番鴨Viperin基因的克隆表達及其抗MDRV的作用研究
2018-08-01 02:03:10陳潔瀅孫亞楠邵文涵池曉娟陳吉龍
中國獸醫雜志 2018年4期
陳潔瀅, 孫亞楠, 邵文涵, 池曉娟, 陳吉龍, 王 松
(福建農林大學動物科學學院, 福建 福州 350002)
番鴨呼腸孤病毒病是由番鴨呼腸孤病毒(MDRV)引起的一種以腳軟、肝脾有大量白色壞死點、高發病率和高病死率為特征的新的急性傳染病,對番鴨養殖業造成極為重大的經濟損失。機體天然免疫系統是抗病毒感染的第一道防線,病毒感染宿主后,天然免疫系統會被激活,經細胞中一系列的信號傳導,誘導宿主分泌多種抑制病毒的細胞因子。干擾素(Interferon,IFN)是最重要的細胞因子之一,在抵抗病毒的過程中發揮著重要作用,干擾素可以與細胞表面的特異性受體結合,進而誘導上千種干擾素刺激基因(IFN-stimulated genes, ISGs)的表達[1]。MDRV感染后能迅速激活宿主天然免疫信號通路,誘導IFNs及某些重要ISGs的大量表達[2]。從ISGs被發現以來,國內外學者不斷探索ISGs在病毒侵襲宿主過程中發揮的作用及作用機理,目前對包括Viperin在內的少數ISGs的抗病毒機制有較深入的了解[3]。
Viperin,也被稱為cig5和RSAD2,是一種重要的ISGs,是與內質網相關的抗病毒蛋白,可被干擾素、細菌脂多糖(LPS)、poly ( I ∶ C) 和多種病毒等誘導表達,具有廣譜抗病毒活性[4],對淋巴細胞脈絡叢腦膜炎病毒(LCMV)[5]、丙型肝炎病毒(HCV)[6]、登革熱病毒(DENV)[7]、基孔肯雅病病毒(CHIKV)[8]、仙臺病病毒(SeV)[9]、A型流感病毒(IAV)[10]、日本腦炎病毒( JEV)[11]、人類免疫缺陷病毒(HIV)[12]和西尼羅河病病毒(WNV)[13]等多種病毒具有抑制作用。但目前還未有文獻報道番鴨Viperin蛋白在番鴨體內的表達情況,以及其對MDRV復制影響的相關研究。本研究首先檢測了番鴨Viperin基因在感染MDRV細胞和雛番鴨組織中的表達情況;然后,克隆番鴨Viperin 基因到真核表達質粒,并對Viperin在MDRV復制中的作用進行了研究。本研究首次證實番鴨Viperin蛋白具有抗MDRV的作用,為抗MDRV藥物的篩選和研制提供了新的選擇,也為番鴨Viperin 蛋白抗病毒活性的初步評價提供了依據。
1 材料
1.1 細胞、病毒、實驗動物和菌株 293T細胞,由本實驗室保存;番鴨鴨胚成纖維細胞(MDEF),分離自11~13日齡的番鴨胚;MDRV病毒YB株,由福建農林大學動物科學學院吳寶成老師惠贈;番鴨購自廣東溫氏食品集團有限公司莆田分公司;大腸桿菌DH5α感受態細胞,購自天根生化科技( 北京) 有限公司。
