六倍體小黑麥重要性狀的改良潛力探究
2018-08-28 08:40:36崔紫霞張傳量許小宛孫道杰
麥類作物學報 2018年8期
崔紫霞,張傳量,馮 潔,許小宛,孫道杰
(西北農林科技大學農學院,陜西楊凌 712100)
近年來,小黑麥以強大的生物量以及優良的營養品質成為值得倡導的飼料作物[1-2]。小黑麥(Triticale)是由小麥屬(Triticum)和黑麥屬(Secale)物種經屬間有性雜交和雜種染色體數加倍后人工合成的新物種,不僅擁有小麥高產和優質的特點,還具有生長茂盛、莖葉多汁、含糖量高、綜合抗性強、分蘗力強、大穗大粒、籽粒中蛋白質及賴氨酸含量高等優良性狀[3-5]。小黑麥在生產上應用最多的是八倍體和六倍體。六倍體(2n=6X=42,AABBRR)小黑麥是四倍體小麥和二倍體小黑麥雜交后再經過染色體加倍形成的異源六倍體。我國生產中應用的六倍體小黑麥分為糧用型及糧飼兼用型[4]。前者一般籽粒產量較高、株高與小麥相近,后者則株高較高,生物產量大。
苗相是鑒定小黑麥材料為冬性或春性的標準,越匍匐說明該品種冬性越強,但冬性越強并不能說明抗寒性越強。研究表明,春化和抗寒性由不同基因控制,冬性強的品種其春化溫度低、時間長,低溫春化過程也是幼苗抗寒性形成的過程,有利于增強幼苗的抗寒性[6-8]。
株高、穗長、穗頸長、穗下節長和倒二節長屬于株型性狀,單株穗數、小穗數、穗粒數和千粒重屬于產量相關性狀[9-10]。何賢芳等[9]研究表明,產量與旗葉寬和穗長顯著或極顯著正相關;吳同彥等[11]研究發現,穗長與千粒重呈不顯著負相關,穗下節長與穗粒數呈正相關,倒二節長與旗葉寬呈顯著正相關。海 燕等[12]利用小麥花藥培養技術構建的DH群體對株高、單株穗數、穗長、穗下節長等性狀進行了遺傳研究,發現被測性狀均有較高的遺傳力。王瑞清等[13]用6個小黑麥品種按完全雙列雜交方法配置雜交組合,對親本、F1、F2的產量性狀進行了遺傳分析,發現提高穗粒數和千粒重可以有效增加產量。胡立芹等[14]分析了從CIMMYT引進的111份六倍體小黑麥材料的遺傳多樣性特點,發現農藝性狀和品質性狀均表現出較大的差異,具有豐富的多樣性。
