老芒麥種子離區酶活變化及組織學分析
2018-07-30 07:29:58張俊超謝文剛趙旭紅張宗瑜趙永強王彥榮
草業學報 2018年7期
張俊超,謝文剛,趙旭紅,張宗瑜,趙永強,王彥榮
(蘭州大學草地農業生態系統國家重點實驗室,蘭州大學農業農村部草牧業創新重點實驗室,蘭州大學草地農業科技學院,甘肅 蘭州 730020)
老芒麥(Elymussibiricus)是禾本科披堿草屬的多年生草本植物,是披堿草屬的模式種[1]。老芒麥主要分布于北半球的多個地區,包括東歐、北美、中國、瑞典及日本等地[2]。國內老芒麥資源亦較豐富,在新疆、內蒙古、青海、西藏等省區均有分布[3]。由于其較強的耐寒耐旱性,對環境的適應性強、產草量高、草品質好等優點,在我國西北地區,尤其是青藏高原地區常作為家畜的優良牧草[4]。同時老芒麥還有植被恢復、綠化護坡等用途[5]。然而,嚴重的落粒性常導致老芒麥種子產量的極大損失,由此限制了老芒麥新品種的研發及推廣應用。
國內外有關植物種子落粒的研究報道較多,主要集中在一些引種馴化早,栽培歷史較長的谷類作物,如水稻(Oryzasativa)、高粱(Sorghumbicolor)、小麥(Triticumaestivum)、大麥(Hordeumvulgare)等[6-9]。其中,Pourkheirandish等[10]認為野生大麥種子落粒是由于其具有較脆的花序軸,這一性狀由Btr1/2基因控制;Konishi等[6]研究表明水稻落粒過程會伴有離層的形成,且qSH1基因控制這一性狀。此外,在高粱和小麥種子脫落過程也存在這一現象[7-8];Ji等[11]認為多聚半乳糖醛酸酶和β-內切葡聚糖酶會引起水稻離層細胞的胞間層和細胞壁降解;Yoon等[12]研究發現OSH15蛋白可通過抑制粳稻(Oryzasativasubsp.japonica)離區木質素合成而誘導種子落粒。這些研究從形態學、組織學、生理學、基因組學及蛋白組學等層面闡述了引起種子脫落的機制。相對而言,關于禾本科牧草種子落粒的研究較少,已有文獻則集中在落粒評價、解剖結構觀察、表型性狀觀測等方面[13-18],對于綜合性與深層次分析落粒機理鮮有報道[19-20]。由于老芒麥作為栽培牧草被馴化與栽培歷史相對較短[21],國內外對老芒麥落粒的研究主要包括落粒率的評價[13],落粒與農藝性狀及產量性狀的關系[22],落粒個體間雜交[23]等,對老芒麥種子落粒解剖結構和生理機制研究仍相對較少[20,24]。有學者認為多年生禾本科牧草單粒種子脫落與離區發育有關[25]。因此,在開展落粒評價的同時,對材料離區解剖結構及發育狀況的研究將有利于揭示其落粒機理。
本研究通過對青藏高原地區野生老芒麥種子解剖結構和落粒關鍵酶的研究,分別從組織學及生理學方面揭示老芒麥種子落粒機理,為篩選低落粒材料及培育高產優質老芒麥新品種奠定研究基礎。
1 材料與方法
1.1 試驗材料
6種供試老芒麥均來源于甘肅省,于2016年3月使用種子進行室內萌發,待其長至3片葉子時,將材料移栽至榆中試驗地,采取常規田間管理方式進行管理。前期試驗已表明6種材料在落粒方面有所差異[26](表1)。
1.2 方法
1.2.1切片制作 在老芒麥抽穗后3和5周進行取樣。取花序中間1/3區域的小花(種子)并剪取花-花梗結構(長度2~4 mm),將材料置于FAA固定液(無水乙醇∶冰乙酸∶甲醛∶水=60∶5∶5∶30,添加少量甘油)中4 ℃保存。將固定24 h后的材料取出,經過不同濃度的酒精溶液進行脫水,再分別經過無水乙醇與二甲苯混合液(無水乙醇∶二甲苯=1∶1)和二甲苯溶液(100%)進行透明,之后轉移至石蠟中在烘箱浸蠟,利用硬質紙盒將材料包埋,在石蠟切片機上將蠟塊切至4~8 μm厚的蠟片,在烘箱中進行烤片,然后在不同濃度的二甲苯、無水乙醇溶液中進行脫蠟并復水,通過1%番紅溶液進行染色,最后用中性樹膠進行封片。待樹膠風干后于顯微鏡下成像并拍照。

表1 供試材料相關信息Table 1 Information of six E. sibiricus accessions in this study
WAH:抽穗后周數Weeks after heading.