1.2 培養基及主要試劑 DMEM、胎牛血清,Gibco 公司生產; 核酸限制性內切酶EcoRⅠ、BamHⅠ、T4 DNA 連接酶、Pyrobest 高保真DNA 聚合酶、Taq酶、Oligo d( T) 18 Primer、Recombinant Dnase I、Cloned Ribonuclease Inhibitor、dNTP Mixture、核酸Marker,均由TaKaRa 公司生產;真核轉染試劑VigoFect,威格拉斯生物技術有限公司生產;NC膜,Millipore 公司生產;蛋白預染Marker,Thermo Scientific 公司生產;ECL 化學發光液,普利萊基因技術有限公司生產;Flag 標簽抗體、過硫酸銨和十二烷基磺酸鈉,均由Sigma 公司生產; 羊抗鼠二抗,Santa Cruz 公司生產;丙烯酰胺,國藥集團化學試劑有限公司生產。
1.3 主要儀器 細胞恒溫培養箱(型號為MCO175),SANYO 公司生產;生物安全柜(型號為Biobase BSC-1100IIA2-X),BioBase 公司生產;倒置顯微鏡(型號為Ti-E),Nikon公司生產;普通離心機(型號為Micro-17 )、冷凍離心機(型號為Theromo Fisher labofuge 400R),均由Thermo Scientific公司生產;低溫離心機(型號為Eppendorf 5424R)由Eppendorf公司生產;普通PCR儀(型號為T100)、凝膠成像系統(型號為GeIDoc XR + ),均由Bio-Rad公司生產;熒光定量PCR儀(LightCycler?96),由羅氏公司生產;化學發光和多色熒光成像系統(FluorChem M),由美國ProteinSimple公司生產;水平電泳槽、電泳儀(型號為BG-MIDI),均由北京百晶生物技術有限公司生產;隔水式電熱恒溫培養箱(型號為GRP-9080),上海培因實驗儀器有限公司生產;恒溫搖床(型號為ZWY-240),太倉市科教器材廠華利達實驗設備公司生產。
2 方法
2.1 引物的設計 從NCBI上查找目的基因mRNA序列,由于沒有番鴨序列,所以根據綠頭鴨Viperin (登錄號為KP198582.1) mRNA序列設計克隆引物1對,在5′端和3′端分別引入EcoRⅠ(GAATTC)和BamHⅠ(GGATCC) 酶切位點;再設計RT-PCR 引物1對。根據MDRV的結構蛋白P10 (登錄號為 EF547375.1) mRNA 序列設計RT-PCR引物1對。根據人GAPDH (登錄號為KJ891221.1) mRNA 序列設計RT-PCR 引物1對。根據番鴨actin(登錄號為 KY352479.1) mRNA序列設計RT-PCR 引物1對。
設計的引物序列見表1。