利用冬、春性小黑麥雜交培育廣適性小黑麥的研究開始于20世紀70年代,雜交所產生的遺傳變異能使春化和光照發育特性產生一系列變異,不僅表現在籽粒灌漿期、灌漿速度、成熟期的變異上,還表現在抗病性、抗逆性、品質及產量性狀上[15]。艾尼瓦爾等[16]認為,在新疆選育與推廣冬、春性雜交小黑麥品種能與新疆復雜的生態條件以及耕作制度相適應。馬翎健等[17]曾對不同冬春性類型小麥進行組配,發現“冬麥×春麥”雜交組合的F1代穗分化優勢、產量優勢明顯優于其他類型組合。曹新有等[18]研究發現,冬小麥在其他特性相當的情況下,抗寒性強的偏春性品種表現出更廣泛的適應性,可見冬、春麥雜交可以作為提高麥類作物廣適性的重要育種途徑。
本研究以小黑麥冬性與春性、春性與春性雜交的F1、F2群體為材料,進行六倍體小黑麥性狀的遺傳分析及改良潛力探究,為糧飼兼用型小黑麥育種提供借鑒。
1 材料與方法
1.1 供試材料以及田間種植
供試材料為冬性小黑麥蘭小黑(LXH)、春性小黑麥CM-12和CM-13及其雜交構建的CM-13/CM-12、CM-12/LXH和CM-13/LXH春/春型、冬/春型的F1和F2群體,試驗材料LXH由本試驗室保存,CM-12和CM-13由國際玉米小麥改良中心(CIMMYT)提供,3個材料均為糧飼兼用型小黑麥。3個群體的親本以及F1代和F2代分別于2015年10月-2016年6月和2016年10月-2017年6月種植于西北農林科技大學試驗田,群體小區行長2 m,行距25 cm,株距為15 cm,常規管理,生長期間未發生嚴重病蟲害。
1.2 農藝性狀的測定
小黑麥性狀調查參照《小麥種植資源描述規范和數據標準》。在越冬前苗期調查苗相,苗相作為冬春性研究的一個鑒定標志,分為直立、半匍匐、匍匐三種,春性材料為3級直立,冬性材料為1級匍匐。兩個春性親本均為3級直立,冬性材料LXH為1級匍匐。按單株分別記錄。
