SIRT1信號通路介導咖啡因預防早產兒支氣管肺發(fā)育不良
2018-07-05 11:14:30董文斌雷小平劉興鈴李清平
實用藥物與臨床 2018年6期
劉 洋,董文斌,雷小平,劉興鈴,李清平,康 蘭,趙 帥,張 嬋
0 引言
隨著醫(yī)療技術的提高,早產兒存活率逐漸增加,同時,早產兒支氣管肺發(fā)育不良(Bronchopulmonary dysplasia,BPD)的發(fā)生率也隨之增加。目前認為,任何氧依賴超過28 d的新生兒即可診斷支氣管肺發(fā)育不良[1]。高氧暴露是BPD發(fā)生的重要因素。長時間暴露在高氧環(huán)境中,產生大量活性氧簇(Reactive oxygen species,ROS),超過機體清除能力時,可發(fā)生DNA損傷、線粒體功能障礙、生物膜脂質過氧化,從而造成細胞功能障礙,引起組織器官的損傷[2]。沉默接合型信息調節(jié)因子2 同源蛋白1(Sirtuin1,SIRT1)是依賴NAD+(煙酰胺腺嘌呤二核苷酸)的去乙酰化酶,可在第734位賴氨酸上發(fā)生小泛素化樣修飾(Small ubiquitin-related modifier,SUMO),其去SUMO修飾的過程則是由SUMO特異性蛋白酶1(SUMO-specific protease1,SENP1)來介導。SENP1去SUMO化作用可以降低去乙酰化酶活性,使SIRT1的去乙酰化減少,促進SIRT1核漿轉位,依賴caspase途徑使細胞凋亡蛋白活性增加,促進細胞凋亡[3-4]。氧化應激能提高大多數細胞的p53乙酰化水平。乙酰化p53轉錄激活其下游基因的表達,誘導細胞凋亡[5]。本課題組已證實ROS可能通過激活SENP1,使SIRT1去SUMO化作用增強,促發(fā)SIRT1核漿穿梭,乙酰化p53增加,從而導致細胞凋亡[4,6-7]。
咖啡因屬甲基黃嘌呤類藥物,非特異性腺苷受體阻滯劑,該類藥物用于預防和治療早產兒呼吸暫停已30余年,且早期咖啡因的使用能夠預防BPD的發(fā)生[8-10],但是具體機制尚未明確。有學者發(fā)現,咖啡因可以減少人肺泡上皮細胞(A549)以及大鼠肺泡上皮細胞(MLE12)的高氧損傷[11-12]。但咖啡因預防BPD的機制以及與SENP1-SIRT1-p53的關系尚不清楚。本研究探討咖啡因對BPD的預防機制,為咖啡因臨床應用于BPD的預防提供理論依據。