表1 引物序列信息
h:人源引物 ; d:鴨源引物
2.2 番鴨Viperin 編碼區序列的PCR擴增 從番鴨脾臟中提取總RNA,然后反轉錄獲得cDNA。以此cDNA 為模板,PCR擴增Viperin編碼區序列。PCR反應條件: 95 ℃預變性10 min; 95 ℃變性30 s,55 ℃退火30 s,72℃延伸40 s,共30 個循環;72 ℃再延伸10 min。1.0%瓊脂糖凝膠電泳跑膠,回收目的條帶。
2.3 真核表達載體的構建 將PCR擴增獲得的Viperin基因和pFlag-CMV-5a 載體分別經EcoRⅠ和BamHⅠ雙酶切后純化、回收,再用T4 DNA 連接酶將目的片段和載體的雙酶切產物在16 ℃條件下連接過夜。連接產物轉化大腸桿菌DH5α 感受態細胞,通過瓊脂糖凝膠電泳跑膠篩選重組質粒。
2.4 克隆質粒的鑒定EcoRⅠ和BamHⅠ雙酶切重組質粒,電泳鑒定;并將重組質粒送上海生工生物工程技術服務有限公司測序。將構建成功的含番鴨Viperin基因質粒命名為pFlag-CMV-Viperin。
2.5 真核表達質粒的轉染 待293T 細胞密度為80%~90%時,將舊培養基更換成轉染液( 無血清無雙抗的DMEM培養基),取3 μg pFlag-CMV-Viperin,加入轉染液中至總體積為50 μL,輕輕混勻;取VigoFect 2μL,加入轉染液中至總體積為50 μL,輕輕混勻,室溫放置5 min;將含有VigoFect的轉染液逐滴加入稀釋的DNA溶液中,輕輕混勻,室溫放置20 min;將混合液逐滴加入細胞中,輕輕搖勻,置于37 ℃、5% CO2培養箱中培養4 h后更換成完全培養基,繼續培養24 h,獲得過表達Viperin和空載體pFlag-CMV-5a的293T細胞。這兩種細胞感染同劑量的MDRV(MOI為1),收集感染24 h后的細胞,提取RNA,反轉錄獲得cDNA,用RT-PCR和qRT-PCR檢測細胞中Viperin的表達量和病毒基因P10的表達水平。
2.6 RT-PCR檢測Viperin基因的表達 提取細胞/組織RNA,并反轉錄獲得cDNA。以此cDNA 為模板,PCR擴增Viperin、P10和GAPDH /actin基因。PCR反應條件為:9 5℃預變性5 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸30 s,共30個循環,72 ℃再延伸10 min。以Actin為內參基因,1.2%瓊脂糖凝膠電泳跑膠,觀察目的條帶的灰度值,進而分析判斷目的基因mRNA 的表達情況。
