牛源防御素類抗菌肽的生物信息學分析
2018-08-25 06:04:22杭柏林董萌萌徐彥召張慧輝胡建和
現代畜牧獸醫 2018年7期
關鍵詞:分析
張 煒,杭柏林,董萌萌,胡 斌,徐彥召,張慧輝,胡建和
(河南科技學院動物科技學院,河南 新鄉 453003)
防御素(defensins)是一類陽離子多肽,是抗菌肽家族中的重要成員,有多種生物學活性,如抗菌、抗病毒、抗腫瘤和免疫調節等功能,且不易產生耐藥性,其應用前景巨大[1-2]。防御素分布廣泛,如哺乳動物、昆蟲和植物等生物體內。根據二硫鍵連接方式的不同和半胱氨酸殘基位置的差異,將防御素分為α-防御素、β-防御素和θ-防御素三大類[3-4],其中β-防御素的抗菌活性高效、抗菌譜廣,如抗細菌、抗真菌、抗病毒和抗寄生蟲等,對一些耐藥性細菌也具有抑殺活性[5-6],但作用機制稍有差異,如hBD2可作用于結核分枝桿菌的細胞膜,而hBD3抑制葡萄球菌細胞壁的生成[7]。β-防御素的抗菌活性易受到離子種類和濃度的影響,如hBD2的殺白色念珠菌活性與鹽離子濃度關系密切[8],合成的bBD-1對鹽敏感,在低鹽離子濃度時,對乳腺炎大腸桿菌分離株有抑殺活性,但對乳腺炎葡萄球菌分離株的抑殺作用較弱[9]。
防御素專業網站(http://defensins.bii.astar.edu.sg/)收集了不同生物的防御素,共363種,其中牛源防御素共17種,均屬于β-防御素[10]。TAP是從牛氣管黏膜上皮中發現的牛的第一個防御素[11]。牛源防御素類抗菌肽可分布于體內不同組織部位,如TAP可分布于牛的氣管和肺泡巨噬細胞,BNBD5可分布于中性粒細胞、氣管、肺泡巨噬細胞,bBD可分布于牛的乳頭黏膜、腎、陰道、卵巢、結腸等[12]。不同的牛源防御素類抗菌肽有著不同的抗微生物活性,如BNBD4和BNBD5對大腸桿菌和金黃色葡萄球菌具有較好的抑殺活性[13],TAP對金黃色葡萄球菌、大腸桿菌、銅綠假單胞菌、肺炎克雷伯菌、白色念珠菌具有較好的抑殺活性[12],LAP對金黃色葡萄球菌、大腸桿菌、銅綠假單胞菌和白色念珠菌具有較好的抑殺活性[14]。