1.2.2電鏡掃描 取樣時期為抽穗后3~5周,部位及材料固定同1.2.1,將固定過的材料取出,用鑷子輕輕掰斷花-花梗連接部位,將材料用膠粘附在金屬塊上。借助JSM-5600LV掃描電子顯微鏡(日本電子光學公司)進行觀察。
1.2.3纖維素酶和多聚半乳糖醛酸酶的測定 取樣時期為抽穗后1~5周,取樣部位同1.2.1,須在低溫環境下剪取花-花梗結構,并用錫箔紙包好,投入液氮中,再轉移至超低溫冰箱。提取時將低溫保存的樣品取出,放至液氮中,在電子天平(精確至0.0001 g)稱取約0.05 g的樣品,轉移至研缽中加液氮研磨呈粉末狀,加入500 μL提取緩沖液(預先配制好的檸檬酸-檸檬酸鈉緩沖液),其中提取纖維素酶時加入的緩沖液pH值為4.8,提取多聚半乳糖醛酸酶時加入的緩沖液pH值為6.0。用移液器將溶液轉移至1.5 mL離心管,4 ℃下12000 r·min-1離心20 min,之后吸取上清液250 μL至新離心管中,4 ℃保存。
試驗對纖維素酶和多聚半乳糖醛酸酶均采用Elisa試劑盒(上海酶聯生物科技有限公司)進行反應并在多功能酶標儀(Thermo Varioskan Flash 全波長多功能酶標儀)進行測定。
1.2.4斷裂拉伸張力測定 在老芒麥抽穗后4~5周,各材料隨機選取3~5個穗,借助電子拉力計測定斷裂拉伸張力[26]。主要測定步驟為:a)將組裝后的電子拉力計置于水平臺面,并將模式調整為峰值模式;b)將種子長度的一半處用電子拉力計的夾頭固定好;c)用手輕輕拉動穗部,待種子的種柄處發生斷裂時,電子拉力計會記錄數值。這個數值就表示該種子脫離母體時的斷裂拉伸張力[breaking tensile strength,BTS (N)]。每材料測定20個種子,則各材料的BTS值為其平均值。
1.3 數據處理
兩種酶的測定數據通過Excel 2013進行整理,借助Origin 8.5進行繪圖。
2 結果與分析
2.1 老芒麥種子離區解剖結構分析