1 資料與方法
1.1 研究對象 選取2015年8月至2016年6月西南醫(yī)科大學附屬醫(yī)院新生兒科入院的65例早產兒為研究對象。納入標準:出生胎齡<32周、體重<1 500 g,生后24 h內入院,入院即診斷為新生兒呼吸窘迫綜合征。排除標準:住院時間<28 d,合并復雜先天性心臟病、先天性傳染病、溶血性疾病及其他先天畸形的早產兒。
根據吸氧濃度分為對照組(n=47)和高氧組(n=18)。對照組氧濃度(FiO2)為21%或>21%但總吸氧時間<24 h;高氧組FiO2>40%且總吸氧時間>24 h。再根據家屬意愿是否使用咖啡因,分為對照組(22例)、對照+咖啡因組(25例)、高氧組(6例)、高氧+咖啡因組(12例)。對照組、對照+咖啡因組、高氧組、高氧+對照組胎齡分別為(30.28±0.94)、(30.18±1.38)、(29.5±1.11)、(29.26±1.88)周;出生體重分別為(1 228±110)、(1 230±87)、(1 138±114)、(1 242±24) g;四組患兒胎齡、出生體重、PS使用率、呼吸機使用率、呼吸機使用時間比較差異無統(tǒng)計學意義(P>0.05),高氧組及高氧+咖啡因組激素使用率及使用時間比較差異無統(tǒng)計學意義(P>0.05)。若對照組中患兒出現呼吸暫停需晚期使用(生后3 d后)咖啡因或(和)呼吸機治療則退出研究。
本研究中咖啡因的使用結合患兒病情及家屬的經濟條件,根據患兒家屬意愿使用,且本研究經醫(yī)院倫理委員會批準及患兒家屬知情同意。
枸櫞酸咖啡因的使用參照文獻[13],入院后早期(生后3 d內)預防性使用咖啡因,首次負荷劑量為20 mg/kg(相當于咖啡因10 mg/kg) 30 min內泵入,24 h后使用維持劑量5 mg/kg(相當于咖啡因2.5 mg/kg),每24 h進行1次緩慢靜脈輸入,若病程中出現呼吸暫停加重,可再次給予10 mg/kg,并將維持量增加20%;于校正胎齡34~36周,生后5~7 d無呼吸暫停發(fā)生,則停用。