本研究利用在線生物信息學軟件對牛源防御素類抗菌肽的理化性質、二級結構、磷酸化和糖基化位點等特征進行了分析,結合已知牛源防御素類抗菌肽的生物學活性,初步分析牛源防御素類抗菌肽的作用機制,為深入研究與應用牛源防御素類抗菌肽提供更多的信息基礎資料。
1 材料與方法
1. 1 抗菌肽氨基酸序列 牛源防御素類抗菌肽的氨基酸序列主要來自于文獻[10],其序列如表1所示。

表1 牛源防御素類抗菌肽的氨基酸序列Table 1 Amino acid sequence of AMPs in bovine defensins family
1.2 生物信息學分析方法 利用ProtParam進行理化性質的分析,利用SOMA進行二級結構分析,利用NetOGlyc 4.0 Server進行糖基化分析,利用NetPhos3.1Server進行磷酸化分析,利用Target P 1.1 Server進行亞細胞內定位。
2 結果與分析
2.1 牛源防御素的理化性質 從表2中可以看出,牛源防御素類抗菌肽由37~42個氨基酸殘基構成;除了LAP、BNBD2、BNBD3、BNBD4和BNBD6的等電點大于10之外,其余牛源防御素類抗菌肽的等電點均大于9且小于或等于10(除BNBD1的等電點小于9),但仍然能表明這些抗菌肽應帶正電荷;通過分析牛源防御素類抗菌肽的帶電荷氨基酸數目,發現帶正電荷氨基酸數目較多,最多的達10個正電荷氨基酸殘基,而帶負電荷的氨基酸殘基數較少乃至缺乏,總體而言,牛源防御素類抗菌肽為帶正電荷的多肽(即陽離子型抗菌肽)。
從表3中可以看出,牛源防御素類抗菌肽均可以通過紫外方法進行檢測;對半衰期進行分析,發現EBD、BNBD2、BNBD8、BNBD11和BNBD12的半衰期相對較長,其他牛源防御素類抗菌肽的半衰期相對較短;對穩定性進行分析,發現TAP、bBD-1、BNBD2、BNBD3、BNBD7、BNBD8和BNBD9為穩定性多肽,其他牛源防御素類抗菌肽為不穩定多肽;對脂溶性指數進行分析,EBD的指數值最高(>80),應具有較好的耐熱性能,BNBD10的指數值次之(>75),而其他牛源防御素類抗菌肽的指數值稍低,耐熱性能應稍差;BNBD12和BNBD13的GRAVY值>0,表明其為疏水性多肽,而其他牛源防御素類抗菌肽的GRAVY值<0,表明其為親水性多肽。