以兩種落粒差異最顯著的材料(ZHN03與XH09)為例(圖1),分析它們在抽穗后3周和抽穗后5周時的離區結構(縱截面)變化。首先,在抽穗后3周時老芒麥種子離層結構已形成,而其種子落粒關鍵期為抽穗后4~5周,表明離層形成時期早于種子脫落。其次,離層由2~3層細胞構成(圖1E深紅色部分),這些細胞比鄰接的細胞體積要小且細胞壁厚度低于鄰接的細胞,形狀多為橢圓形,細胞伸長方向與周圍組織細胞互相垂直,細胞間排列緊湊。在染色過程中,離層細胞極易被番紅染為深紅色,而周圍組織細胞只有細胞壁被染為淺紅色,說明離層細胞的木質化程度遠高于其周圍組織細胞。在抽穗后3周時,兩種材料離層結構均完整,而ZHN03離層細胞木質化細胞數量比XH09多(圖1A,B);在抽穗后5周時,ZHN03離層木質化細胞數量仍多于XH09(圖1C,D),且ZHN03離層結構完整,而XH09離層有明顯的斷裂,因此木質化細胞數量對維持離層結構完整性有重要作用。
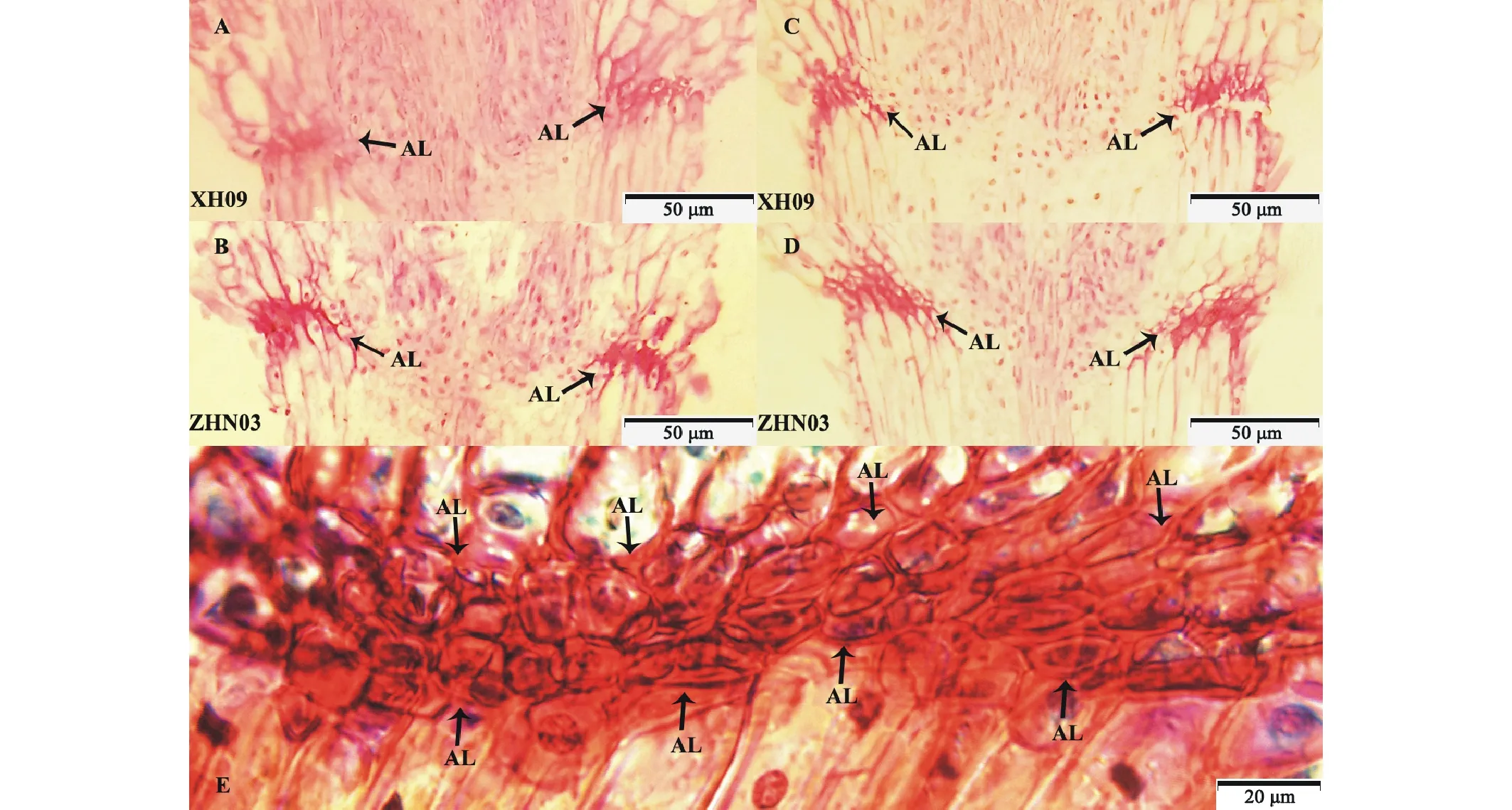
圖1 不同時期老芒麥種柄離區的變化(以ZHN03和XH09為例)Fig.1 Change of pedicel abscission zone at different stages of development in E. sibiricus (showed ZHN03 and XH09) 圖中展示了老芒麥抽穗后3周(A、B)和抽穗后5周(C、D)時離區的解剖結構,以及放大后的離層(E)。黑色箭頭是離層所在。This Figure show the anatomic structure of abscission zone of E. sibiricus at 3 weeks after heading (A, B) and 5 weeks after heading (C, D). And show the magnifying abscission layer (E). Black arrow indicates the location of abscission layer (AL).