在小黑麥乳熟期,3個親本以及3個F1代每個供試材料選取10株調查株高、單株穗數、穗長、小穗數、穗粒數、穗頸長、旗葉長與寬、穗下節長和倒二節長;收獲后測定千粒重。3個F2群體每個群體隨機調查200株用于表型及遺傳分析。
1.3 數據處理
采用Microsoft Excel軟件計算配合力、平均數、變異系數、偏度以及峰度值,用IBM SPSS Statistics 22軟件計算3個小黑麥F2群體的9個性狀的相關系數和遺傳力。
遺傳力計算公式:
H2=VG/Vp×100%=VG/(VG+VE)×100%
式中,VP代表表型方差,VG代表遺傳方差,VE代表環境方差,H2代表遺傳力。
2 結果與分析
2.1 親本各個性狀的配合力分析
苗相是鑒定麥類作物冬、春性的主要依據,苗相一般配合力較小的品種在田間表現出較強的抗凍性。由表1、表2可以看出,LXH苗相的一般配合力最低,以LXH為親本的兩個組合苗相的特殊配合力均低于春/春型小黑麥組合,最低的是CM-12/LXH,說明冬/春型雜交有利于小黑麥的抗寒性改良,印證了前人的結論[18]。
小麥穗頸長與收獲指數及單株產量密切相關,在一定株高條件下,穗下節長度在30 cm左右時對產量有一定促進作用[19]。株高越高,越容易倒伏,不利于高產;但株高過低會減少小黑麥的生物量。親本LXH的株高、穗頸長、穗下節長和倒二節長的一般配合力均最高,其次是CM-13,最后是CM-12,但后兩者相差不大。株高、穗長、穗頸長、穗下節長和倒二節長特殊配合力最大的是CM-13/LXH組合,其次是CM-12/LXH組合,最小是CM-12/CM-13組合。關于穗長,親本CM-12的一般配合力最大,CM-12/LXH組合的特殊配合力最大。
就小穗數、旗葉長和旗葉寬而言,LXH的一般配合力最大,CM-13/LXH組合的特殊配合力最大;關于單株穗數,LXH的一般配合力最大,CM-12/LXH組合的特殊配合力最大;關于穗粒數,CM-13的一般配合力最大,CM-13/LXH的特殊配合力最大。綜合考慮,作為糧飼兼用型小黑麥最優的組合為CM-12/LXH,其次是CM-13/LXH,然后是CM-12/CM-13。
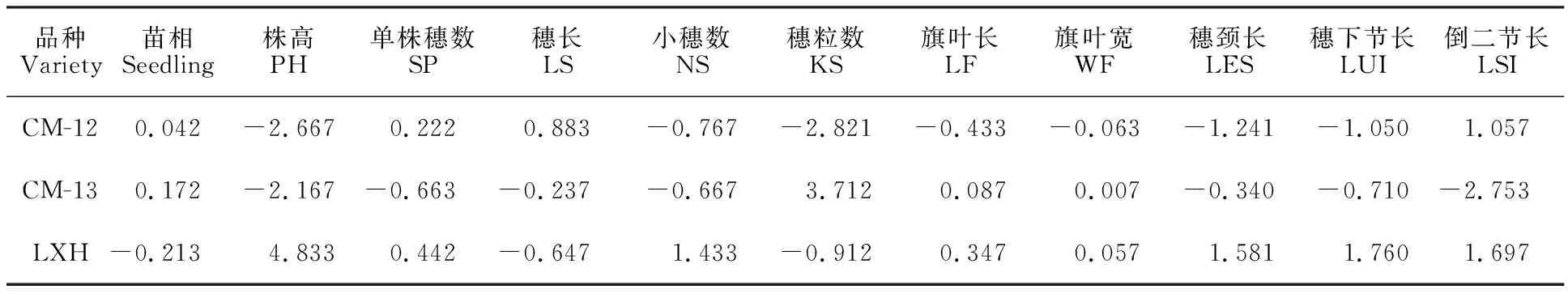
