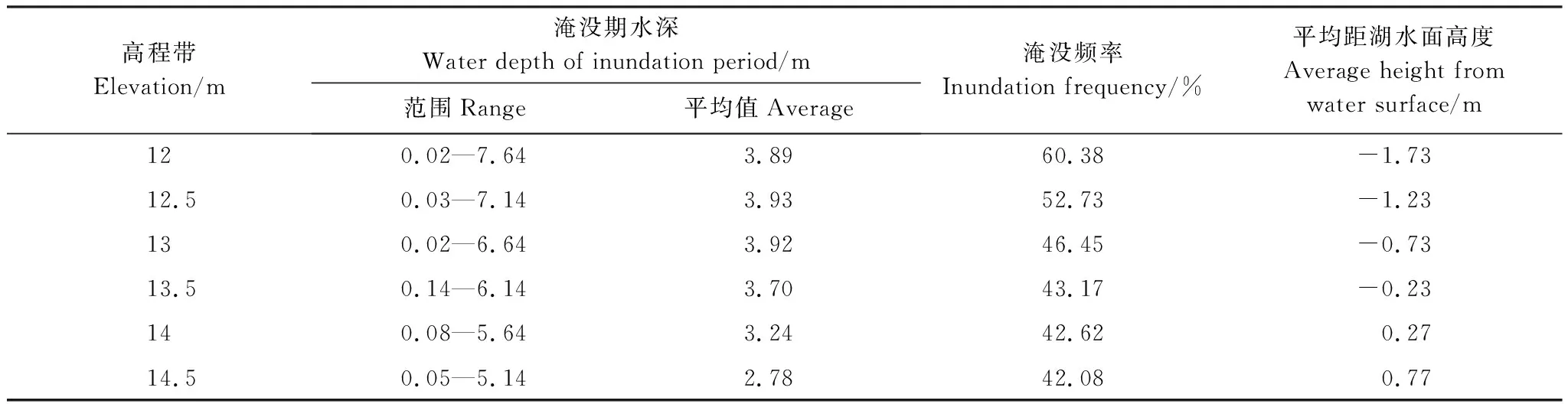
高程帶Elevation/m淹沒期水深Water depth of inundation period/m范圍Range平均值Average淹沒頻率Inundation frequency/%平均距湖水面高度Average height from water surface/m120.02—7.643.8960.38-1.7312.50.03—7.143.9352.73-1.23130.02—6.643.9246.45-0.7313.50.14—6.143.7043.17-0.23140.08—5.643.2442.620.2714.50.05—5.142.7842.080.77
3 討論
3.1 灰化苔草水分適應策略
淡水濕地植物的空間分布及其生長發育過程與高程、水文狀況、土壤性質以及植物種類等因素密切相關,高程及其形成的水文條件起著至關重要的作用[2,34]。同一植物種類處在不同的水文條件下,植物會通過形態和自身生理機能的調整來適應變化的水文環境,形成能夠反映生境條件的生態與生理特征[35]。對于水位周期性波動的淡水濕地來說,洲灘或湖(河)濱帶經常會遭受不同程度的淹水脅迫[26],濕地植物往往通過枝條伸長生長的逃避策略或者休眠、緩慢生長的耐受策略來應對水分脅迫[36-37],不同的淹水條件下各種植物會采取不同的適應策略。已有研究表明,在淹水條件下雙穗雀稗(Paspalumdistichum)、空心蓮子草(Alternantheraphiloxeroides)和牛鞭草(Hemarthriaaltissima)等濕地植物會通過伸長生長來逃離淹水環境[38],而虉草和狗牙根(Cynodondactylon)則通過減緩莖的伸長和不產生新的分枝等方式來提高自身對水淹的耐受性[39]。
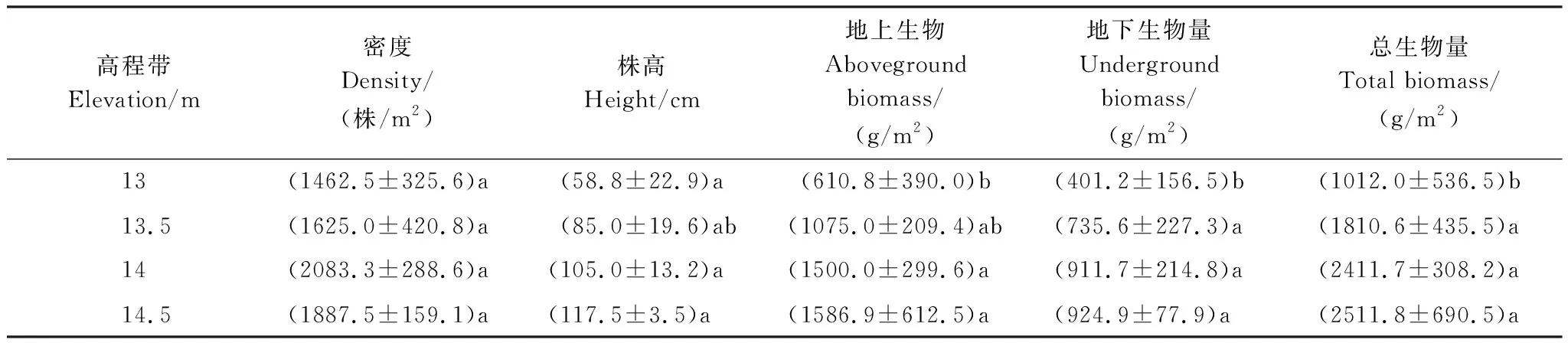
表2 不同高程帶灰化苔草種群特征及差異(n=13)
數據為平均值±標準差;每列數據右上角標有不同字母表示高程間存在顯著差異(P<0.05)
本研究發現,灰化苔草在生長季會根據水淹狀況來調整自身的生存策略。13 m以下高程帶的灰化苔草在3月下旬即被湖水淹沒,長期的水淹導致水中供氧和光照不足,光合作用和葉綠素合成受到嚴重抑制,灰化苔草通過地上植株死亡和地下根系休眠的耐受策略來增加存活率;13 m高程帶在4月上旬一直處于半水淹狀態,該期間灰化苔草株高和生物量增長速度減緩,而在4月中下旬湖水位發生波動式的下降,灰化苔草經歷間歇性的半水淹,但大部分時間植株完全出露水面,這一期間灰化苔草生長發育過程明顯加快,株高及生物量的增加幅度達到整個生長季的最大值,灰化苔草通過快速伸長生長的逃避策略來應對短期的間歇性半水淹環境;在13.5 m及以上高程帶上,灰化苔草5月上旬之前基本未遭受淹水脅迫,株高和生物量在各生長階段均高于13 m高程的低灘位,且生長后期差異更加明顯,然13.5 m及以上高程帶由于萌芽較早,在4月下旬即步入成熟期,該階段雖然株高仍在增長,但生物量開始逐漸向地下部分轉移以完成生活史,導致地上生物量出現一定程度的降低,但13.5 m和14 m高程帶的總生物量仍在增加,而14.5 m高程帶由于提前進入枯萎衰落期,有機質合成減弱,呼吸消耗增多,總生物量有所下降。