2.2 老芒麥種子斷裂面掃描結構分析
6種老芒麥種子離區(橫截面)的電鏡掃描結果如圖2所示,由圖中可看出供試老芒麥種子斷裂面為非平面結構,離區(單個離層)由6~8圈細胞呈放射狀分布在外圍,單個細胞為橢圓形,細胞間排列緊湊而有規律。斷裂面中心有3個維管束(種子通過維管束與植株相連,在此具有物質輸送和支撐種子作用),呈凹陷狀,不同時期,維管束結構并未發生明顯變化(個別材料在抽穗后5周時離區中心位置出現了孔洞,這是材料在進行掃描電鏡前抽真空處理所致)。
6種材料在抽穗后3~5周內離區結構變化有所不同。其中低落粒材料在抽穗后3~5周期間,離區結構并未發生明顯變化,表現為離區表面粗糙,細胞間隙很明顯;高落粒材料在抽穗后3~4周時離區表面粗糙,細胞間隙明顯;抽穗后5周時離區表面變得光滑,細胞輪廓模糊,反映出該區域細胞發生降解(降解程度XH09>HZ02>LQ03>LT04)。
2.3 老芒麥種柄離區纖維素酶和多聚半乳糖醛酸酶活性變化
圖3為供試材料纖維素酶(cellulase,CE)活性在不同時期的變化情況。由圖中可看出6種材料在不同時期,它們的CE活性變化均不同。在抽穗后1~3周,6種材料CE活性變化較為一致,均呈現先下降后升高的趨勢,這一時期ZHN03與XH09的CE活性低于其他4種材料。在抽穗后3~5周,6種材料CE活性變化出現差異,其中ZHN03與XH09兩種材料CE持續升高,且XH09的CE活性升高幅度遠高于ZHN03;其他4種材料CE活性在此時期持續下降,其中MQ01與LT04下降幅度較大。在抽穗后5周時,ZNH03、MQ01與LT04的CE活性較為相近,HZ02與LQ03的CE活性相近,XH09為供試材料中CE活性最高的材料(XH09WAH5=496.38 IU·L-1),而ZHN03為最低(352.98 IU·L-1)。6種材料在落粒關鍵期CE活性的平均值大小為:XH09>HZ02>LQ03>LT04>MQ01>ZHN03。
