青海省海晏縣線(xiàn)葉嵩草內(nèi)生細(xì)菌的生物功能鑒定及測(cè)定
2018-08-23 11:53:00魏立娟
草地學(xué)報(bào) 2018年3期
關(guān)鍵詞:植物
魏立娟, , *,
(1.甘肅農(nóng)業(yè)大學(xué),甘肅 蘭州730070; 2.青海大學(xué), 青海 西寧810003)
植物內(nèi)生細(xì)菌是指能定殖在健康植物組織內(nèi)并與植物建立和諧關(guān)系的一類(lèi)微生物[1-2],隨著植物微生態(tài)系統(tǒng)研究的不斷深入,內(nèi)生細(xì)菌的存在和作用已得到廣泛共識(shí)[3]。幾乎所有的植物中都伴隨植物內(nèi)生細(xì)菌的存在,其長(zhǎng)期生活于植物體內(nèi)的特殊環(huán)境中[4-5],內(nèi)生細(xì)菌可利用風(fēng)、土壤顆粒、水、農(nóng)業(yè)器具等多種外力條件和人類(lèi)、鳥(niǎo)、昆蟲(chóng)、線(xiàn)蟲(chóng)等多種媒介從根際土壤定殖在寄主植物根內(nèi)部,具有溶磷、產(chǎn)生植物激素、固氮、合成鐵載體、誘導(dǎo)植物產(chǎn)生抗性、產(chǎn)生抗真菌代謝產(chǎn)物等多種生物功能[3]。王娜等[6]將內(nèi)生細(xì)菌接入棉花苗后再接種黃萎病菌,對(duì)黃萎病菌的防治效果可達(dá)79.52%~89.79%,有助于棉花病害的防治,為棉花增產(chǎn)提供有力幫助;杜曉寧等[7]研究表明枸杞可培養(yǎng)內(nèi)生細(xì)菌遺傳多樣性豐富,對(duì)植物病原菌有較強(qiáng)的抑制活性;蔡學(xué)清等[8]得出內(nèi)生細(xì)菌在寄主植物的定殖情況與其保鮮效果之間存在一定的正相關(guān)性,即內(nèi)生定殖是其達(dá)到保鮮效應(yīng)的機(jī)制之一。植物內(nèi)生細(xì)菌也是重要的微生物農(nóng)藥來(lái)源[9],目前在生物防治中具有很大潛力,也作為增產(chǎn)菌或潛在的生防載體菌而加以利用。
線(xiàn)葉嵩草(Kobresiacapillifolia) 是高寒草地牧區(qū)的優(yōu)勢(shì)牧草,不僅能適應(yīng)惡劣的環(huán)境,還能為家畜提供優(yōu)質(zhì)的營(yíng)養(yǎng)物質(zhì)。崔月貞等[10]與高曉星等[11]對(duì)東祁連山牧草內(nèi)生細(xì)菌研究表明,其具有溶磷、固氮、產(chǎn)吲哚乙酸(indole-3-acetic acid,IAA)作用。本試驗(yàn)對(duì)青藏高原青海省線(xiàn)葉嵩草中內(nèi)生細(xì)菌的特性及其抗菌活性進(jìn)行研究,以期其同樣具有溶磷、固氮、產(chǎn)IAA等能力,本研究以期明確青藏高原高寒草地線(xiàn)葉嵩草內(nèi)生細(xì)菌的多樣性和生物功能,為合理利用極端生境牧草提供依據(jù)。
1 材料與方法
1.1 材料
1.1.1供試菌株 線(xiàn)葉嵩草采于2016年8月37°4′8″ N,100°52′42″ E的青海省海晏縣青海湖鄉(xiāng),并對(duì)其根部進(jìn)行分離得到內(nèi)生細(xì)菌。
1.1.2供試指示菌 馬鈴薯炭疽病菌(Colletotrlchumcoccodes),馬鈴薯枯萎病菌(Fusarlumavenaceum),馬鈴薯壞疽病菌(Phomafoveata)和番茄早疫病菌(Alternariasolani),由甘肅農(nóng)業(yè)大學(xué)植物保護(hù)學(xué)院植物病原細(xì)菌及細(xì)菌多樣性實(shí)驗(yàn)室提供。
