miRNA-24-3P 靶向調(diào)節(jié)SP1對RAW264.7細胞功能的影響*
2018-07-05 06:07:00程東良陳衍晨
重慶醫(yī)學 2018年17期
程東良,梁 媛,陳衍晨,卿 娣
(1.鄭州大學人民醫(yī)院/河南省人民醫(yī)院兒科,鄭州 450000;2.河南省開封市兒童醫(yī)院血液科 470000;3.南方醫(yī)科大學珠江醫(yī)院兒科,廣州 510282;4.廣東省廣州市婦女兒童醫(yī)療中心呼吸科 510623)
急性呼吸窘迫綜合征(actu respiratory distress syn drom,ARDS)現(xiàn)在仍是兒童重癥監(jiān)護室(pediatric intensive careunit,PICU)中威脅生命的重要原因之一。ARDS的發(fā)病機制未完全明了,目前主要考慮與炎癥有關(guān),探尋ARDS的發(fā)病機制及有效的治療手段是危重病醫(yī)學面臨的重要臨床課題。
微小RNAs(microRNAs,miRNA)是一類具有20~22個核苷酸單位的調(diào)控性RNA,通過與信使RNA序列的結(jié)合或者在轉(zhuǎn)錄后水平抑制目的基因的表達來發(fā)揮作用[1]。通過前期的研究[2],筆者發(fā)現(xiàn)在ARDS小鼠模型的肺組織中miR-24-3P顯著差下調(diào),對miR-24-3P進行靶基因的預測發(fā)現(xiàn)SP1基因是miR-24-3P其中一個候選靶基因。SP1是炎性反應過程中的重要反應蛋白,本研究將SP1基因作為miR-24-3P的候選目的基因,探討SP1是否為miR-24-3P的直接靶基因蛋白。
1 材料與方法
1.1材料 RAW264.7細胞、HEK293細胞由南方醫(yī)科大學珠江醫(yī)院再生醫(yī)學研究所提供,胎牛血清為澳大利亞的Hyclone血清,脂多糖購自美國的Sigma公司,miRNA mimics、miRNA inhibitor及陰性對照購自廣州銳博公司,小鼠SP1、核因子-κB(NF-κB)、GAPDH一抗購自Abcam公司,Trizol、M-MLV反轉(zhuǎn)錄試劑盒、SYBR Green qPCR試劑盒、Lipofectamine 2000均購于美國Invitrogen公司,雙熒光素酶報告基因檢測試劑盒購自Promega公司。倒置顯微鏡為日本Olympus公司產(chǎn)品,定量PCR儀為ABI PRISM 7500 Sequence Detection System,Western blot電泳儀及轉(zhuǎn)膜儀為美國的Bio-Rad品牌產(chǎn)品。
1.2方法
1.2.1細胞培養(yǎng)及實驗分組 從液氮中取出凍存的細胞RAW264.7、HEK293細胞,水浴中劇烈搖晃使細胞快速溶解,3 000 r/min離心5 min,吸棄凍存液,用磷酸鹽緩沖液(PBS)洗滌細胞2遍,再用新鮮的完全培養(yǎng)基DMEM(含10%的胎牛血清、100 U青/鏈霉素)懸浮細胞。培養(yǎng)條件均為37 ℃、5% CO2及飽和濕度,倒置顯微鏡觀察細胞生長狀態(tài)。等待細胞長滿后用0.25%胰蛋白酶消化,凍存或傳代。實驗分為5組:具體加樣見下表。Mock組(miR-24-3P mimics+OPTI-MEI培養(yǎng)基)、miR-24-3P mimics組(miR-24-3P mimics+Lipofectamine 2000)、miR-24-3P mimics control組(miR-24-3P mimics control+Lipofectamine 2000)、miR-24-3P inhibitor組(miR-24-3P inhibitor+Lipofectamine 2000)、miR-24-3P inhibitor control組(miR-24-3P inhibitor+Lipofectamine 2000)。