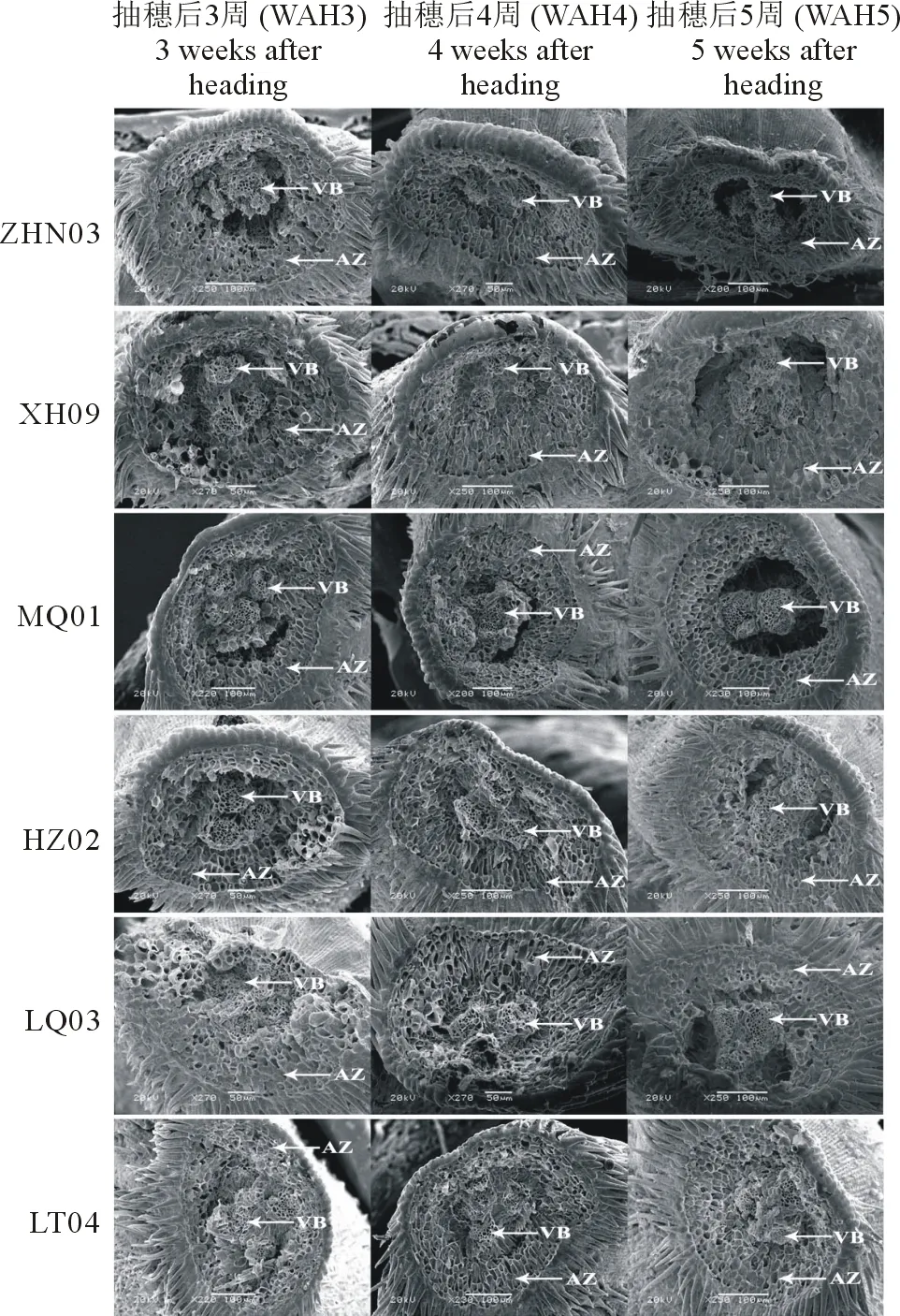
圖2 老芒麥種子離區電鏡掃描Fig.2 Scanning electron microscope of the abscission zone of E. sibiricus VB:維管束Vascular bundle;AZ:離區Abscission zone.
圖4為供試材料多聚半乳糖醛酸酶(polygalacturonase,PG)活性在不同時期的變化趨勢。由圖可看出,供試材料PG在不同時期變化與CE變化有所不同。在抽穗后1~3周,6種供試材料PG活性變化較一致,均呈先下降后升高趨勢,且ZHN03與XH09的PG活性低于其余4種材料。在抽穗后3~5周,XH09、LQ03與MQ01的PG活性呈現先升高后降低的趨勢,而其余3種材料的PG活性在此時期呈先下降后升高的趨勢。在抽穗后5周時,ZHN03與MQ01的PG活性較為接近,LQ03與LT04的PG活性相近,XH09的PG活性最高(XH09WAH5=186.5 pg·mL-1),HZ02次之(HZ02WAH5=165.06 pg·mL-1),而ZHN03為最低(ZHN03WAH5=124.77 pg·mL-1)。6種材料在落粒關鍵期PG活性的平均值大小為:XH09>HZ02>LQ03>LT04>MQ01>ZHN03。
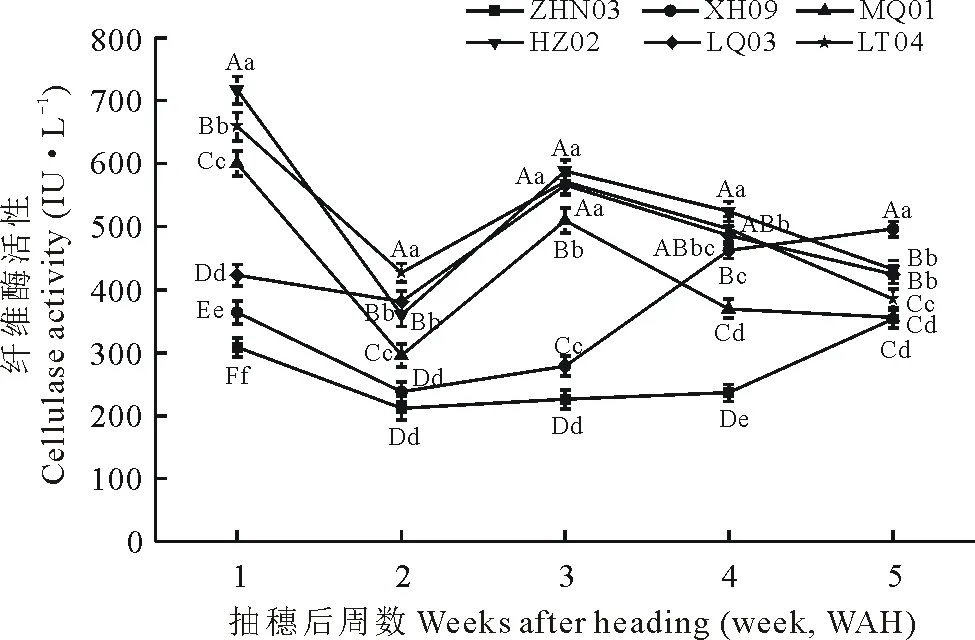