在5月中旬之后,湖水位大幅上漲,鄱陽湖進入豐水期,灰化苔草分布洲灘完全被湖水淹沒,各高程帶地上部分逐漸死亡,灰化苔草進入休眠狀態,完成其完整的春季生長季。由此可見,各高程帶灰化苔草對不同的水文經歷采取差異性適應策略,這種生存策略是灰化苔草對鄱陽湖濕地周期性水文節律長期適應進化的結果。
3.2 灰化苔草水分利用策略
濕地植物在不同生長階段會根據實際的水分條件來調節自身的水分利用策略,通過植物干鮮比可以對其采取的水分利用策略予以反映。植物干鮮比標志著植物體含水量的多少,它是植物對水分條件以及植物不同生長階段所表現出的一種響應,濕地植物對不同的水分條件會產生趨異響應[17]。上官鐵梁等[40]通過對汾河河漫灘濕地植物群落研究發現,植物干鮮比隨著群落水分增加而增大;而王麗等[17]對三江平原濕地小葉章群落研究也發現,在生長季初期小葉章主要通過儲水策略來保持體內一定的儲水量,干鮮比較低,在生長季后期由于可獲得足夠的水分資源,干物質積累多且迅速,干鮮比較高。在本研究中,鄱陽湖濕地灰化苔草也采用相似的水分利用策略,在生長初期干鮮比較低,而生長中后期則顯著增大,這種變化一方面與灰化苔草自身的生長特性有關,另一方面也與鄱陽湖濕地特定的水文過程有關。在灰化苔草生長初期,由于植株矮小,氣溫較低,灰化苔草蒸騰作用和呼吸作用較弱,而此時正值鄱陽湖區的少雨季,濕地處于一年當中的低水位期,水分條件十分有限,灰化苔草主要采取儲水策略以維持自身生長對水分的需求,干鮮比較低;到灰化苔草生長的中后期,由于降水增多以及湖水位的抬升,水分資源較為充足,灰化苔草進入快速生長期,但該時期灰化苔草的蒸騰作用和呼吸作用明顯增強,植株體內的水分被大量消耗,而由光合作用形成的干物質較前期增多,引起干鮮比出現增大。灰化苔草生長過程中干鮮比的變化一定程度上也反映出其對鄱陽湖濕地水位節律性變化的生態適應。
3.3 灰化苔草水文條件閾值
每種濕地植物在生長發育過程中對水文條件都有特定的需求,在某個閾值內植物可以存活并生長。眾多水文因子中,水深和淹沒頻率是影響濕地植物定植生長、繁殖和分布的重要環境因子[41- 42]。對于濕地沉水植物而言,其整個生長發育過程均在水中進行,水深通過影響水下光照、風浪和底泥特征來作用于植物生長繁殖,如黑藻(Hydrillaverticillata)和苦草(Vallisnerianatans)分別在水深0.5—2.5 m和0.5—5.6 m才能夠定植生存,當水深繼續增加時,光照強度會低于光補償點,光合作用受到抑制,植物將難以存活[43-44]。而對于某些濕地挺水植物和濕生植物來說,在生長過程中會經歷一定程度的水淹和干旱,因此,除水深因子外,植物定植生長也會受到淹沒頻率的影響[41]。已有研究表明,適度的干濕交替有利于挺水植物和濕生植物的生長繁殖,而長期的過度水淹和無水淹會減弱植物光合作用和養分吸收能力,對植物生長起到明顯的抑制作用[35,45]。