65例早產兒中發(fā)生BPD 19例,BPD的診斷及分度參照文獻[14]:任何氧依賴(FiO2>21%)超過28 d的患兒即可診斷BPD,<32周早產兒于糾正胎齡36周時評估BPD嚴重程度,分為輕度(未用氧)、中度(FiO2<30%)、重度(FiO2≥30%或需要機械通氣、CPAP輔助通氣)。
1.2 免疫熒光技術檢測SIRT1定位 各組患兒于入院時(0 d)、7、14、28 d取臨床化驗后廢棄用血1 ml,利用Ficoll密度梯度離心法分離單個核細胞(PBMCs)。細胞涂片和固定,滴入80 μl 0.2% TritonX-100 室溫下打孔15 min,滴加正常山羊血清工作液,滴加抗體稀釋液稀釋的兔抗人SIRT1抗體(1∶80),4 ℃濕盒孵育過夜,避光滴加FITC標記山羊抗小兔IgG (1∶800),37 ℃孵育1 h,DAPI 染核,PBS 洗滌3 次,抗熒光淬滅劑封片,用激光共聚焦顯微鏡檢測:紅色熒光(激發(fā)波長488 nm)反映SIRT1定位情況;DAPI所染細胞核為藍色熒光,兩種熒光合成為紫色熒光。應用Image Pro Plus中的Line profile測定細胞內紫色熒光分布,出現紫色熒光在細胞核中分布減少且出現在細胞核外緣即為發(fā)生SIRT1轉位。取8個視野,以陽性細胞數占細胞總數的構成比計算轉位率。
1.3 Western blot檢測SENP1、細胞核及細胞質中SIRT1、乙酰化p53蛋白 提取各組各時間點PBMC的胞核與胞漿蛋白(操作按說明書進行),進行SDS-PAGE電泳,將目的蛋白轉移至PVDF 膜上,用5%脫脂牛奶封閉液室溫封閉1 h,加入1∶500的一抗(內參一抗終濃度為1∶1 000),4 ℃孵育過夜。TBST洗膜后加入1∶1 000二抗溫育1.5 h。TBST洗膜后加入ECL曝光液(A、B液 1∶1混勻后均勻覆蓋在整片膜上,反應2 min后曝光檢測。采用UVP凝膠圖像處理系統(tǒng)Lakworks 4.6 軟件分析目的條帶的灰度值,結果以目的條帶與內參之比表示。