表1 3個親本各性狀的一般配合力Table 1 General combining ability of traits of three parents
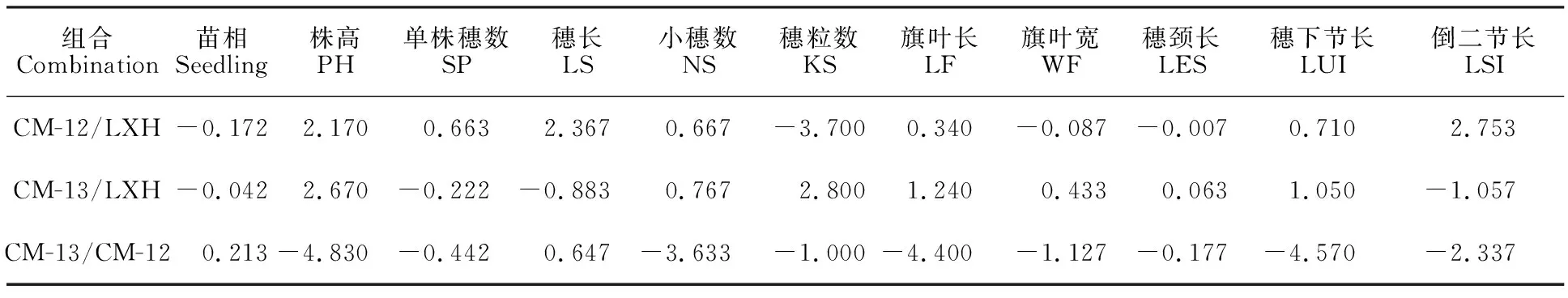
表2 3個組合的特殊配合力Table 2 Special combining ability of three combinations
由表3可以看出,在CM-12/LXH的F2群體中,株高與穗長、小穗數、旗葉長、旗葉寬、穗頸長、穗下節長、倒二節長呈極顯著正相關;穗長與小穗數、旗葉長、旗葉寬呈極顯著正相關,與穗下節長呈顯著正相關;單株穗數與穗頸長、穗下節長、倒二節長呈不顯著負相關;穗長與穗頸長呈不顯著負相關;小穗數與旗葉長、旗葉寬呈極顯著正相關,與穗下節長呈顯著正相關;旗葉長與旗葉寬、穗下節長呈極顯著正相關;旗葉寬與穗下節長呈極顯著正相關,與穗頸長呈顯著正相關;穗頸長與穗下節長、倒二節長呈極顯著正相關;穗下節長與倒二節長呈極顯著正相關。
在CM-13/LXH的F2群體中,株高與穗長、小穗數、穗頸長、穗下節長以及倒二節長呈極顯著正相關;單株穗數與穗長、旗葉寬、穗頸長、倒二節長呈極顯著正相關,與旗葉長呈顯著正相關,與倒二節長呈不顯著負相關;穗長與小穗數、旗葉長、旗葉寬、穗頸長、穗下節長呈極顯著正相關;小穗數與旗葉寬、穗下節長呈極顯著正相關,與旗葉長呈顯著正相關;旗葉長與旗葉寬、穗下節長呈極顯著正相關,與穗頸長呈顯著正相關,與倒二節長呈不顯著負相關;旗葉寬與穗頸長、穗下節長呈極顯著正相關,與倒二節長呈不顯著負相關;穗頸長與穗下節長呈極顯著正相關。
在CM-12/CM-13的F2群體中,株高與穗長、旗葉長、倒二節長呈極顯著正相關,與旗葉寬、穗下節長呈顯著正相關,與穗頸長呈不顯著負相關;單株穗數與穗長、旗葉長、旗葉寬呈極顯著正相關,與小穗數、穗下節長呈顯著正相關,與穗頸長呈不顯著負相關;穗長與小穗數、旗葉長、旗葉寬、穗下節長、倒二節長呈極顯著正相關,與穗頸長呈不顯著負相關;小穗數與旗葉寬、穗下節長呈極顯著正相關,與穗頸長呈顯著正相關;旗葉長與旗葉寬、穗下節長呈極顯著正相關,與倒二節長呈顯著正相關,與穗頸長呈不顯著負相關;旗葉寬與穗下節長呈極顯著正相關;穗頸長與穗下節長呈極顯著正相關,與倒二節長呈不顯著負相關。