1.2.2RAW264.7細胞轉(zhuǎn)染 轉(zhuǎn)染前1 d,1×106個/孔細胞鋪至6孔培養(yǎng)板中,過夜培養(yǎng),待第2天細胞生長至50%融合度時進行轉(zhuǎn)染。轉(zhuǎn)染前2 h給細胞上OPTI-MEM(無血清、無雙抗)培養(yǎng)基,1.5 mL/孔。采用250 μL的OPTI-MEI培養(yǎng)基稀釋各組siRNA至適當濃度(mimics、mimics control轉(zhuǎn)染時終濃度為50 nmol/L;inhibitor、inhibitor control轉(zhuǎn)染時終濃度為100 nmol/L),輕輕混勻。輕輕搖勻轉(zhuǎn)染劑Lipofectamine 2000,取10 μL溶于250 μL的OPTI-MEI培養(yǎng)基中,室溫孵育5 min。將前兩步所稀釋的液體混合成轉(zhuǎn)染液,輕輕混勻,室溫孵育20 min。將轉(zhuǎn)染液加入培養(yǎng)板,輕輕搖勻。置于37 ℃、5% CO2培養(yǎng)箱中培養(yǎng),轉(zhuǎn)染后(用于RNA檢測需轉(zhuǎn)染16 h,用于蛋白檢測則需轉(zhuǎn)染36 h)每孔中加入終濃度為100 ng/mL脂多糖進行刺激[3],3 h后收集細胞進行檢測。
1.2.3實時熒光定量PCR(Q-PCR)檢測各組細胞中miR-24-3P的表達水平 取以上各組處理后的細胞,用PBS沖洗 3 遍,使用Trizol、氯仿等提取出各組細胞的總RNA,使用分光光度計算出各組總RNA的濃度及A260 nm/A280 nm比值以驗證RNA的純度。并進行瓊脂糖RNA電泳檢驗RNA的完整性。使用通用反轉(zhuǎn)錄得到cDNA,U6為內(nèi)參,具體操作按試劑盒說明書進行。miR-24-3P上游引物為5′-TGG CTC AGT TCA GCA GGA ACA G-3′;下游引物為5′-GAT CCA GTC TCA GGG TCC GAG-3′。U6上游引物為5′-CTC GCT TCG GCA GCA CA-3′,下游引物為5′-AAC GCT TCA CGA ATT TGC GT-3′。反應體系:cDNA(1∶20) 5.0 μL,上游引物 0.5 μL,下游引物 0.5 μL,SYBR Green PCR Master Mix 10.0 μL,H2O 4.0 μL,共20 μL。反應條件:95 ℃ 5 min;95 ℃ 15 s,65 ℃,15 s,72 ℃ 32 s,40個循環(huán)。為了建立PCR產(chǎn)物的熔解曲線,擴增反應結(jié)束后繼續(xù)從 72 ℃緩慢加熱到 95 ℃。每個標本重復 3 次。反應結(jié)束后分析PCR反應曲線,得到各組的Ct值。目的基因相對表達量:實驗組Ct值/對照組Ct值。
1.2.4Q-PCR檢測各種細胞中腫瘤壞死因子α(TNF-α)mRNA、白細胞介素-6(IL-6)mRNA、SP1 mRNA、NF-κB mRNA的相對表達水平 取以上各組轉(zhuǎn)染及處理后的細胞,用PBS沖洗3遍,使用Trizol、氯仿等提取出各組細胞的總RNA,使用分光光度計算出各組總RNA的濃度及A260 nm/A280 nm比值以驗證RNA的純度。并進行瓊脂糖RNA電泳檢驗RNA的完整性。按照說明書進行反轉(zhuǎn)錄得到cDNA,以GAPDH為內(nèi)參進行PCR反應。