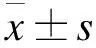
2 結果
2.1 免疫熒光檢測SIRT1定位 SIRT1在細胞質及細胞核中均有表達,但以后者為主。四組中氧暴露第7、14、28天時,SIRT1的轉位率以高氧組最高,且隨氧暴露時間推移而增強;而高氧+咖啡因組在14、28 d時,SIRT1的轉位率降低。見圖1、表1。
2.2 SENP1、細胞核及細胞質中SIRT1、乙酰化p53 蛋白表達的檢測
2.2.1 四組不同時間點SENP1含量比較 第7、14、28天時,SENP1蛋白表達以高氧組最高,且隨氧暴露時間推移而增強;而高氧+咖啡因組在第14、28天時,SENP1蛋白表達降低,降低程度達對照組水平。見表2、圖2。
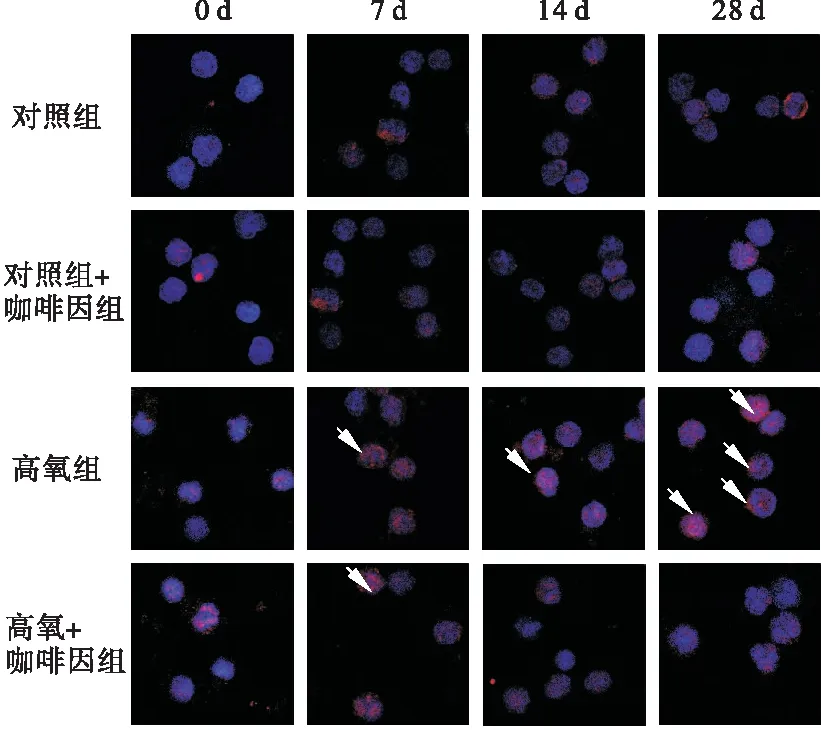
圖1 各組不同時間點的SIRT1轉位情況(熒光顯微鏡,400×)
2.2.2 四組不同時間點細胞核內SIRT1含量比較 第7、14、28天時,細胞核內SIRT1蛋白表達以高氧組最低,且隨氧暴露時間推移而明顯降低;而高氧+咖啡因組在第14、28天時,細胞核內SIRT1蛋白表達升高。見圖3、表3。
2.2.3 四組不同時間點細胞質內SIRT1含量比較 第7、14、28天時,細胞質內SIRT1蛋白表達以高氧組最高,且隨氧暴露時間推移而逐漸升高;而高氧+咖啡因組在第14、28天時,細胞質內SIRT1蛋白表達降低。見圖4、表4。
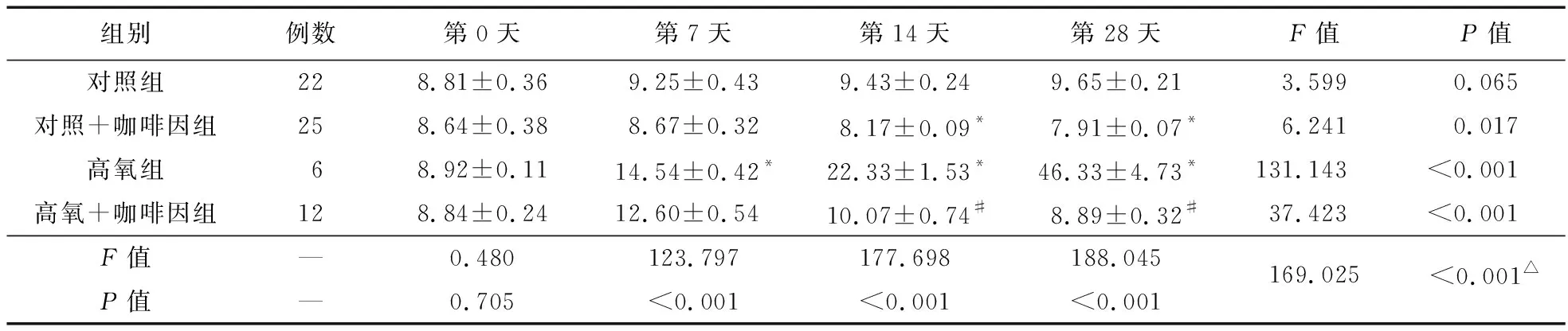
表1 各組PBMCs中不同時間點的SIRT1轉位率比較(%)
注:與對照組比較,*P<0.05;與高氧組比較,#P<0.05;△不同組別不同時間點存在交互效應
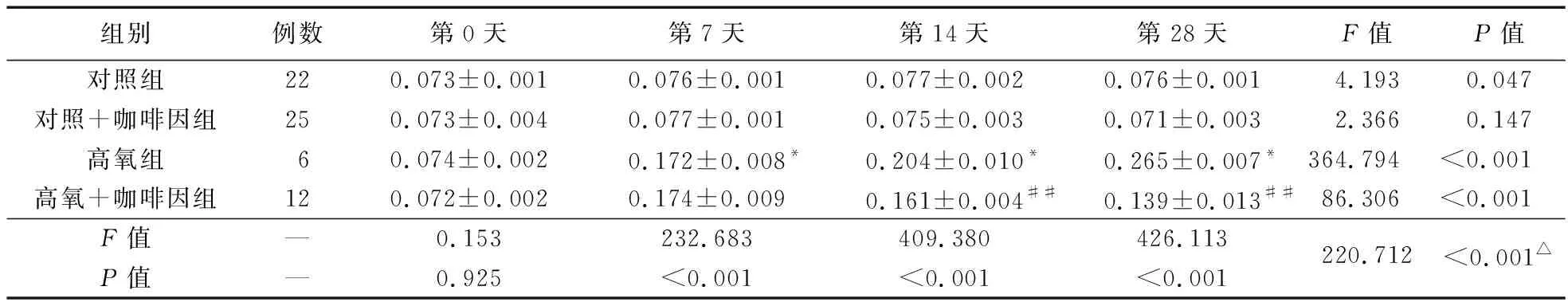
表2 各組PBMCs中不同時間點的 SENP1含量比較
注:與對照組比較,*P<0.05;與高氧組比較,##P<0.01;△不同組別不同時間點存在交互效應
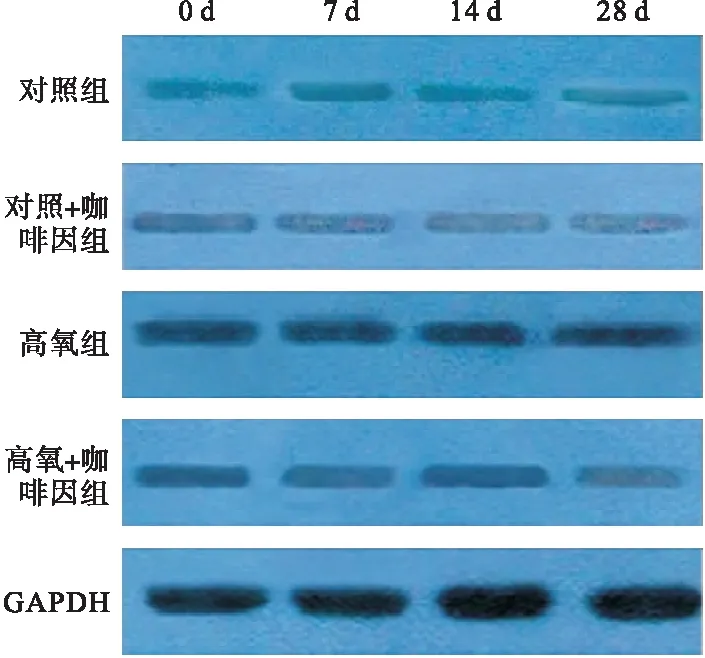