在CM-12/LXH的F2群體中,株高、穗長、小穗數、旗葉長、旗葉寬、穗頸長、穗下節長、倒二節長之間相關性高,有些達到極顯著水平,小穗數、旗葉長、旗葉寬和穗頸長都能影響產量,說明在提高生物量性狀的同時,產量相關性狀也會提高,但是在CM-12/LXH的F2群體中,單株穗數跟莖稈特性呈顯著負相關或相關性較弱,所以在提高生物量性狀的同時要考慮單株穗數,使糧飼兼用型小黑麥達到最優性狀組合,從而達到高產目標。而其他兩個群體的產量相關性狀跟株型性狀之間的相關性較CM-12/LXHF2群體弱,在CM-13/LXH的F2群體中,單株穗數、穗長、小穗數跟其他性狀之間都具有正相關關系,相關程度不同,因此,在以收獲籽粒為目的的小黑麥育種中,CM-13/LXH優于其他兩個群體。
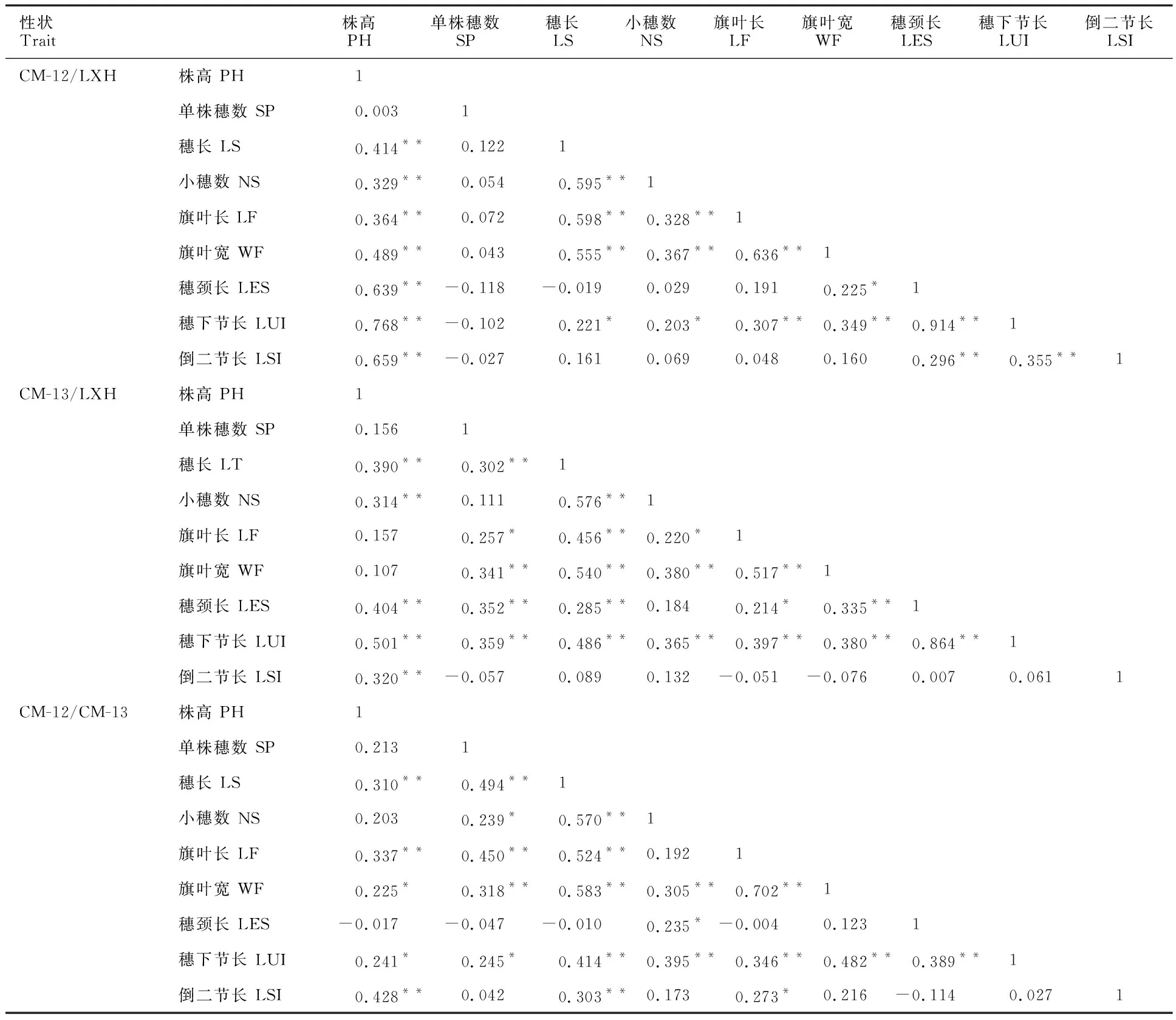
表3 3個群體各個性狀的相關系數Table 3 Correlation coefficients of each trait in the three populations
2.2 各性狀的變異系數及遺傳力比較
由表4可以看出,3個群體中,株高的變異系數為6.08%~11.78%,單株穗數的變異系數為34.30%~46.57%,穗長的變異系數為9.78%~12.53%,小穗數的變異系數為8.67%~11.73%,旗葉長的變異系數為14.51%~17.81%,旗葉寬的變異系數為10.65%~16.56%,穗頸長的變異系數為13.31%~23.04%,穗下節長的變異系數為10.26%~15.38%,倒二節長的變異系數為12.61%~14.01%。9個性狀變異程度表現為單株穗數>穗頸長>旗葉長>倒二節長>旗葉寬>穗下節長>穗長>小穗數>株高。CM-12/LXH的9個性狀變異系數均高于其他兩個群體,說明CM-12/LXH組合后代性狀分離范圍廣,變異類型多,多樣性豐富,進行新品種選育的潛力較大。