TNF-α上游引物為5′-CCC CAG TCT GTA TCC TTC TAA-3′,下游引物為5′-TCG AGG CTC CAG TGA ATT-3′,上游引物為5′-ACA ACC ACG GCC TTC CCT ACT T-3′,下游引物為5′-AGC CTC CGA CTT GTG AAG TGG T-3′;SP1上游引物為5′-GAT TTC TCC ACA GCA CTC ATG-3′,下游引物為5′-TTA GAG GAG CTG AAG GAC AGA-3′;NF-κB 上游引物為5′-GAC AAA AAC TGG GCC ACT CT-3′,下游引物為5′-GAT CAC TTC AAT GGC CTC TGT-3′;GAPDH上游引物為5′-GGC CTC CAA GGA GTA AGA AA -3′,下游引物為5′-GCC CCT CCT GTT ATT ATG G-3′。反應體系:cDNA(1∶20)5.0 μL,上游引物 0.5 μL,下游引物 0.5 μL,SYBR Green PCR Master Mix 10.0 μL,H2O 4.0 μL,共20.0 μL。反應條件:95 ℃ 2 min;95 ℃ 15 s;60 ℃ 32 s,40 個循環(huán),為了建立PCR產(chǎn)物的熔解曲線,擴增反應結(jié)束后繼續(xù)從 60 ℃緩慢加熱到 95 ℃。每個標本重復 3 次。反應結(jié)束后分析PCR反應曲線,得到各組的Ct值。目的基因相對表達水平:實驗組Ct值/對照組CT值。
1.2.5Western blot檢測各組細胞中SP1、NF-κB的表達水平 取上述各組處理后的細胞,用PBS沖洗3遍,按說明書提取各組細胞中的核蛋白,使用Bradford分光光度法測定蛋白含量。每組樣品取20 μg核蛋白樣品分別加到預制的聚丙烯酰胺凝膠后80 V電泳2 h,然后,20 V 30 min轉(zhuǎn)移至PVDF膜,封閉過夜,分別加入一抗抗體 4 ℃孵育12 h,TBST充分洗膜后再分別與二抗室溫孵育1 h,TBST充分洗膜,進行ECL化學發(fā)光顯色。曝光后掃描,分別以SP1/GAPDH、NF-κB(P65)/GAPDH顯影條帶灰度值的比值來代表SP1、NF-KB(P65)的相對表達水平。
1.2.6雙熒光素酶報告基因檢測 野生型及突變型SP1 3′-UTR載體構(gòu)建及鑒定由上海吉凱生物技術(shù)有限責任公司完成。轉(zhuǎn)染前24 h將對數(shù)生長期的HEK293細胞接種于24孔板中,當細胞貼壁并且匯合度達到30%時,按照Lipofectmine 2000說明書進行瞬時轉(zhuǎn)染。設(shè)轉(zhuǎn)染miR-24-3P mimics的實驗組和轉(zhuǎn)染對照序列的陰性對照組。轉(zhuǎn)染后24 h,用1×PBS洗滌細胞,盡可能的棄盡殘存液體。每孔各加入100 μL PLB,室溫搖床搖動20 min充分裂解細胞。將20 μL熒光酶素測試試劑(LAR)加人孔板中,儀器上讀取熒光素酶的熒光值(整合時間2 s,讀數(shù)時間10 s),然后再加入20 μL緩沖液,放入儀器讀取海腎熒光素酶的熒光值。
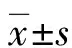
2 結(jié) 果
2.1轉(zhuǎn)染后各組細胞中miR-24-3P的相對表達水平 轉(zhuǎn)染后在熒光顯微鏡下觀察見到細胞質(zhì)中有綠色熒光。收集細胞后對各組細胞做Q-PCR,后計算各組2-△△Ct值為各組細胞miR-24-3P相對表達水平,結(jié)果顯示:Mock組miR-24-3P的相對表達水平是256.85±90.12,miR-24-3P mimics組194 215.66±12 555.57,miR-24-3P mimics control組145.99±22.4,miR-24-3P inhibitor組1.04±0.23,miR-24-3P inhibitor control組173.59±15.94。miR-24-3P mimics組、miR-24-3P inhibitor組和mock組比較差異有統(tǒng)計學意義(P<0.05),提示轉(zhuǎn)染成功,見圖1。