1.1.3供試培養(yǎng)基 肉汁胨培養(yǎng)基(nutrient Ager,NA)用于內(nèi)生細(xì)菌分離、純化和保存等[12];馬鈴薯葡萄糖培養(yǎng)基(potato Dextrose Agar,PDA)用于植物病原真菌的培養(yǎng)和對(duì)峙實(shí)驗(yàn)[13];金氏(King)培養(yǎng)基用于產(chǎn)IAA的試驗(yàn);Pikovaskaia培養(yǎng)基(pikovaskaia’s,PKO)和蒙金娜培養(yǎng)基(Mehknha)分別用于溶解無(wú)機(jī)磷和有機(jī)磷試驗(yàn)[14],阿須貝無(wú)氮培養(yǎng)基(Ashby)用于篩選固氮菌[15]。
1.2 方法
1.2.1內(nèi)生細(xì)菌的分離純化 將線(xiàn)葉嵩草根部組織表面用清水清洗干凈,晾干后稱(chēng)取2 g,表面消毒后置于無(wú)菌研缽中研碎,吸取植物組織研磨浸出液按10倍濃度梯度稀釋后,吸取0.1 mL涂布于NA培養(yǎng)基上,以最后一次洗滌水為對(duì)照,28℃培養(yǎng)3~7 d,根據(jù)菌落形態(tài),顏色,大小,邊緣整齊度及表面形態(tài)等分類(lèi),純化后4℃保存?zhèn)溆肹16]。
1.2.2生物功能測(cè)定
1.2.2.1 拮抗能力測(cè)定
采用平板對(duì)峙法測(cè)定抑菌效果[17-19],用直徑5 mm的打孔器取指示菌菌塊于PDA平板中央,在指示菌四周3 cm處接種供試菌,4次重復(fù),將馬鈴薯壞疽病菌放于15℃培養(yǎng),其余3個(gè)指示菌放于28℃培養(yǎng)。以不接種供試內(nèi)生菌為對(duì)照,待對(duì)照滿(mǎn)皿后測(cè)量菌落直徑,計(jì)算抑菌率[20]。
1.2.2.2 固氮能力測(cè)定
將純化得到的內(nèi)生細(xì)菌接種于NA液體培養(yǎng)基中,置于搖床28℃、120 r·min-1震蕩培養(yǎng)24 h,取培養(yǎng)所得到的菌懸液0.1 mL接種于阿須貝無(wú)氮平板和液體培養(yǎng)基中,以無(wú)菌水為對(duì)照,3次重復(fù),平板置于28℃培養(yǎng)箱培養(yǎng),液體置于120 r·min-1震蕩培養(yǎng),待第7天觀察,平板上有菌落或液體培養(yǎng)基變渾濁記為“+”,否則記為“-”。
1.2.2.3 溶磷能力測(cè)定
將活化的內(nèi)生細(xì)菌涂布于PKO(無(wú)機(jī)磷)和蒙金娜(有機(jī)磷)培養(yǎng)基上,28℃培養(yǎng)14天后觀察解磷圈,3次重復(fù),根據(jù)解磷圈與菌落直徑比值大小確定其溶磷能力。
1.2.2.4 產(chǎn)IAA能力測(cè)定
采用Salkowski比色法[21],分別在100 mg·L-1含色氨酸和不含色氨酸的金氏培養(yǎng)液中接種0.1 mL菌懸液[13],加等量無(wú)菌水為對(duì)照,置于28℃,120 r·min-1恒溫震蕩培養(yǎng)12 d,取50 μL菌懸液加入等量PC比色液,室溫靜置15 min后,觀察顯色反應(yīng),3次重復(fù),全部變紅表明能產(chǎn)IAA,記為“+”,否則記為“-”。
1.2.3內(nèi)生細(xì)菌的鑒定
1.2.3.1 培養(yǎng)性狀觀察