但不同濕地植物對水文條件的喜好性和耐受性有所差異,如鄱陽湖濕地南荻生長于相對干旱的環境中,不能忍受長時間的洪水淹沒,其主要集中分布于平均水深2.2—2.4 m、淹沒頻率15%—40%的區域,而假儉草(Eremochloaophiuroides)對水文條件適應范圍較寬,在平均水深1.8—14.6 m、淹沒頻率12%—100%的區域均可定植生長[42]。灰化苔草是鄱陽湖濕地洲灘典型的濕生植物,生長于相對濕潤的環境中,對水淹和干旱有一定的耐受能力,具有較大的水分生態幅[46],適度的水淹和露干交替有利于生長所需水分的獲取和光合作用的進行,可促進灰化苔草的伸長生長和生物量的累積,而長期水淹或干旱將超出灰化苔草的耐受極限,不利于植株體的存活和種群的發育。本研究發現,灰化苔草在淹沒期平均水深2.78—3.93 m、淹沒頻率42.1%—60.38%、平均距湖水面高度-1.73—0.77 m的水文條件下能夠定植生長,而淹沒期平均水深3.70—2.78 m、淹沒頻率42.08% —43.17%、平均距湖水面高度-0.23—0.77 m的水文環境則對灰化苔草的生長更為有利,其種群的平均高度和生物量均達到最大。
許多研究表明,部分濕地植物種群的生態特征(如株高、生物量)對水文因子(如水深)的響應符合高斯分布[47-49],然在野外環境中,影響濕地植物種群的環境因子眾多,除水文條件以外,植物種內及種間競爭、土壤養分、溫度、降水等環境因子也會產生影響,實際上濕地植物種群生態特征是多環境因子綜合作用的結果[50],鄱陽湖濕地灰化苔草種群生態指標沿水分梯度并未呈現出明顯的高斯分布,而是在水分閾值內大致隨水分梯度的降低而增加,這種分布格局的形成與其他環境因子的空間分異及作用強度也存在一定關系,但對于其具體的形成機制和作用機理還有待今后進一步地深入研究。
4 結論
(1)鄱陽湖濕地灰化苔草作為洲灘優勢植物種群之一,在春草生長季內各高程帶種群變化規律基本一致,但由于高程帶間水文條件與水文經歷的不同,灰化苔草生長發育過程對節律性水文波動產生差異性適應和響應。在13.5 m以下的低灘位,由于頻繁的水淹而使灰化苔草生長發育受到水分條件的抑制,植株矮小,生物量積累相對較少;而在13.5 m及以上中高灘位,形成灰化苔草適生的水文環境,更利于灰化苔草的伸長生長和生物量的形成與積累。
(2)灰化苔草在不同生長階段會根據實際的水分條件來調節自身的水分利用策略。生長初期灰化苔草主要通過儲水策略來應對水分相對缺乏形成的干旱脅迫;而生長末期則主要通過增加干物質含量來應對湖水位上漲形成的淹水脅迫。
(3)灰化苔草定植與生長發育對水文條件存在一定的適應范圍。在淹沒期平均水深2.78—3.93 m、淹沒頻率42.1%—60.38%、平均距湖水面高度-1.73—0.77 m的條件下灰化苔草能夠定植生長;而淹沒期平均水深3.70—2.78 m、淹沒頻率42.08%—43.17%、平均距湖水面高度-0.23—0.77 m的水文環境則更有利于灰化苔草的生長發育。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