BNBD7、BNBD9和BNBD10含有E(谷氨酸),僅BNBD1、BNBD2、BNBD3、BNBD6、BNBD7、BNBD8和BNBD9含有H(組氨酸),僅TAP、BNBD2、BNBD3、BNBD5和BNBD6不含有L(亮氨酸),僅BNBD4和BNBD5不含有K(賴氨酸),僅 EBD、bBD-1、BNBD2、BNBD3、BNBD6、BNBD7、BNBD8和BNBD9不含有M(甲硫氨酸),僅TAP、LAP、bBD-1和BNBD10不含有F(苯丙氨酸),僅BNBD6、BNBD7、BNBD8和BNBD9不含有S(絲氨酸),僅TAP、LAP、bBD-1、BNBD7、BNBD8和BNBD9不含有W(色氨酸),僅bBD-1、BNBD6和BNBD10含有Y(酪氨酸),僅bBD-1不含有V(纈氨酸)。
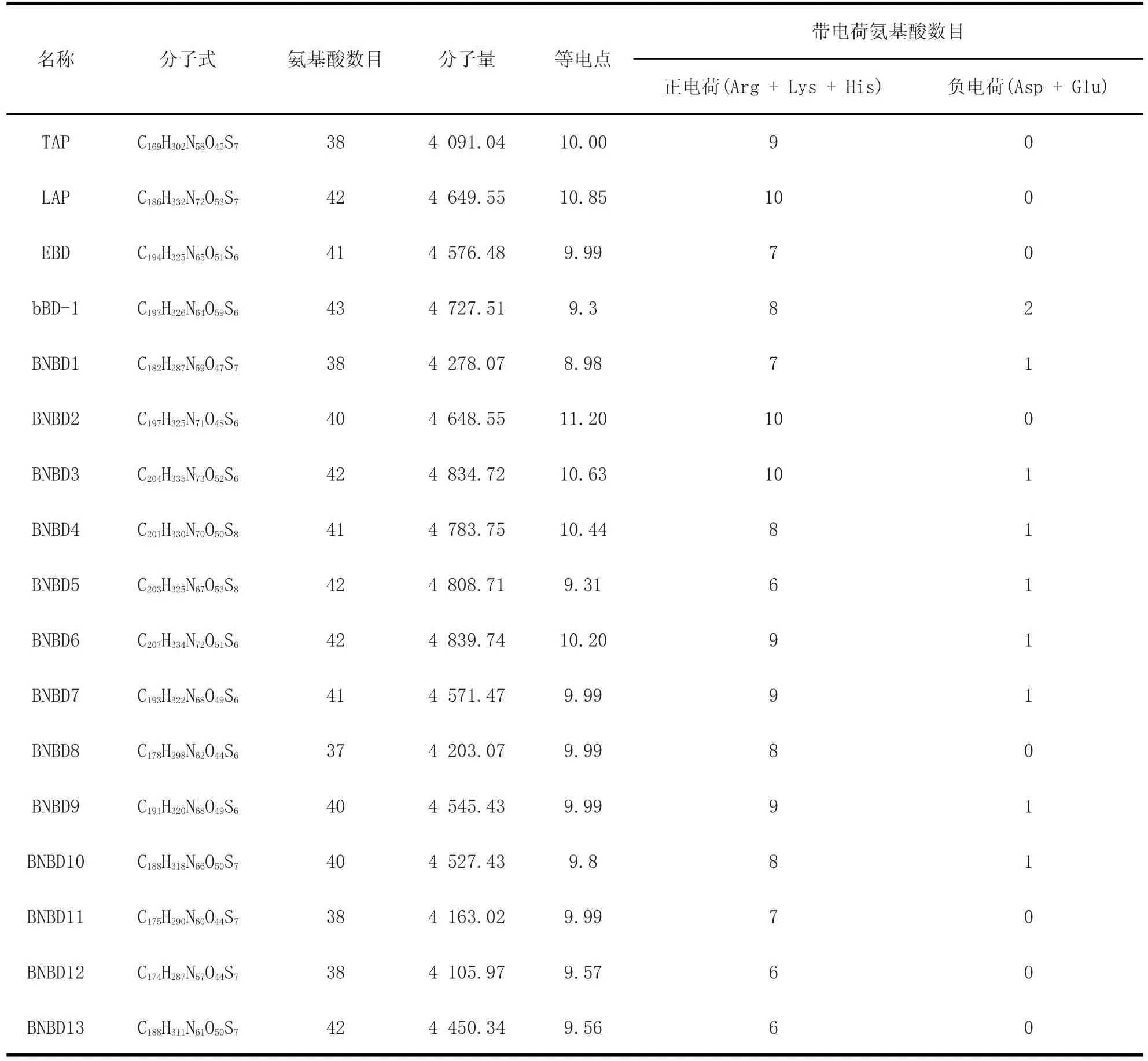
表2 牛源防御素的氨基酸數目、等電點、分子質量、帶電氨基酸數目Table 2 Total number of amino acids,pI,MW and number of charged amino acids of bovien defensins
表4中可以看出,牛源防御素類抗菌肽TAP、LAP、EBD、bBD-1、BNBD1、BNBD2、BNBD3、BNBD4、BNBD5、 BNBD6、 BNBD7、 BNBD8、 BNBD9、 BNBD10、BNBD11、BNBD12和BNBD13分別由13、14、14、15、18、14、15、15、14、15、14、13、14、17、15、15和15種不同的氨基酸殘基構成,均含有的氨基酸包括R(精氨酸)、N(天冬酰胺)、C(半胱氨酸)、Q(谷氨酰胺)、G(甘氨酸)、I(異亮氨酸)、P(脯氨酸)和T(酪氨酸),僅TAP、LAP、bBD-1、BNBD1和BNBD10含有A(丙氨酸),僅bBD-1和BNBD1含有D(天冬氨酸),僅 bBD-1、BNBD3、BNBD4、BNBD5、BNBD6、

表 3 牛源防御素的紫外檢測、半衰期、穩定性、脂溶性和親水性等信息Table 3 Detection with UV,half life,stability,index of fatty amino acids and hydrophily of bovine defensins