圖2 Western blot檢測四組不同時間點SENP1含量
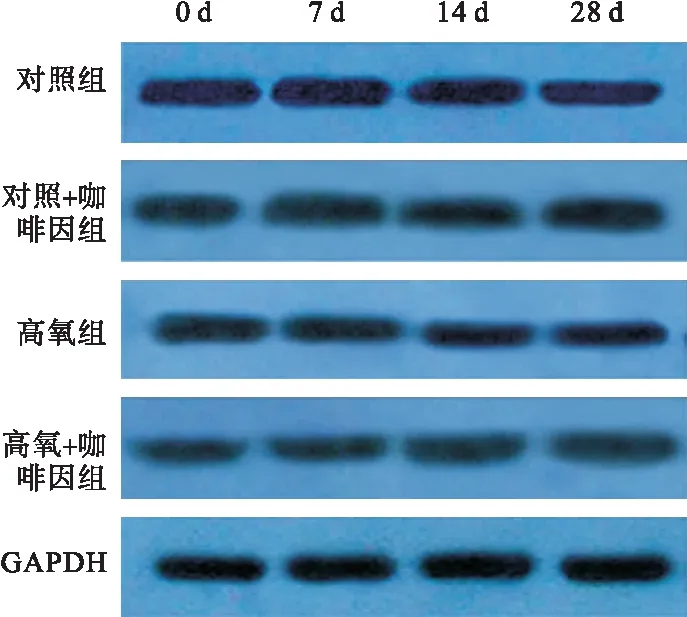
圖3 Western blot檢測四組不同時間點細胞核內SIRT1含量
2.2.4 四組不同時間點乙酰化p53含量比較 四組中第7、14、28天時,乙酰化p53蛋白表達以高氧組最高,且隨氧暴露時間推移而逐漸升高;而高氧+咖啡因組在14、28 d時,出現乙酰化p53蛋白表達降低。見表5、圖5。
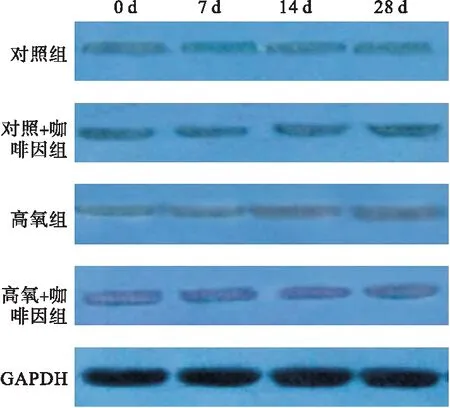
圖4 Western blot檢測四組不同時間點細胞質內SIRT1含量
2.3 各組BPD發(fā)生率比較 對照組、對照組+咖啡因組沒有BPD發(fā)生,高氧組、高氧+咖啡因組BPD的發(fā)生率分別為83%和25%,兩組比較差異有統(tǒng)計學意義(χ2=4.938,P<0.05)。見表6。
3 討論
隨著早產兒數量越來越多,胎齡越來越小,BPD的發(fā)生率越來越高。BPD在1967年由Northway首次報道并命名。隨著國內外學者研究的深入,已明確其病因是氧中毒、感染/炎癥、機械通氣在不成熟的肺組織造成的損傷及異常修復[14]。早期的高氧暴露、機械通氣所致的氣壓傷、
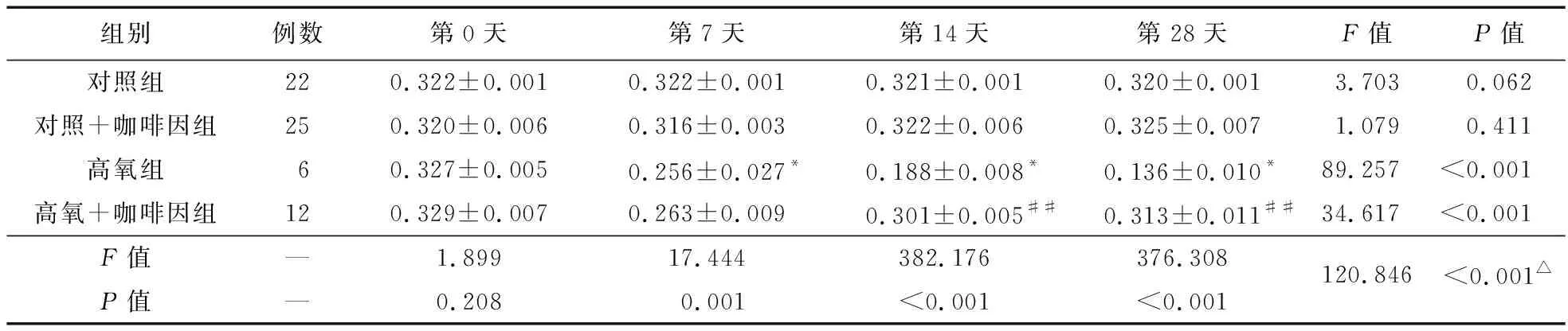
表3 各組PBMCs中不同時間點細胞核內SIRT1的含量比較
注:與對照組比較,*P<0.05;與高氧組比較,##P<0.01;△不同組別不同時間點存在交互效應
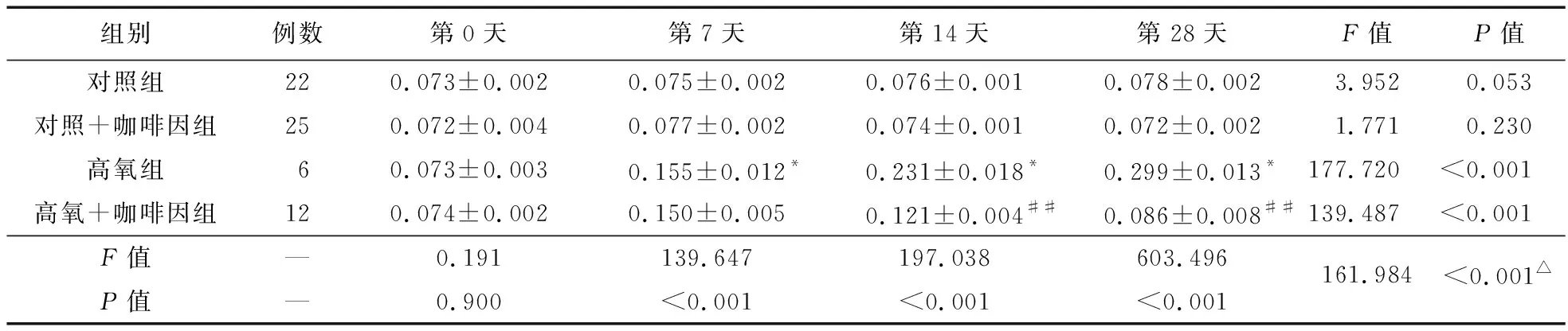
表4 各組PBMCs中不同時間點細胞質內SIRT1的含量比較
注:與對照組比較,*P<0.05;與高氧組比較,##P<0.01;△不同組別不同時間點存在交互效應
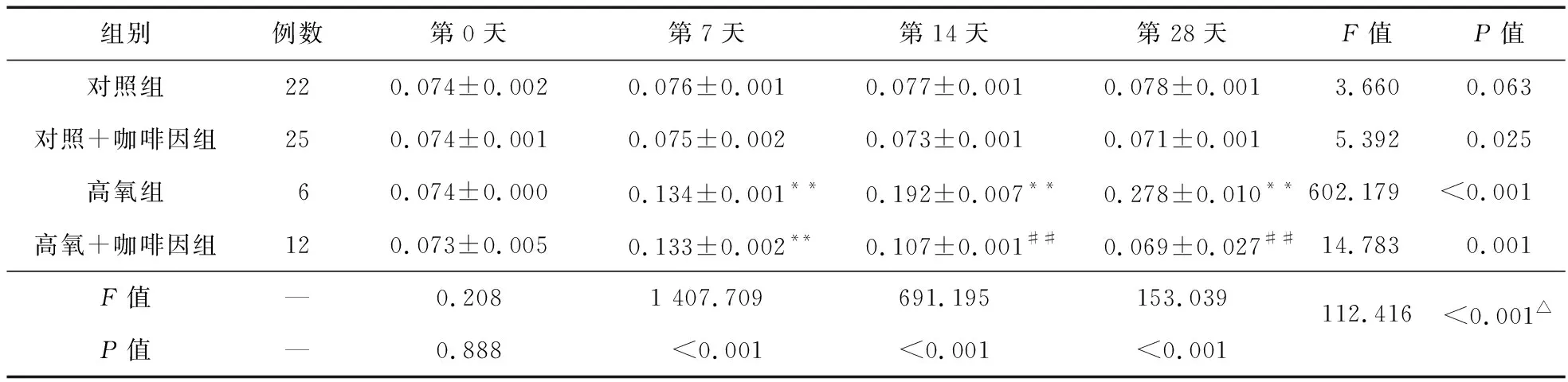
表5 各組PBMCs不同時間點乙酰化p53的含量比較
注:與對照組比較,**P<0.01;與高氧組比較,##P<0.01;△不同組別不同時間點存在交互效應