圖3 老芒麥材料不同時期纖維素酶活性變化Fig.3 Change of cellulase activity at different stages of development in E. sibiricus
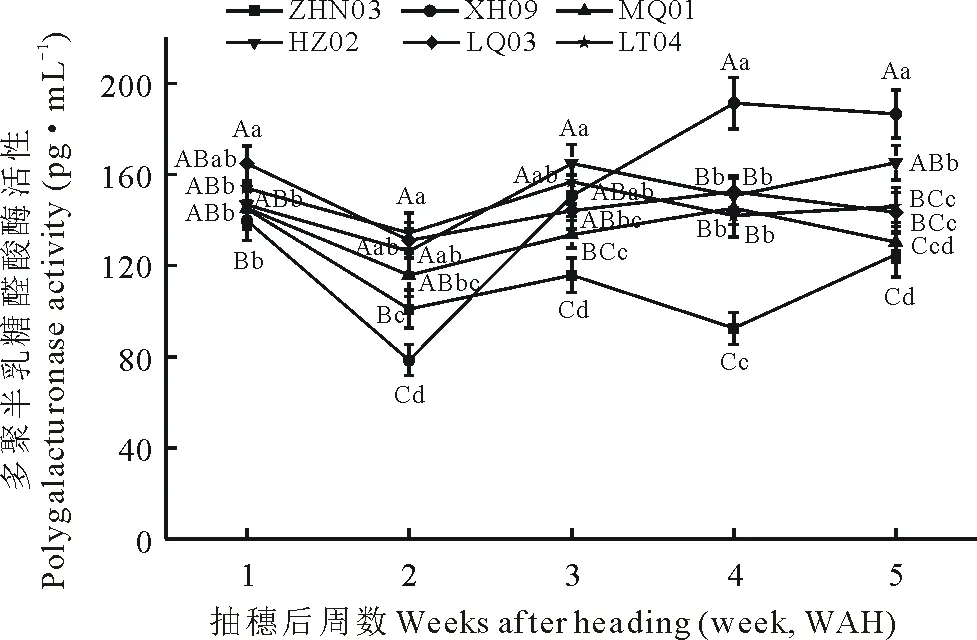
圖4 老芒麥材料不同時期多聚半乳糖醛酸酶活性變化Fig. 4 Change of polygalacturonase activity at different stages of development in E. sibiricus
同一時期內不同小寫和大寫字母表示不同材料間在0.05和0.01水平存在顯著性差異。Different small and capital letters during the same period mean significant difference among accessions at 0.05 and 0.01 levels, respectively.
2.4 老芒麥斷裂拉伸張力與細胞壁水解酶活性關系
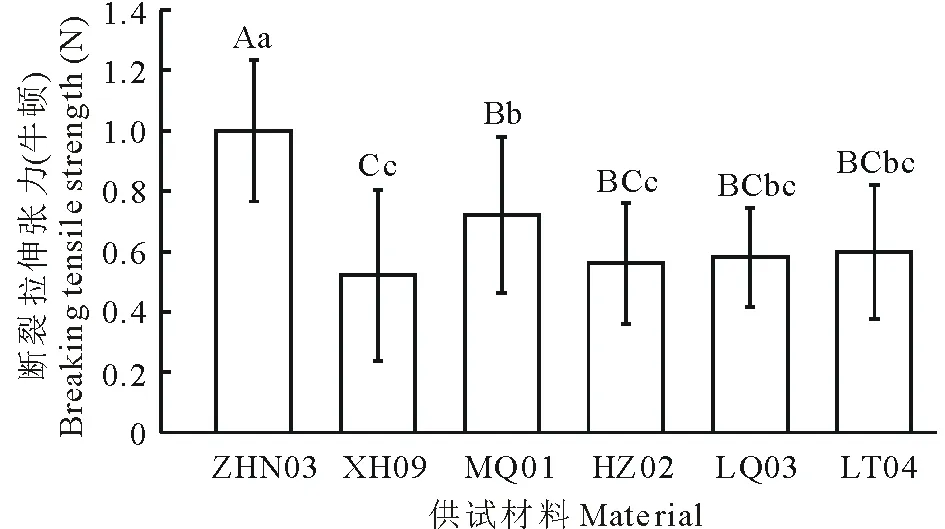
圖5 老芒麥6種供試材料斷裂拉伸張力值Fig.5 The value of breaking tensile strength of six E. sibiricus accessions in this study 不同小寫和大寫字母表示不同材料間在0.05和0.01水平存在顯著性差異。Different normal and capital letters mean significant difference among accessions at 0.05 and 0.01 levels, respectively.