2.2 牛源防御素的二級結構 牛源防御素的二級結構預測(參數:視窗寬度為17,相似度閾值為8,狀態數目為4)結果如表5所示。從表5中可以看出,TAP、LAP、bBD-1、BNBD2、BNBD3、BNBD4、BNBD6、BNBD7、BNBD8和BNBD10由α螺旋、β折疊、β轉角和無規則卷曲構成,EBD、BNBD1、BNBD5、BNBD11、BNBD12和BNBD13由β折疊、β轉角和無規則卷曲構成;在17種牛源防御素類抗菌肽中,BNBD10的α螺旋含量最高,BNBD8的β折疊含量最高,LAP的β轉角含量最高,EBD的無規則卷曲含量最高。
2.3 牛源防御素的細胞內定位 經在線軟件分析,牛源防御素EBD、BNBD2、BNBD4、BNBD8、BNBD11定位于線粒體,而其他防御素可能定位于細胞外。
2.4 牛源防御素的糖基化和磷酸化位點 經在線軟件分析,牛源防御素類抗菌肽不存在糖基化。從表6中可以看出,除BNBD7不存在磷酸化位點外,其他16種防御素類抗菌肽存在絲氨酸和蘇氨酸的磷酸化位點,但不存在酪氨酸的磷酸化位點;不同的防御素有不同數目的磷酸化位點,最高的如bBD-1有6個位點的磷酸化,EBD次之,其他的有1~3個磷酸化位點;有的位點可被多種激酶磷酸化,如防御素LAP的第6位點的絲氨酸(S)可被unsp、PKA、DNAPK和ATM等激酶磷酸化。

表 4 牛源防御素的氨基酸組成Table 4 Amino acid composition of bovine defensins

表 5 牛源防御素的二級結構Table 5 Secondary structure of bovine defensins
3 討論
絕大多數抗菌肽是陽離子型的,其抗菌作用與帶正電荷的氨基酸殘基有關。抗菌肽的帶正電荷氨基酸殘基與帶負電荷的細菌細胞膜相結合,然后通過不同的作用機制(如桶板模型、地毯模型、蟲孔模型和凝聚模型等)發揮抑菌或殺菌活性[15-16]。精氨酸、賴氨酸和組氨酸等帶正電荷的氨基酸與抗菌肽的抗菌活性有密切關系[17-18]。牛源防御素類抗菌肽的一級結構中存在較多的精氨酸、賴氨酸和組氨酸等堿性氨基酸,使其具有5~10個凈正電荷,這應是牛源防御素類抗菌肽發揮抑菌或殺菌活性的重要基礎。