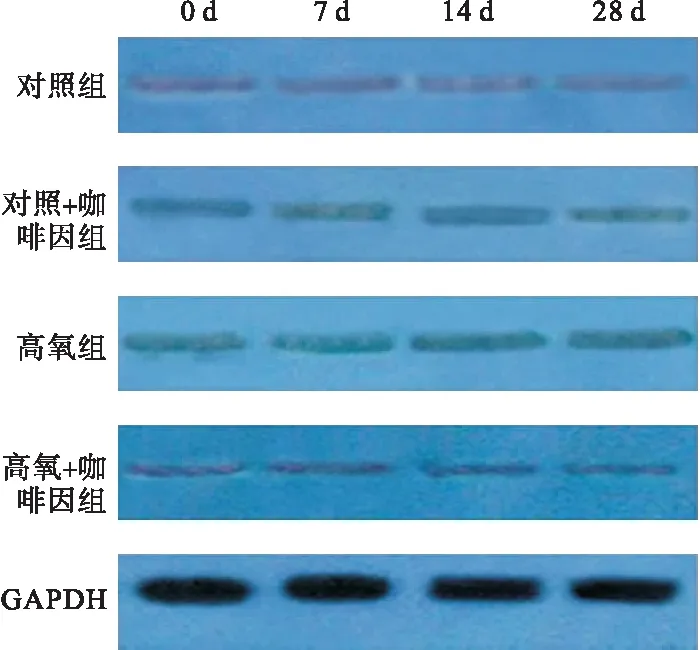
圖5 Western blot檢測四組不同時間乙酰化p53含量

組別例數BPD對照組220對照+咖啡因組250高氧組65(83)高氧+咖啡因組122(25)#
注:#與高氧組比較,χ2=4.938,P<0.05
容量傷都具有促炎作用,早期即迅速產生大量的炎癥因子,如IL-1β、IL-6、IL-8、MCP-1、TNF-α,而抗炎因子IL-10的增長速度較促炎因子慢,促炎因子與抗炎因子的失衡使肺組織發(fā)生炎癥反應,肺泡及肺部毛細血管的上皮細胞纖維化,導致肺損傷,這與BPD的發(fā)生密切相關[15]。BPD的存在可以產生慢性氧依賴,部分患兒需要長期家庭氧療,且呼吸系統(tǒng)疾病發(fā)生率、呼吸道藥物使用率、再次住院率均高于非BPD患兒,可以出現肺功能受損[16]。因此,BPD的發(fā)生不僅使早產兒的死亡率升高,而且降低早產兒的生存質量,增加家庭負擔。
咖啡因屬甲基黃嘌呤類藥物,非特異性腺苷受體阻滯劑,通過非特異性阻斷腺苷A1及A2a受體,導致去甲腎上腺素、多巴胺、5-羥色胺、乙酰膽堿與谷氨酸等神經遞質釋放[17]。在過去的30多年被用于預防和治療早產兒呼吸暫停的過程中,已發(fā)現咖啡因較氨茶堿在BPD、腹脹、心率增快、喂養(yǎng)不耐受、胃潴留的發(fā)生率低[18]。咖啡因的使用可以改善新生兒期肺功能情況,可能與咖啡因使用后可以增加對二氧化碳的敏感性,從而興奮呼吸有關,同時,咖啡因有支氣管擴張作用,可減少周期性呼吸,增加膈肌的活動,減少缺氧性呼吸抑制[19]。國外一篇Meta分析也表明,在極低出生體重兒中早期使用咖啡因(生后3 d內)有益于降低BPD的發(fā)生率,眾多回顧性資料分析也得出應用咖啡因可以降低BPD發(fā)生率的結論[8,12,20-21]。
從BPD的發(fā)病過程可見,生后7~14 d是BPD的進展階段,生后(28±7) d是確診BPD的治療階段。從本實驗結果中可以看出,高氧組中SENP1蛋白升高、細胞核中SIRT1蛋白降低,細胞質中SIRT1蛋白升高、乙酰化p53升高,結合免疫熒光結果,說明高氧可以促進SENP1表達,促進SIRT1從細胞核向細胞質中的核漿轉位,提高p53的乙酰化水平,這與目前的研究結果一致,同時印證了處于BPD的進展階段以后,這些蛋白水平的變化與BPD的發(fā)生緊密相關。