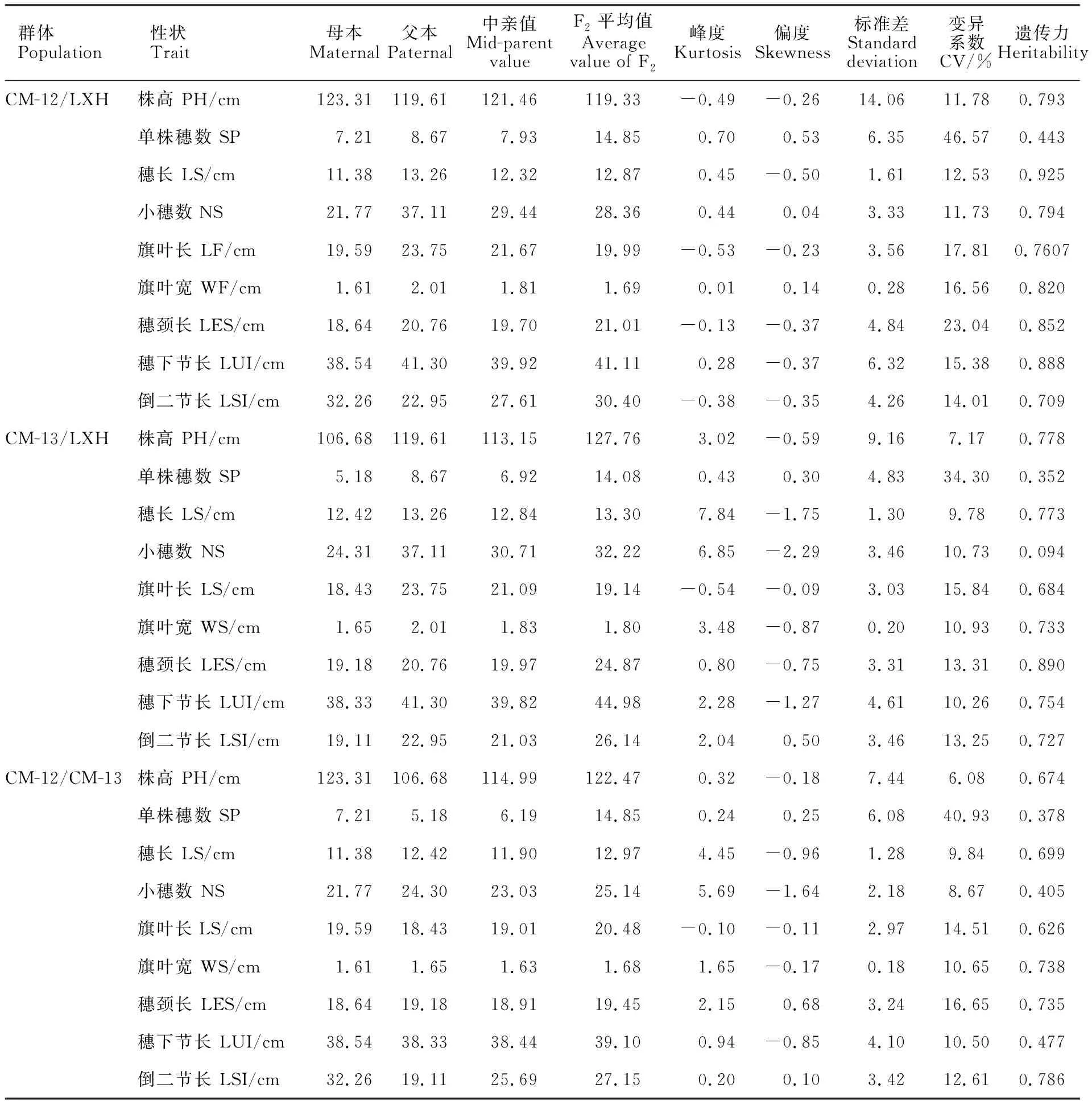
表4 F2群體與親本的各性狀值Table 4 Traits of F2 population and their parents
3個群體株高的遺傳力為0.674~0.793,單株穗數的遺傳力為0.352~0.443,穗長的遺傳力為0.699~0.925,小穗數的遺傳力為0.094~0.794,旗葉長的遺傳力為0.626~0.761,旗葉寬的遺傳力為0.738~0.820,穗頸長的遺傳力為0.735~0.890,穗下節長的遺傳力為0.477~0.888,倒二節長的遺傳力為0.709~0.786。9個性狀的遺傳力大小為穗頸長>穗長>旗葉寬>株高>倒二節長>穗下節長>旗葉長>小穗數>單株穗數。
在CM-12/LXH的F2群體中,遺傳力最小的是單株穗數(0.443),遺傳力最大的是穗長(0.925),其他性狀的遺傳力普遍高于其他兩個群體,說明CM-12/LXH的F2群體中各個性狀的遺傳力受環境影響較小。在CM-13/LXH的F2群體中,遺傳力最小的是小穗數(0.094),遺傳力最大的是穗頸長(0.890)。在CM-12/CM-13的F2群體中,倒二節長(0.786)的遺傳力最大,而單株穗數(0.378)的遺傳力最小。
分析3個群體9個性狀正態分布的偏度和峰度,推測在CM-13/LXH的F2群體中,株高可能存在微效基因間的互作;穗長、小穗數和穗頸長可能存在微效基因的重疊作用;旗葉寬、穗下節長和倒二節長可能存在微效基因間的互補作用。在CM-12/CM-13的F2群體中,穗長、小穗數、旗葉寬和穗頸長可能存在微效基因間互補作用。其他性狀的微效基因相互作用不明顯。
3 討 論