不同抗菌肽的二級結構是不同的[12]。例如,抗菌肽LK為α螺旋形[19],抗菌肽HNP-2為β折疊型[20]。抗菌肽的二級結構主要由α螺旋、β折疊、β轉角和無規則卷曲等構成。在牛源防御素類抗菌肽的二級結構中,無規則卷曲結構所占比例最高。而無規則卷曲結構有利于抗菌肽的穩定性。Lee等[21]認為這種穩定性與甘氨酸有關。牛源防御素類抗菌肽的氨基酸序列中均含有一定比例的甘氨酸。因此,可以認為甘氨酸對牛源防御素類抗菌肽的穩定性具有重要作用。
不同抗菌肽的亞細胞定位是不同的[22]。例如,雞內源性抗菌肽NK-lysin定位于細胞質內[23],雞內源性抗菌肽LEAP-2定位于細胞核內[24]。經分析,牛源防御素類抗菌肽僅少數定位于線粒體,大多數定位于細胞外。對于定位于細胞外的牛源防御素類抗菌肽,可能是其前原肽在信號肽的引導下到達細胞外等靶標部位,在某些酶的作用下,產生抗菌肽的成熟肽,從而對細胞外的病原微生物或腫瘤細胞發揮拮抗作用,或與其受體結合而發揮免疫調節作用。對于定位于線粒體的牛源防御素類抗菌肽,可能其對線粒體等細胞器具有一定作用,但究竟是何種作用還值得深入探討。
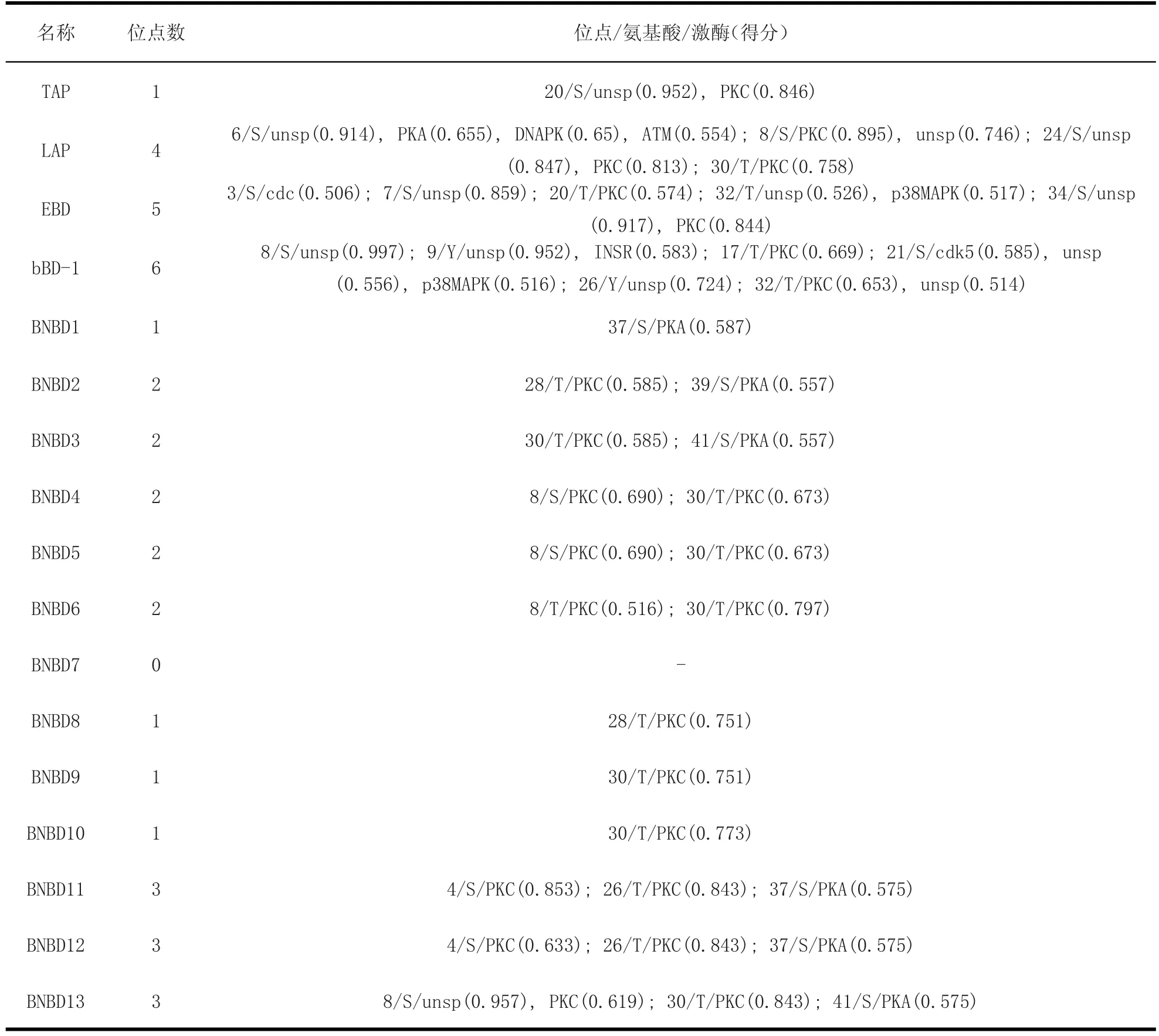
表 6 牛源防御素的的磷酸化位點Table 6 Phosphorylation site of bovine defensins
蛋白磷酸化在細胞分化、凋亡、信號傳遞和免疫等生物學過程中具有調節作用[25]。肌紅蛋白發生磷酸化修飾后,二級結構發生改變,穩定性降低[26],但C/EBPβ蛋白的磷酸化修飾能增強其在3T3-L1前脂肪細胞成脂分化過程中的穩定性[27]。經預測發現,除了BNBD7外,牛源防御素類抗菌肽存在不同數目的絲氨酸和蘇氨酸的磷酸化位點。可以推測,牛源防御素類抗菌肽發生磷酸化后,其二級結構會發生相應改變,但這種改變對牛源防御素類抗菌肽的穩定性、抗菌活性等生物學功能有何具體影響還值得深入研究。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