第14天、第28天,高氧+咖啡因組與高氧組比較,SENP1表達降低,細胞核中SIRT1表達升高,細胞質中SIRT1表達降低、乙酰化p53降低,結合免疫熒光結果中SIRT1的轉位率降低,說明咖啡因的使用降低SENP1表達、抑制SIRT1從細胞核向細胞質的核漿轉位,降低p53乙酰化水平,目前研究表明,細胞核中的SIRT1對機體具有保護作用,具有抗凋亡作用,而細胞質中的SIRT1則來源于細胞核中,具有促凋亡作用[22],SIRT1是p53負調控因子,可以在K382位點催化p53的去乙酰化,使用咖啡因后,細胞核內SIRT1蛋白表達增加,使p53的去乙酰化作用增強,同時,乙酰化作用減弱,抵抗了乙酰化p53的促凋亡作用,從而在一定程度上減輕了高氧所致的肺損傷[5,23-24]。
本實驗中,在咖啡因的使用上,均采用首次負荷劑量20 mg/kg 30 min內泵入,間隔24 h后使用維持劑量5 mg/kg,但是在病程中若出現呼吸暫停加重,治療上將予以咖啡因加量,但是Tiwari等[12]已在體外實驗中證實,咖啡因在臨床使用劑量下,對細胞周期及活力沒有影響,并具有抗氧化性質,因此,臨床中咖啡因劑量的調整不會對該通路造成影響。
綜上所述,咖啡因降低高氧條件下BPD的發(fā)生率,其機制是通過降低SENP1表達、抑制SIRT1核漿穿梭、降低p53乙酰化水平,為咖啡因的使用提供了理論依據。
參考文獻:
[1] Jobe AH,Bancalari E.Bronchopulmonary dysplasia[J].Am J Respir Crit Care Med,2001,163(7):1723-1729.
[2] Ryter SW,Choi AM.Regulation of autophagy in oxygen-dependent cellular stress[J].Curr Pharm Des,2013,19(15):2747-2756.