圖5為供試材料在抽穗后4~5周時測定的斷裂拉伸張力值(BTS)。由上圖可以看出不同材料在種子落粒方面存在差異。其中,低落粒材料ZHN03的BTS值在0.01和0.05水平均顯著高于4種高落粒材料;低落粒材料MQ01的BTS值在0.01和0.05水平均顯著高于高落粒材料XH09。
表2分析了BTS與不同時期細胞壁水解酶活性的關系,其中BTS與WAH4時期纖維素酶活性及WAH3和WAH4時期多聚半乳糖醛酸酶活性呈極顯著負相關(r=-0.952,P<0.01;r=-0.900,P<0.01;r=-0.900,P<0.01),與WAH5時期纖維素酶和多聚半乳糖醛酸酶活性均呈顯著負相關(r=-0.746,P<0.05;r=-0.761,P<0.05)。該結果表明,老芒麥種子落粒與WAH4及WAH5時期兩種細胞壁水解酶活性密切相關。
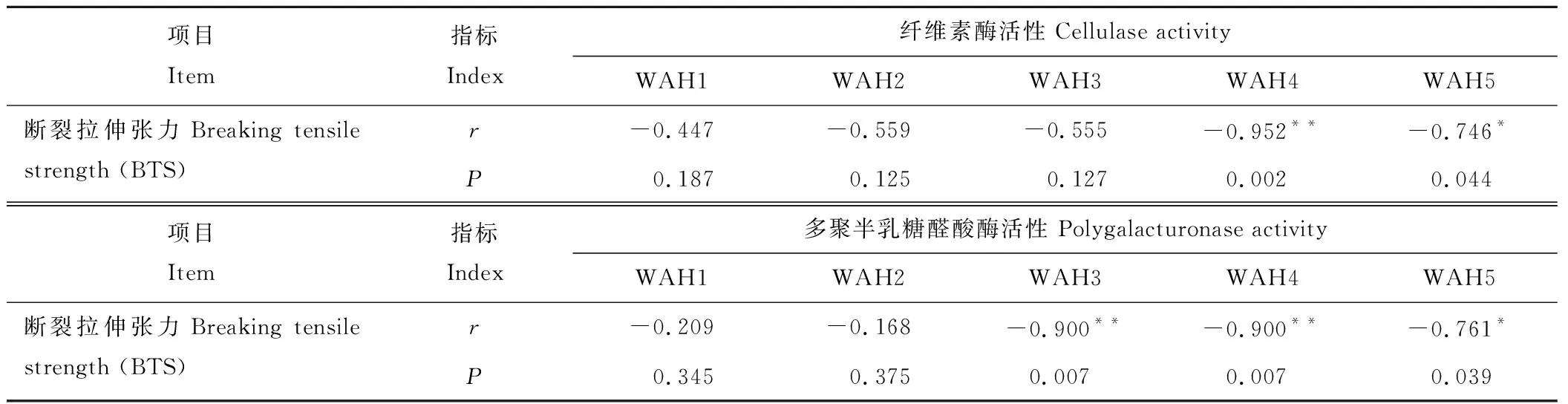
表2 老芒麥斷裂拉伸張力與細胞壁水解酶活性的關系分析Table 2 The relationship between breaking tensile strength and activities of cell wall hydrolases in E. sibiricus
注:*表示在0.05水平相關性顯著,**表示在0.01水平相關性顯著。
Note: *shows significant correlation at the level of 0.05,**shows significant correlation at the level of 0.01.