將內(nèi)生細(xì)菌接種于NA培養(yǎng)基上,在28℃無(wú)光照條件下培養(yǎng)72 h,觀察內(nèi)生細(xì)菌菌落形態(tài)特征并描述和照相。
1.2.3.2 形態(tài)觀察
內(nèi)生細(xì)菌在NA平板上培養(yǎng)經(jīng)18~24 h革蘭氏染色和鏡檢,觀察菌體顏色和形態(tài),并顯微拍照。
1.2.3.3 16S rDNA鑒定
以10株內(nèi)生細(xì)菌DNA為模板,使用細(xì)菌16S rRNA 基因通用引物進(jìn)行擴(kuò)增,擴(kuò)增引物1為5'-CCG GAT CCA GAG TTT GAT CAT GGC TCA GCA-3',引物2為5'-CGG GAT CCT ACG GCT ACC TTG TTA CGA CTT-3',擴(kuò)增體系包括25 μL 2×MasterMix,模板 2 μL,上下游引物各2 μL,ddH2O 19 μL,總體積50 μL,PCR程序?yàn)椋?4℃預(yù)變性 4 min, 94℃變性30 s, 60℃退火30 s, 72℃延伸90 s,30個(gè)循環(huán),72℃延伸10 min。獲得特異片段送到武漢金開(kāi)瑞生物工程有限公司測(cè)序,并將序列上傳至NCBI 進(jìn)行BLAST比對(duì)分析(https://blast.ncbi.nlm.nih.gov/Blast.cgi)。用Maga 6.0軟件構(gòu)建系統(tǒng)發(fā)育樹(shù),確定其系統(tǒng)發(fā)育學(xué)地位。
2 結(jié)果與分析
2.1 內(nèi)生細(xì)菌的分離純化
根據(jù)菌落大小、顏色、形態(tài)挑取不同單菌落,純化后從線(xiàn)葉嵩草根部分離出10株內(nèi)生細(xì)菌,分別編號(hào)為2G1、2G2、2G3、2G4、2G5、2G6、2G7、2G8、2G9和2G10保存?zhèn)溆谩?duì)照沒(méi)有細(xì)菌菌落長(zhǎng)出,說(shuō)明表面消毒徹底,所得細(xì)菌均為內(nèi)生細(xì)菌。
2.2 生物功能測(cè)定
除2G1和2G10對(duì)4種指示菌均無(wú)拮抗作用外,其他8株內(nèi)生細(xì)菌對(duì)4種指示菌均有拮抗作用,其中對(duì)馬鈴薯炭疽病菌的拮抗作用最強(qiáng),最強(qiáng)菌株是2G6,抑菌率達(dá) 88.58%, 2G8抑菌率最弱,也達(dá)66.98%,都顯著高于對(duì)其他菌株的抑菌率(P<0.05)。8株內(nèi)生細(xì)菌對(duì)番茄早疫病菌、馬鈴薯枯萎病菌和馬鈴薯壞疽病菌抑菌率介于32.59%~63.61%之間,其中2G5對(duì)番茄早疫病菌的抑菌率也較高為63.61%,(圖1,表1)。該結(jié)果表明,除2G1和2G10以外的8株內(nèi)生細(xì)菌均具有抑菌能力,具有良好的生物防治應(yīng)用潛力。

圖1 2G9的抑菌作用Fig.1 The antagonism effect of 2G9
10株內(nèi)生細(xì)菌均能固氮,但無(wú)溶磷能力,2G1和2G10分泌IAA(表1)。該結(jié)果說(shuō)明線(xiàn)葉嵩草根部?jī)?nèi)生細(xì)菌具有功能多樣性。
2.3 內(nèi)生細(xì)菌的鑒定
2.3.1培養(yǎng)形狀觀察 從10株內(nèi)生細(xì)菌培養(yǎng)性狀看線(xiàn)葉嵩草內(nèi)生細(xì)菌在形態(tài)上存在多樣性(圖2)。
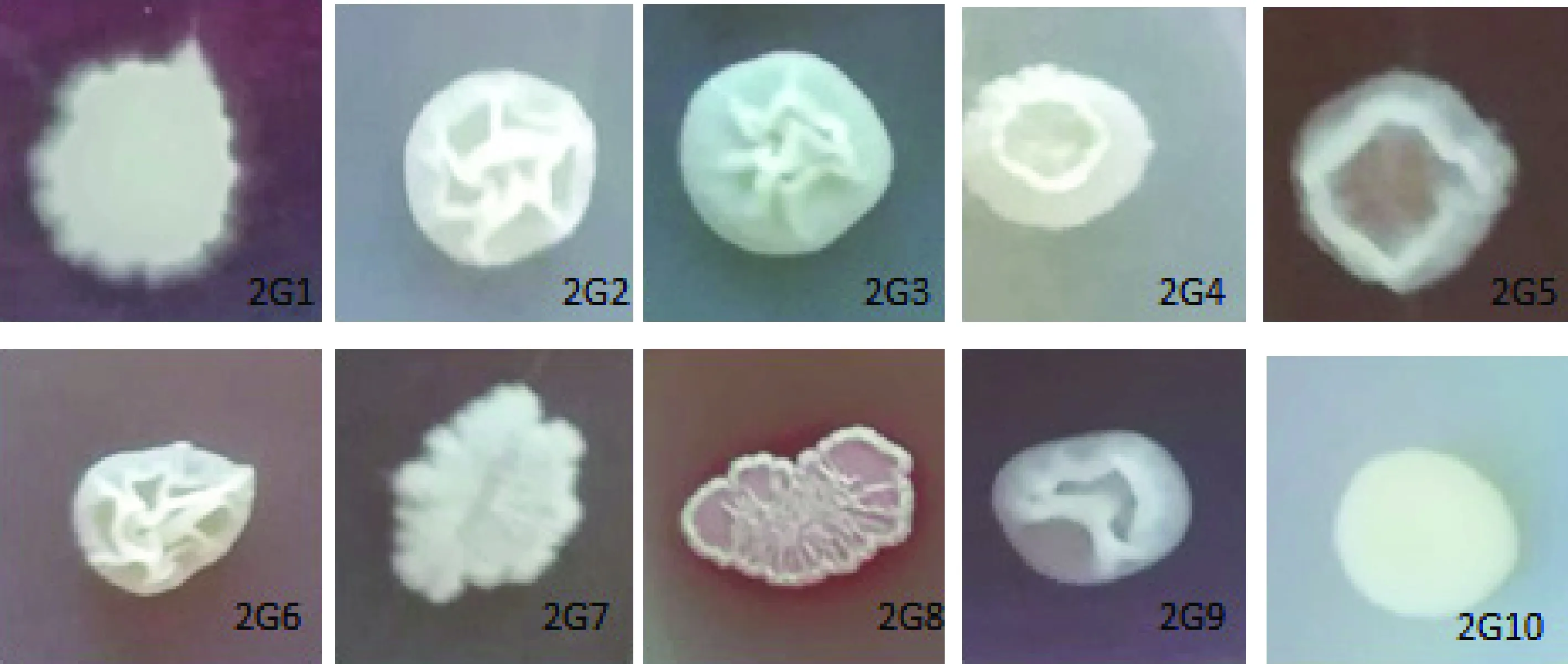