[3] Yang Y,Fu W,Chen J,et al.SIRT1 sumoylation regulates its deacetylase activity and cellular response to genotoxic stress[J].Nat Cell Biol,2007,9(11):1253-1262.
[4] 楊熙,董文斌,李清平,等.早產兒氧暴露后外周血單個核細胞活性氧增加使SIRT1產生核質穿梭[J].細胞與分子免疫學雜志,2015,31(12):1669-1672,1676.
[5] Raz L,Zhang QG,Han D,et al.Acetylation of the pro-apoptotic factor,p53 in the hippocampus following cerebral ischemia and modulation by estrogen[J].PLoS One,2011,6(10):e27039.
[6] 王霞,董文斌,李清平,等.高氧對人肺泡上皮細胞凋亡的影響及其作用機制[J].山東醫(yī)藥,2016,56(3):14-16.
[7] 劉興鈴,董文斌,李清平,等不同濃度氧暴露對早產兒外周血單個核細胞SIRT1通路的影響[J].山東醫(yī)藥,2017,57(14):31-36.
[8] Taha D,Kirkby S,Nawab U,et al.Early caffeine therapy for prevention of bronchopulmonary dysplasia in preterm infants[J].J Matern Fetal Neonatal Med,2014,27(16):1698-1702.
[9] Lodha A,Seshia M,McMillan DD,et al.Association of early caffeine administration and neonatal outcomes in very preterm neonates[J].JAMA Pediatr,2015,169(1):33-38.
[10]Park HW,Lim G,Chung SH,et al.Early caffeine use in very low birth weight infants and neonatal outcomes:a systematic review and Meta-analysis[J].J Korean Med Sci,2015,30(12):1828-1835.
[11]Endesfelder S,Zaak I,Weichelt U,et al.Caffeine protects neuronal cells against injury caused by hyperoxia in the immature brain[J].Free Radic Biol Med,2014,67:221-234.
[12]Tiwari KK,Chu C,Couroucli X,et al.Differential concentration-specific effects of caffeine on cell viability,oxidative stress,and cell cycle in pulmonary oxygen toxicity in vitro[J].Biochem Biophys Res Commun,2014,450(4):1345-1350.
[13]杜立中.早產兒呼吸暫停的藥物治療[J].中國實用兒科雜志,2015,30(2):88-92.
[14]Costeloe KL,Hennessy EM,Haider S,et al.Short term outcomes after extreme preterm birth in England:comparison of two birth cohorts in 1995 and 2006 (the EPICure studies)[J].BMJ,2012,345:e7976.
[15]Schneibel KR,Fitzpatrick AM,Ping XD,et al.Inflammatory mediator patterns in tracheal aspirate and their association with bronchopulmonary dysplasia in very low birth weight neonates[J].J Perinatol,2013,33(5):383-387.
[16]韓明珊,謝辰儀,黃花榮,等.支氣管肺發(fā)育不良早產兒呼吸系統(tǒng)疾病預后的研究進展[J].中華婦幼臨床醫(yī)學雜志:電子版,2016,12(5):617-620.
[17]唐曉艷,王丹華.咖啡因在新生兒中的應用進展[J].中國新生兒科雜志,2014,29(5):343-346.
[18]孔凡玲.枸櫞酸咖啡因治療早產兒呼吸暫停臨床應用觀察[J].臨床醫(yī)學研究與實踐,2016,1(10):38.
[19]Adzikah S,Maletzki J,Rüegger C,et al.Association of early versus late caffeine administration on neonatal outcomes in very preterm neonates[J].Acta Paediatr,2017,106(3):518.
[20]Dobson NR,Patel RM,Smith PB,et al.Trends in caffeine use and association between clinical outcomes and timing of therapy in very low birth weight infants[J].J Pediatr,2014,164(5):992-998.e3.
[21]Patel RM,Leong T,Carlton DP,et al.Early caffeine therapy and clinical outcomes in extremely preterm infants[J].J Perinatol,2013,33(2):134-140.
[22]Tajbakhsh N,Sokoya EM.Regulation of cerebral vascular function by sirtuin 1[J].Microcirculation,2012,19(4):336-342.
[23]Langley E,Pearson M,Faretta M,et al.Human SIR2 deacetylates p53 and antagonizes PML/p53-induced cellular senescence[J].EMBO J,2002,21(10):2383-2396.
[24]Vaziri H,Dessain SK,Ng EE,et al.hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase[J].Cell,2001,107(2):149-159.