2.7 Western-Blot檢測Viperin蛋白的表達 收獲細胞后提取細胞總蛋白。10% SDS-PAGE電泳分離目的蛋白并轉膜,用5%脫脂牛奶室溫封閉2 h。一抗用1×TBS 適當稀釋,室溫下搖動孵育2~4 h;二抗用3%脫脂牛奶稀釋,室溫下搖動孵育2~4 h。用1×TBS洗膜3次,10min/次;最后用化學發光顯色液沖洗膜5 min,曝光并分析結果。
3 結果與分析
3.1 MDRV感染能夠誘導Viperin高表達 MDRV感染293T細胞和MDEF細胞,分別感染不同時間后收集細胞樣品,提取RNA,檢測Viperin的變化情況。結果顯示,隨著病毒復制量的增多,Viperin表達量也呈上升的趨勢(圖1A、B)。此外,用MDRV感染5日齡的雛番鴨,總感染時間為4 d,每隔1 d處死番鴨并收取脾臟來檢測病毒P10和Viperin的表達量。結果顯示,番鴨脾臟中Viperin的表達量在MDRV感染后有明顯的升高,與細胞水平一致(圖1C)。由此說明Viperin的表達與MDRV的增殖可能有著一定的關系。

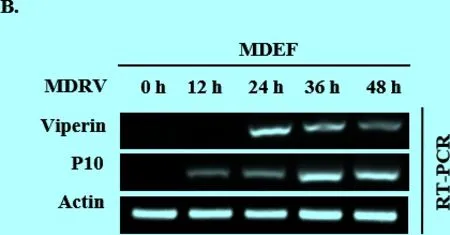
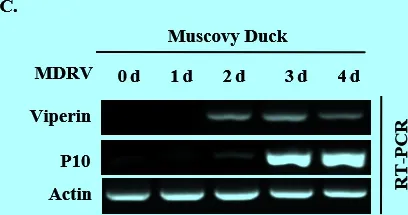
圖1MDRV感染293T、MDEF和番鴨后P10和Viperin表達量
3.2 重組質粒pFlag-CMV-Viperin的構建 為了進一步研究Viperin基因在MDRV感染機體過程中發揮的作用,我們構建了番鴨Viperin基因真核表達質粒:采用RT-PCR方法從番鴨脾臟組織中擴增得到1條長度約為1 000 bp的特異條帶(圖2),與預期片段大小相符。利用EcoR I 酶和BamH I 酶雙酶切PCR獲得的Viperin基因片段,同時用這2種酶雙酶切pFlag-CMV-5a載體,將回收的基因片段與載體片段連接,轉化后提取質粒,再利用限制性內切酶EcoR I和BamH I對質粒進行雙酶切鑒定,可以獲得預期約1 100 bp大小的目的片段(圖3)。最后,將該質粒送上海生工生物工程技術服務有限公司測序,測序得到的質粒基因序列與綠頭鴨Viperin的基因序列相似度為97.12%,說明重組質粒構建成功,將其命名為pFlag-CMV-Viperin。
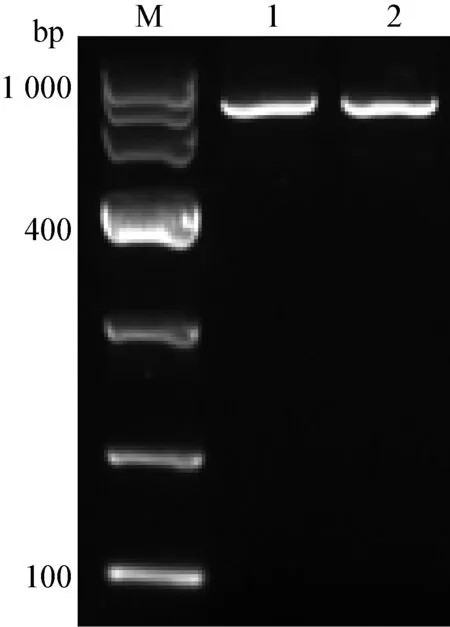
圖2 番鴨Viperin編碼區序列的PCR擴增
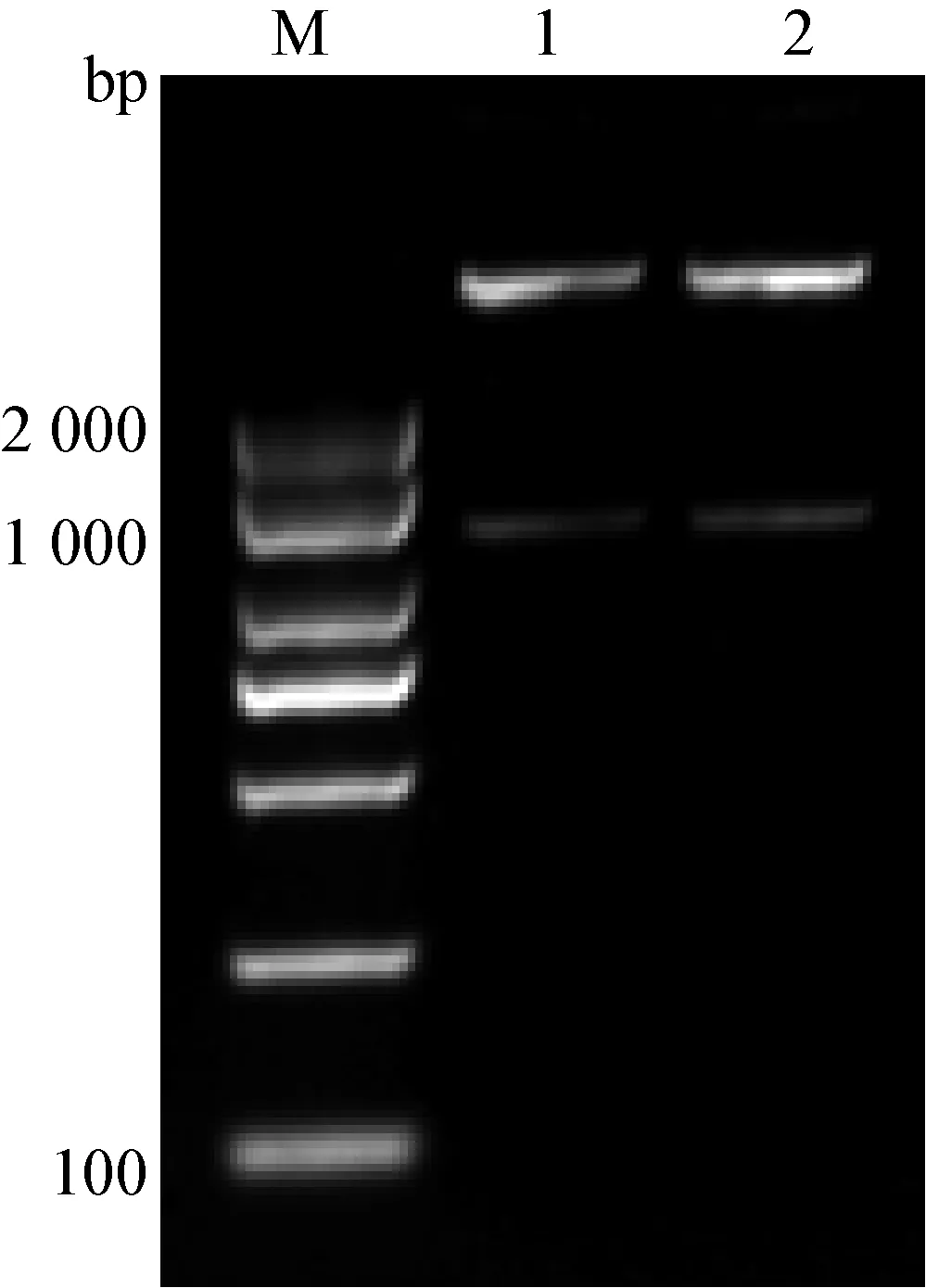
圖3 重組質粒pFlag-CMV-Muscovy-Viperin的酶切鑒定
3.3 重組質粒蛋白表達水平的檢測 由于番鴨鴨胚成纖維細胞無法傳代培養,且原代細胞轉染效率較低,因此選擇在293T模式細胞中檢測番鴨Viperin重組質粒的表達情況。pFlag-CMV-Viperin重組質粒轉染293T細胞24 h后收取細胞制備蛋白樣,Western-Blot 檢測Viperin蛋白表達水平。pFlag-CMV-Viperin帶有Flag標簽,因此一抗采用Flag標簽抗體。結果顯示,pFlag-CMV-Viperin重組質粒能夠在293T細胞中大量表達(圖4) 。