圖2 內(nèi)生細(xì)菌培養(yǎng)性狀Fig.2 Endophytic bacteria culture traits

表1 線(xiàn)葉嵩草內(nèi)生細(xì)菌的生物功能測(cè)定Table 1 Determination of biological functions of endophyte bacterium from Kobreais capillifolia
注:“+”:有功能;“-”無(wú)功能;同一行標(biāo)不同字母表示在0.05水平差異顯著
Note: “+”:With biological function;“-”:Without biological function; Different lowercase letters within the same line mean significant difference among different treatments at the 0.05 level
2.3.2形態(tài)觀察 10株內(nèi)生細(xì)菌均為革蘭氏陽(yáng)性菌,且均為桿狀,菌體差別明顯,2G10菌體最長(zhǎng),達(dá)20.29 μm,2G1菌體最寬,為1.60 μm,其他菌株長(zhǎng)和寬均介于這兩者之間(圖3,表2),表現(xiàn)出明顯的形態(tài)多樣性。

圖3 內(nèi)生細(xì)菌的革蘭氏染色Fig.3 Gram stain of endophytice bacteria from A.inebrians

表2 內(nèi)生細(xì)菌形態(tài)特征Table 2 Morphological characteristics of endophytic bacteria
注:G+:革蘭氏陽(yáng)性
Note: G+: Gram-positive bacteria
2.3.316S rDNA鑒定 所測(cè)序列與GenBank數(shù)據(jù)庫(kù)中序列比對(duì)和構(gòu)建系統(tǒng)發(fā)育樹(shù)表明,10株內(nèi)生細(xì)菌都屬于芽孢桿菌屬,2G1和2G10分別與芽孢桿菌屬的Bacillusthuringiensis(KT965084.1)、Bacillussubtilis(JX994100.1)和Bacillussimpix(KF818620.1)一致性在99%以上,并且在系統(tǒng)發(fā)育樹(shù)上聚在一起,初步將其分別鑒定為芽孢桿菌屬的Bacillusthuringiensis和Bacillussimpix,2G2和2G9分別與Bacillusmethylotrophicus(KU877329.1、KM817258.1)的一致性在99%以上,在系統(tǒng)發(fā)育樹(shù)上聚在一起,初步鑒定為Bacillusmethylotrophicus,2G3和2G6分別與Bacillusamyloliquefaciens(KU321524.1)的一致性在99%以上,并在系統(tǒng)發(fā)育樹(shù)上聚在一起,初步鑒定為Bacillusamyloliquefaciens,2G4和2G5分別與Bacillussiamensis(KC851838.1)的一致性達(dá)99%,并且在系統(tǒng)發(fā)育樹(shù)上聚在一起,初步鑒定為Bacillussiamensis,2G7和2G8分別Bacillussubtilis.(KU551205.1、KU5563 29.1)一致性達(dá)99%以上,并且在系統(tǒng)發(fā)育樹(shù)上聚在一起,初步鑒定為Bacillussubtilis(圖3)。結(jié)合前文中的培養(yǎng)性狀和形態(tài)特征測(cè)定,將10株內(nèi)生細(xì)菌分屬于1個(gè)屬6個(gè)種,得出線(xiàn)葉嵩草根部?jī)?nèi)生細(xì)菌在屬的種類(lèi)上比較單一,但在種的水平上有很大差別。