2.2Q-PCR檢測各種細胞中TNF-α mRNA、IL-6 mRNA、SP1 mRNA、NF-κB mRNA的相對表達水平 與Mock組比較,TNF-α mRNA、IL-6 mRNA在轉(zhuǎn)染miR-24-3P mimics組細胞中的表達水平顯著下降,在miR-24-3P inhibitor組細胞中的表達水平顯著上升,差異均有統(tǒng)計學意義(P<0.05,圖2)。SP1 mRNA、NF-κB mRNA 在各組細胞中的相對表達水平?jīng)]有顯著變化,差異無統(tǒng)計學意義(P>0.05),見圖2。

1:mock組;2:miR-24-3P mimics組;3:miR-24-3P mimics control組;4:miR-24-3P inhibitor組;5:miR-24-3P inhibitor control組
圖1 miR-24-3P在各組細胞中的相對表達水平分析圖

1:Mock組;2:miR-24-3P mimics組;3:miR-24-3P mimics control組;4:miR-24-3P inhibitor組;5:miR-24-3P inhibitor control組
圖2 TNF-α mRNA、IL-6 mRNA、SP1 mRNA、NF-κBmRNA在各組細胞中的相對表達水平分析圖
2.3Western blot檢測各組細胞中SP1、NF-κB的表達水平 與Mock組比較,SP1在miR-24-3P mimics組細胞中的表達水平顯著下降,在miR-24-3P inhibitor組細胞中的表達水平顯著上升,差異均有統(tǒng)計學意義(P<0.05,圖3)。NF-κB在各組細胞中的相對表達水平無顯著意義,差異均無統(tǒng)計學意義(P>0.05),見圖3。

1:Mock組;2:miR-24-3P mimics組;3:miR-24-3P mimics control組;4:miR-24-3P inhibitor組;5:miR-24-3P inhibitor control組
圖3 SP1、NF-κB在各組細胞中的表達水平
2.4雙熒光素酶報告基因檢測 雙熒光素酶報告基因檢測結(jié)果顯示,與陰性對照組比較,轉(zhuǎn)染miR-24-3 mimics后能夠使標化的熒光值降低59%(P<0.01),而突變株熒光值則未有明顯改變(P>0.05),見圖4。

圖4 雙熒光素酶報告基因檢測結(jié)果
3 討 論
ARDS是指膿毒癥、重癥肺炎等肺內(nèi)外疾病侵襲后出現(xiàn)的以肺泡毛細血管損傷為主要表現(xiàn)的臨床綜合征。其臨床特征包括呼吸頻率加快和呼吸窘迫,進行性低氧血癥,胸部X射線呈現(xiàn)彌漫性肺泡浸潤。現(xiàn)階段ARDS的發(fā)病機制主要為:多種炎性介質(zhì)大量釋放引起肺泡上皮細胞及肺泡內(nèi)皮細胞的損傷、肺泡中積聚富含蛋白及中性粒細胞的滲出液,形成透明膜[4-5],后期出現(xiàn)肺纖維化。參與炎性反應的細胞主要有多形核白細胞、巨噬細胞等。巨噬細胞受到刺激后可釋放白三烯、IL-1、TNF-α等炎性介質(zhì),引發(fā)強烈的炎性反應,同時在肺泡局部釋放大量蛋白溶解酶、氧自由基,強烈趨化多形核白細胞在肺內(nèi)聚集,導致炎癥的級聯(lián)反應及ARDS的發(fā)生。ARDS目前尚無特效治療方法,臨床上主要采取積極治療原發(fā)病、控制感染、抑制炎癥、呼吸及循環(huán)功能支持、防治并發(fā)癥及多器官功能衰竭等綜合措施[6-8]。