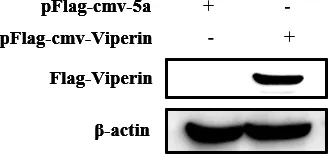
圖4 Viperin 蛋白表達水平的檢測
3.4 番鴨Viperin蛋白具有抗MDRV增殖的作用 基于成功構建了過表達番鴨Viperin的質粒,我們對其在MDRV復制過程中的功能進行了研究。將pFlag-CMV-Viperin和空載體pFlag-CMV-5a分別轉染至293T細胞,24 h后感染MDRV(MOI =1),感染24 h后收樣,提取總RNA,用RT-PCR和qRT-PCR檢測細胞中病毒P10基因的表達情況。結果表明,轉染pFlag-CMV-Viperin的P10基因表達量明顯低于對照組。因此說明,番鴨Viperin蛋白對MDRV的增殖起到抑制作用(圖5)。
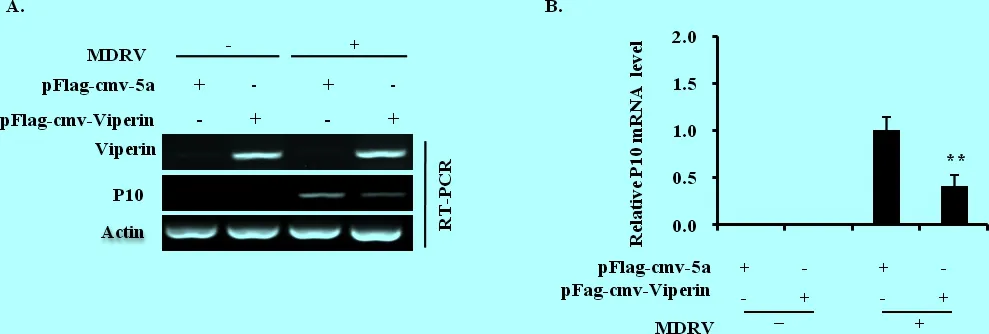
圖5 過表達Viperin抑制了MDRV在293T細胞中的復制
4 討論
番鴨呼腸孤病毒病在福建、廣東和浙江等多個省份暴發,對我國番鴨養殖業造成了重大的經濟損失[15],研制高效低毒的抗MDRV藥物勢在必行。先天免疫因子Viperin是一種可被干擾素、多種病毒、細菌脂多糖(LPS)等誘導表達的與內質網相關的抗病毒蛋白,具有廣譜抗病毒活性。Viperin在抑制包括DNA和RNA病毒,細菌和寄生蟲在內的廣泛的微生物入侵中扮演著重要角色。
本研究發現番鴨Viperin基因在感染MDRV的細胞和組織中大量表達,為了探明該現象與病毒復制的相關性,我們構建了番鴨Viperin 基因真核表達質粒,對其功能進行了研究。結果發現,番鴨Viperin蛋白在MDRV感染過程中發揮了重要作用:能夠顯著抑制MDRV的復制。后續工作可以聚焦于番鴨Viperin蛋白具體的作用機制及其調控的相關信號通路,以期更加全面地了解番鴨Viperin蛋白抗MDRV的分子機制。這將為闡明MDRV與宿主相互作用機制及病毒的致病機理提供重要科學依據,同時也為新型抗MDRV藥物的研發提供理論參考。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小學科學(學生版)(2020年10期)2020-10-28 07:52:12
中國化肥信息(2020年7期)2020-03-19 01:54:02
中國軍轉民(2017年6期)2018-01-31 02:22:28
海峽科技與產業(2016年3期)2016-05-17 04:32:12
汽車零部件(2014年11期)2014-09-18 11:57:16
