RNAi沉默CAPN1基因對Aβ 25-35誘導的原代神經細胞的神經毒性及凋亡的影響
2018-07-04 11:45:42楊保平唐震宇
中風與神經疾病雜志 2018年6期
關鍵詞:檢測
楊保平, 羅 拯, 鄒 可, 唐震宇
阿爾茨海默病(Alzheimer disease,AD)是一種漸進性不可逆的以神經變性為特征的疾病,其主要病變包括大量的β-淀粉樣蛋白沉積(β-amyloid protein,Aβ)形成的神經炎性斑塊、由微管相關蛋白tau蛋白異常過度磷酸化聚集而引起的神經纖維纏結和選擇性的神經元凋亡,β淀粉樣蛋白(Aβ)沉積及其神經毒性級聯反應被認為在阿爾茨海默病的發病過程中起到至關重要的起始及樞紐作用[1]。在諸多的發病假說中,不溶解的Aβ對神經元有毒性作用,是構成老年斑的核心成分,在導致細胞凋亡的同時,激活膠質細胞誘發生炎癥級聯反應,升高胞內Ca2+濃度,使神經細胞鈣穩態失調[2]。關于阿爾茨海默病發病的分子機制目前還不完全清楚,鈣調失常在該病的發病過程中起了一定作用。有研究表明鈣蛋白酶的過度激活與AD的發病有關[3]。鈣蛋白酶(calpain)是鈣依賴性中性蛋白酶,和caspase同屬半胱氨酸蛋白酶超家族的成員,calpain廣泛分布于哺乳動物神經系統中的胞質、質膜和其他細胞器膜中,正常情況下以無活性的酶原形式存在[4]。該酶不僅參與細胞內正常的信號轉錄,而且參與許多病理過程,包括顱腦損傷、腦缺血等神經元退變[5]。AD病變中,calpain的表達處于失衡狀態[6],而最近的研究表明鈣蛋白酶在AD神經元凋亡的過程中發揮重要的作用[7,8]。
本實驗中利用Aβ 25-35誘導原代胎鼠皮質神經元毒性損傷,旨在探討CAPN1 RNAi對Aβ 25-35誘導的原代神經細胞的神經毒性及凋亡的影響,為阿爾茨海默病的治療提供了新的靶點。
1 資料與方法
1.1 材料和試劑 CCK8試劑盒,generay Trizol RNA提取試劑,takara反轉錄試劑盒,全式金qPCR試劑盒,羅氏AnnexinV/PI凋亡檢測試劑盒,Thermo蛋白提取試劑,CAPN1、CDK5、GSK3β、tau、PHT-tau及β-actin一抗和二抗,慢病毒介導μ-calpain 基因(吉馬)。
1.2 CAPN1 shRNA 重組慢病毒/腺病毒載體構建(neuro-2a-CAPN1 shRNA)篩選 μ-calpain基因有效干擾片段的篩選、質粒載體構建、慢病毒及腺病毒包裝由吉馬基因公司完成。
1.3 原代皮質神經元細胞培養 原代皮質神經元細胞來源于C57BL/6野生型胎鼠,細胞計數后接種到預先用0.1 g/L的多聚賴氨酸包被的24孔板(5×104個細胞/孔),置于含5%CO2的37 ℃恒溫培養箱,神經干細胞B-27培養基培養,含2 mmol/L-谷氨酰胺、105U/L青霉素和100 mg/L鏈霉素。每3 d半量換液一次至原代培養7 d后進行后續試驗。
1.4 Aβ藥物干預方法 用PBS溶解Aβ25-35濃度為10 μmol,置于-80 ℃凍存,使用前置于37 ℃老化7 d。實驗分組:(1)原代胎鼠皮質神經元+PBS;(2)原代胎鼠皮質神經元+Aβ25-35;(3)原代胎鼠皮質神經元+CAPN1 shRNA慢病毒+Aβ25-35;(4)原代胎鼠皮質神經元+CAPN1 shRNA慢病毒+Aβ25-35+P25;取對數生長期的細胞制成單細胞懸液,調整細胞濃度105個/ml,接種至6孔板內,置5%CO2,37 ℃培養箱內培養,細胞達80%匯合給予Aβ25-35等處理,培養72 h后收集細胞或細胞培養液進行各項實驗。
1.5 CCK-8法觀察各組細胞活力的變化 按CCK-8試劑盒說明書進行操作,用胰酶消化對數生長的原代胎鼠神經元細胞,去消化液,加入含胎牛血清的細胞培養液3 ml,將細胞吹打成懸液,1500 r/m 2 min離心,棄上清,用完全培養液重懸,濃度為1×105μmol/ml,接種于96孔板中,于5%CO237 ℃培養箱中培養。細胞生長至70%融合度時用CAPN1shRNA或陰性對照轉染48 h,隨后相應加Aβ25-35、p25處理72 h。用PBS洗滌細胞并用含有CCK8的新鮮DMEM代替,然后將細胞在5%CO2于37 ℃溫育4 h。去培養基后,將DMSO加入各孔中,在酶標儀上測定450 nm處的吸光度值(OD值),重復5次。