3討論與結(jié)論
廣泛分布于青藏高原的高山草甸對(duì)密集放牧非常寬容,這些草地中的植物群落目前正在從莎草科向禾本科物種轉(zhuǎn)變,因此,正在進(jìn)行的植物群落禾草替代將減少青藏高原矮嵩草甸[22],減少青藏高原菌種資源。植物內(nèi)生菌的生境特殊性決定了其既有理論研究的廣度和深度,又有廣泛的應(yīng)用潛力,是潛力巨大且尚待開(kāi)發(fā)的微生物新資源[23]。Forchetti等[24]和Zhang等[25]認(rèn)為具有固氮和分泌IAA能力的植物內(nèi)生細(xì)菌開(kāi)發(fā)為生物菌肥有優(yōu)勢(shì),本試驗(yàn)分離得到的2G1和2G10同時(shí)具有固氮和分泌IAA的能力,能夠作為生物菌肥開(kāi)發(fā)。崔月貞等[10]得出高寒草地優(yōu)勢(shì)牧草組織內(nèi)具有豐富的可分泌IAA的內(nèi)生細(xì)菌,且其中分泌IAA能力較強(qiáng)的菌株主要分布于線(xiàn)葉嵩草葉部和生長(zhǎng)期的根部,但本試驗(yàn)分離出的內(nèi)生細(xì)菌僅有2株具分泌IAA的能力,可能與不同地區(qū)線(xiàn)葉嵩草內(nèi)生細(xì)菌的差異有關(guān),但具體原因仍要進(jìn)一步研究。
馬鈴薯是我國(guó)較為重要的糧飼兼用作物,但是近年來(lái)報(bào)道的馬鈴薯病害日趨嚴(yán)重[26],本試驗(yàn)分離所得內(nèi)生細(xì)菌除2G1和2G10以外,其余8株內(nèi)生細(xì)菌對(duì)馬鈴薯壞疽病菌、馬鈴薯炭疽病菌、馬鈴薯枯萎病菌和番茄早疫病菌都有較好的抑制作用,其中對(duì)馬鈴薯炭疽病的抑制效果最強(qiáng),抑菌率分別為84.88%、84.57%、83.64%、85.80%、88.58%、86.11%、66.98%和82.41%,比崔月貞等[19]報(bào)道的最強(qiáng)抑菌率70.25%平均高13.07%,在馬鈴薯病害防治中展現(xiàn)出了巨大開(kāi)發(fā)為生物制劑的潛力,為該菌能夠更好的在大田中防治馬鈴薯炭疽病,還需進(jìn)一步研究其抑菌機(jī)制及發(fā)酵條件優(yōu)化等。此外本試驗(yàn)還發(fā)現(xiàn)2G2、2G3、2G4、2G5、2G6、2G7、2G8和2G9具有固氮能力,該研究結(jié)果為開(kāi)發(fā)藥肥兼用的微生物產(chǎn)品提供了菌種資源。
利用16S rDNA序列分析方法對(duì)內(nèi)生細(xì)菌進(jìn)行鑒定得出,分離所得10株內(nèi)生細(xì)菌均為芽孢桿菌屬,與張芳芳等[27]研究所得結(jié)果芽孢桿菌屬是分離頻率最高的菌種相似。試驗(yàn)中分離菌株均為芽孢桿菌屬的原因可能在于青海溫度低,其它菌株可能不能越冬,而芽孢桿菌抗逆能力強(qiáng)。此外本試驗(yàn)僅利用16S rDNA序列分析方法對(duì)內(nèi)生細(xì)菌進(jìn)行了鑒定,還需利用生理生化試驗(yàn)及(G+C)%等進(jìn)一步鑒定。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