miRNA是一類廣泛存在于真核生物中的、長約21~25 堿基的非編碼小分子RNA。它的作用機制是以不完全互補的方式和靶基因的3′端非編碼區(qū)序列相結(jié)合,通過抑制蛋白質(zhì)的翻譯發(fā)揮它的生物學功能,這種調(diào)控一般不會影響靶基因mRNA的穩(wěn)定性[9]。miRNA在ARDS發(fā)生發(fā)展過程中的作用越來越引起人們的重視。CAI等[10]通過注射脂多糖建立了ALI/ARDS小鼠模型,然后進行miRNA芯片篩查,發(fā)現(xiàn)了大量差異表達的miRNAs,并用RT-PCR的方法檢驗其表達的差異。使用內(nèi)毒素處理巨噬細胞,然后進行miRNA芯片篩查,發(fā)現(xiàn)miR-132、miR-146a、miR-155表達水平增加,miR-125b水平下降[11-12]。XIE等[13]在博來霉素致小鼠肺纖維化模型中發(fā)現(xiàn)161個差異表達的miRNA。LIU等[14]發(fā)現(xiàn)在特發(fā)性肺纖維化患者及博來霉素致肺纖維化模型小鼠中miR-21均高表達,腫瘤壞死因子-β(TGF-β)也可以促進miR-21的表達。miR-155可以標靶角化細胞生長因子從而調(diào)節(jié)肺纖維化,其水平的高低與肺纖維化程度有關(guān)[15]。
筆者通過氣管內(nèi)滴注脂多糖構(gòu)建小鼠ARDS模型,發(fā)現(xiàn)miR-24-3P顯著下調(diào),預測其靶基因并進行深度分析,選取了候選miRNA miR-24-3P及其候選靶基因SP1進行進一步的研究。通過本實驗的研究筆者發(fā)現(xiàn)miR-24-3P對炎癥因子的分泌具有調(diào)控作用。炎性因子的分泌主要通過兩個途徑進行的,一個是NF-κB通路;另一個是MAPK通路,其通路末端的關(guān)鍵轉(zhuǎn)錄因子SP1、APl、STATl、CREB、MAX均與炎性基因轉(zhuǎn)錄有關(guān)。當人為的調(diào)節(jié)細胞內(nèi)miR-24-3P表達時,細胞內(nèi)的NF-κB蛋白及mRNA的表達均未受到影響,這說明miR-24-3P過表達下調(diào)TNF-α、IL-6 mRNA不是通過調(diào)控NF-κB轉(zhuǎn)錄活性這一途徑實現(xiàn)的。為了檢測miR-24-3P在細胞內(nèi)是否存在對SP1蛋白的調(diào)控作用,筆者在RAW264.7細胞中分別轉(zhuǎn)染了miR-24-3P及其特異抑制劑,檢測核內(nèi)SP1蛋白水平。結(jié)果表明miR-24-3P能抑制核內(nèi)SP1水平,當把miR-24-3P功能抑制后,核內(nèi)SP1水平獲得恢復。當在用miR-1224轉(zhuǎn)染RAW264 7細胞,檢測其對SP1基因mRNA水平的影響,定量結(jié)果顯示miR-24-3P mimics的過表達對SP1基因在mRNA水平上沒有顯著影響。通過熒光素酶報告實驗結(jié)果表明,轉(zhuǎn)染miR-24-3P可以明顯下調(diào)SP1-3′UTR相連的熒光素酶活性。所以得出結(jié)論:miR-24-3P可在轉(zhuǎn)錄后水平上抑制SP1基因的翻譯,限制SPl的蛋白水平,進而影響炎性介質(zhì)的釋放。RAW264.7細胞是小鼠單核/巨噬細胞,單核/巨噬細胞是ARDS過程中主要的反應細胞,所以本實驗選用RAW264.7細胞作為實驗細胞,檢測miR-24-3P對RAW264.7細胞釋放炎性介質(zhì)的影響。當然本實驗具有一定的局限性,未做動物實驗,這是筆者下一步實驗的重點。
[1]OZEN M,CREIGHTON C,OZDEMIR M,et al.Widespread deregulation of microRNA expression in human prostate cancer[J].Oncogene,2008,27(12):1788-1793.