在小黑麥品種的選育中,正確選擇親本材料進行雜交組合配制是篩選優良品種的關鍵。親本的配合力是決定優良性狀傳遞穩定性的主要因素[20]。配合力受多種基因效應支配,一般配合力由基因的加性效應決定,是可以遺傳的性狀,一般配合力的高低由親本所含的有利基因位點的多少決定;特殊配合力由基因的非加性效應決定,受基因間的顯性、超顯性和上位性效應控制,只能在特定的組合中由雙親的等位基因或非等位基因間的互作反映出來,是不能遺傳的性狀[21-22]。本研究發現,在3個親本中,不論株型性狀還是產量性狀,LXH是最優親本;3個F2群體比較,CM-12/LXH的配合力最好,其株型性狀和產量性狀的相關系數、變異系數、遺傳力最高,具有較好應用潛力,可以用于糧飼兼用小黑麥的改良。CM-13/LXH的F2群體可以用于糧用小黑麥后期研究。本研究中,9個被測性狀變異系數表現為單株穗數>穗頸長>旗葉長>倒二節長>旗葉寬>穗下節長>穗長>小穗數>株高,9個性狀的遺傳力表現為穗頸長>穗長>旗葉寬>株高>倒二節長>穗下節長>旗葉長>小穗數>單株穗數。
單株穗數一直是育種家關注的麥類作物產量構成的重要因素[23],馮春葉等[24]認為,拔節前小麥主莖和分蘗對營養物質的競爭會影響最終成穗數;趙廣才等[25]提出了優勢蘗組概念;封超年等[26]認為,莖蘗成穗率與產量線性正向相關。雖然單株穗數對于麥類作物很重要,但對于小黑麥的分蘗性狀研究的較少,程治軍等[27]用八倍體小黑麥研究了 Rht12對小黑麥株高、單株分蘗數和種植飽滿度的影響發現, Rht12的導入提高了八倍體小黑麥的分蘗力。本研究中,單株穗數的變異系數高達46.57%,這對于糧飼兼用型小黑麥的單株穗數來說育種潛力較大,具體生理生化機制還需進一步研究。
旗葉是麥類植物籽粒碳水化合物的主要合成源之一,而旗葉長和旗葉寬直接影響光合速率。研究表明,旗葉性狀和穗粒重、千粒重、穗粒數和小麥產量間存在顯著正相關關系[28]。但是旗葉并不是越長越好,高產小麥株型一般要求葉片上挺、旗葉寬,跟莖稈的夾角小,而旗葉的長度對產量影響不明顯[29-31]。田間觀察發現,CM-12/LXH的F2群體和CM-13/LXH的F2群體的旗葉普遍上挺,說明這兩個群體的旗葉符合高產株型對葉片的要求,并且遺傳力均較高,可以用于高產小黑麥品種的選育。