3 討論
3.1 老芒麥種子落粒與種子離區結構的關系
許多關于禾本科植物種子落粒的研究表明,種子離層的形成及降解過程與落粒具有相關性[8,18,27],如水稻的落粒特性是由某些基因的表達決定,這些基因直接控制著離層的發育[6,28-29]。雜草稻和野生稻雖均有落粒性,但二者的離層形成時期及降解時期均有明顯差異[30]。本研究中的切片和掃描電鏡結果表明供試老芒麥離區具有以下特點:離層細胞位于維管束周圍且其形成時期早于種子脫落,細胞壁加厚程度低;組成離層結構的細胞木質化程度較高;離區有維管束結構存在。這與Xie等[20]和Zhao等[24]對老芒麥離區的觀察結果相近。電鏡掃描結果發現高落粒材料在抽穗后5周時離區細胞有明顯降解,且不同材料間細胞降解程度不同,這與解剖結構觀察發現高落粒材料在抽穗后5周時離層結構不完整相符合。關于種子離區結構的形成與降解,在其他植物中多有報道。Burson等[27]比較百喜草(Paspalumnotatum)和毛花雀稗(Paspalumdilatatum)草種子離層時發現它們的離層在孕穗期就已形成,但毛花雀稗草離層形成時期比百喜草稍早一些。Elgersma等[18]報道了多年生黑麥草(Loliumperenne)種子離區在抽穗階段已形成,但在種子脫落時期未發現明顯的細胞降解過程。Inoue等[31]研究表明野生稻種子離層形成于開花期,并在授粉后開始降解。此外,本研究結果表明,供試老芒麥中的高落粒與低落粒材料在離層木質化細胞數量上存在明顯差異,離層木質化細胞數量越多,細胞壁的機械強度與致密度越大,在種子成熟期維持離層結構穩定性的能力就越強。Yoon等[29]研究認為種子離區木質素含量的高低與水稻種子脫落有關,通常較低的木質素含量更易引起水稻種子落粒。因此,離層細胞的木質化程度是造成老芒麥不同落粒特性的原因之一。
3.2 老芒麥落粒與種子離區細胞壁水解酶的關系
纖維素酶(CE)和多聚半乳糖醛酸酶(PG)是植物細胞壁的兩種主要降解酶。前者主要降解植物細胞壁中的纖維素成分,后者則主要降解細胞壁中的果膠成分[32],許多研究表明這兩種酶參與植物離層細胞的降解過程[33-34]。關于老芒麥種子離層細胞的降解過程,Xie等[20]研究表明在老芒麥種子脫落期間纖維素酶和多聚半乳糖醛酸酶活性均顯著升高,且高落粒材料兩種酶活性遠高于低落粒材料。Zhao等[24]研究發現由栽培材料和品種組成的老芒麥群體與野生老芒麥群體在種子脫落期間兩種酶的差異水平并不顯著。本研究所用材料均為野生老芒麥,酶活測定結果表明,不同時期兩種酶在不同材料間的變化趨勢有所差異,種子落粒關鍵期的兩種細胞壁水解酶活性高低與斷裂面觀察結果相符,這一時期細胞壁水解酶活性與種子落粒呈顯著正相關。關于纖維素酶和多聚半乳糖醛酸酶在種子(果實)脫落(裂莢)過程中起到的作用有不同的報道。如陳發河等[35]研究認為葡萄(Vitisvinifera)離區纖維素酶和多聚半乳糖醛酸酶活性增加會加劇葡萄的落粒進程。Agrawal等[36]研究表明纖維素酶直接參與并促進大豆(Glycinemax)的裂莢過程,而多聚半乳糖醛酸酶在此過程中具有軟化果莢防止其開裂的作用。這些研究結果表明,不同植物中,這兩種酶在落粒(裂莢)過程中產生的作用有所差異,在今后的研究工作中也需要對此加以參考和深入分析。
結合組織觀察和酶活性測定結果,推測老芒麥種子保持及脫離植株的機理為:WAH2~WAH3時期是種子形成重要時期,植株通過維管束向種子內輸送營養物質,種子內部的纖維素酶和多聚半乳糖醛酸酶活性逐漸增加,由于種子離區既存在木質化的細胞,也有維管束的支撐,因而可以保持不落狀態;WAH4~WAH5時期是種子脫落高發期,老芒麥高落粒與低落粒材料相比,其種子離區纖維素酶和多聚半乳糖醛酸酶活性較高且離區細胞木質化水平較低,在細胞壁酶的作用下,離層細胞更易降解,加之維管束因種子脫水作用變得干脆易斷,因而種子在外力的作用下很容易脫落。
4 結論
通過對青藏高原野生老芒麥離區組織學觀察和酶活性測定的研究表明,供試老芒麥種子落粒與其離區的形成及降解過程有關。老芒麥離區在形成過程中的木質化有利于種子成熟前的保持,但落粒關鍵期細胞壁水解酶活性增強會促使離區結構降解,使得種子最終脫離植株。木質化水平高低及落粒關鍵期細胞壁水解酶活性強弱是產生落粒差異的主要原因。該研究從組織學和生理學層面揭示了老芒麥種子落粒機理,為今后篩選老芒麥低落粒材料和培育低落粒品種提供借鑒和參考。