[2]王斌,卿娣,程東良,等.脂多糖誘發(fā)的小鼠急性肺損傷肺組織中microRNA表達譜變化[J].中華實用兒科臨床雜志,2013,28(12):931-935.
[3]NIU Y,MO D,QIN L,et al.Lipopolysaccharide-induced miR-1224 negativelyregulates tumour necrosis factor-α gene expression by modulating Sp1[J].Immunology,2011,133(1):8-20.
[4]LIN J,ZHANG W,WANG L,et al.Diagnostic and prognostic values of Club cell protein 16(CC16) in critical care patients with acute respiratory distress syndrome[J].J Clin Lab Anal,2017,32(2).
[5]EPELBAUM O,ARONOW W S.Mechanical ventilation in the acute respiratory distress syndrome[J].Hosp Pract(1995),2017,45(3):88-98.
[6]YANG T Z,LIU Y,LIU Y Y,et al.The use of rheum palmatum L.In the treatment of acute respiratory distress syndrome:a Meta-analysis of randomized,controlled trials[J].Afr J Tradit Complement Altern Med,2017,14(2):334-347.
[7]VADASZ I.Keratinocyte growth factor in acute respiratory distress syndrome[J].Lancet Respir Med,2017,5(6):459-460.
[8]JONES T K,MEYER N J.What′s in a number? Platelet cunt dynamics as a novel mediator of acute respiratory distress syndrome survival[J].Am J Respir Crit Care Med,2017,195(10):1285-1287.
[9] ZEALY R W,WRENN S P,DAVILA S,et al.MicroRNA-binding proteins:specificity and function[J].Wiley Interdiscip Rev RNA,2017,8(5).
[10]CAI Z G,ZHANG S M,ZHANG Y,et al.MicroRNAs are dynamically regulated and play an important role in LPS-induced lung injury[J].Can J Physiol Pharmacol,2012,90(1):37-43.
[11]AMBROS V.microRNAs:tiny regulators with great potential[J].Cell,2001,107(7):823-826.
[12]ALTMAN N.Replication,variation and normalisation in microarray experiments[J].Appl Bioinformatics,2005,4(1):33-44.
[13]XIE T,LIANG J,GUO R,et al.Comprehensive microRNA analysis in bleomycin-induced pulmonary fibrosis identifies multiple sites of molecularregulation[J].Physiol Genomics,2011,43(9):479-487.
[14]LIU G,FRIGGERI A,YANG Y,et al.miR-21 mediates fibrogenic activation ofpulmonary fibroblasts and lung fibrosis[J].J Exp Med,2010,207(8):1589-1597.
[15]POTTIER N,MAURIN T,CHEVALIER B,et al.Identification of keratinocyte growth factor as a target of microRNA-155 in lung fibroblasts:implication in epithelial-mesenchymal interactions[J].PLoS One,2009,4(8):e6718.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2022年2期)2022-11-25 23:46:31
昆明醫(yī)科大學學報(2022年4期)2022-05-23 13:04:50
美與時代·美術(shù)學刊(2022年3期)2022-04-27 01:18:15
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
國際呼吸雜志(2019年21期)2019-11-25 09:52:20
人大建設(shè)(2019年12期)2019-05-21 02:55:32
天津醫(yī)科大學學報(2015年3期)2015-06-05 12:21:49
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