1.6 細胞凋亡檢測 取對數生長的原代胎鼠神經元細胞消化,離心后用完全培養液重懸,濃度為1×105μmol/ml,接種于6孔板中,實驗分組處理如前所述。各組細胞經處理培養結束后,按照羅氏試劑盒說明書步驟進行Annexin V和PI的雙染,然后用流式細胞儀檢測凋亡。經胰蛋白酶消化、吹打使其成單細胞懸液,移入離心管中,于1000 r/min離心5 min,棄上清,PBS漂洗一次后,重復離心洗滌一次。將細胞重懸于500 μl binding buffer中,再分別加入5 μl的AnnexinV溶液和5 μl PI染液,輕輕渦旋樣品并在25 ℃在黑暗中溫育15 min。在1 h內轉至流式檢測管,流式細胞檢測儀讀數,實驗重復3次。
1.7 實時熒光定量PCR檢測μ-calpain的表達變化 RNA提取試劑盒提取細胞總RNA,用蛋白核酸分析儀測定RNA的純度及定量,1%瓊脂糖凝膠電泳鑒定RNA完整性。RNA反轉錄成cDNA,反轉錄產物-20 ℃保存或直接進入下一步。TaqMan熒光探針技術行實時熒光定量PCR檢測,結果以β-actin作為內參基因,△△Ct法分析基因的相對表達。
1.8 Western blot方法檢測細胞內相關蛋白的表達 培養神經元經按實驗要求處理后,提取細胞總蛋白,采用BCA法進行蛋白濃度測定分別檢測μ-calpain、CDK5、GSK-3β、tau、PHT-tau表達水平的變化。Western blot按常規操作進行:SDS-PAGE電泳分離蛋白質后轉膜,用Blocking Buffer封閉,緩慢搖動孵育轉印膜2 h。封閉后,用1×TBS洗滌3次,每次5 min。加入稀釋好的一抗,4 ℃孵育過夜。用1×TBST洗滌3次,每次10 min。加入稀釋好的二抗,室溫孵育2 h。用1×TBST洗滌3次,每次10 min。用ECL底物對PVDF膜進行化學發光檢測,使用LAS4000 mini化學發光成像儀進行拍照,并使用Quantity One軟件進行灰度分析定量。
2 結 果
2.1 CAPN1 RNAi對神經元細胞活力的影響 與NC組相比,在加入Aβ誘導后,NC+Aβ組的細胞活力顯著下降(P<0.05),而轉染CAPN1 shRNA后,與模型組相比,CAPN1 shRNA+Aβ組細胞活力顯著增高(P<0.05)(見圖1),表明CAPN1基因對細胞活力有顯著影響,在加入P25后,CAPN1 shRNA+Aβ+P25組細胞活力無顯著性差異(P>0.05)。
2.2 CAPN1 RNAi對神經元細胞凋亡的影響 與NC組相比,NC+Aβ組的細胞早期凋亡率顯著升高(P<0.01),而轉染CAPN1 shRNA后,與模型組相比,CAPN1 shRNA+Aβ組早期凋亡率顯著降低(P<0.01)(見圖2),表明CAPN1基因對神經元細胞凋亡有顯著影響。
2.3 CAPN1 RNAi對神經元細胞CAPN1 mRNA表達水平的影響 與NC組相比,CAPN1 shRNA+Aβ組中CAPN1 mRNA的表達顯著升高(P<0.01);而轉染CAPN1 shRNA后,CAPN1 mRNA的表達得到顯著抑制(P<0.01)(見圖3),表明CAPN1 shRNA轉染成功。
2.4 CAPN1 RNAi對神經元細胞相關蛋白表達水平的影響 Western blot方法檢測蛋白表達情況:與NC組相比,NC+Aβ組中μ-calpain、CDK5、p-tau/tau蛋白表達水平顯著升高(P<0.01),GSK3β蛋白表達水平升高(P<0.05);而轉染CAPN1 shRNA后,與模型組相比,μ-calpain、GSK3β、p-tau/tau蛋白表達水平均顯著降低(P<0.01),CDK5蛋白表達水平降低(P<0.05),表明CAPN1基因對神經元細胞CDK5、GSK3β、p-tau/tau的蛋白表達有顯著影響,在加入P25后,CAPN1 shRNA+Aβ+P25組,CDK5、GSK3β、p-tau/tau蛋白表達水平均有所上調,μ-calpain蛋白表達未見明顯改變(P>0.05)(見圖4),表明CAPN1基因對神經元細胞CDK5、GSK3β、p-tau/tau的蛋白表達有顯著影響。
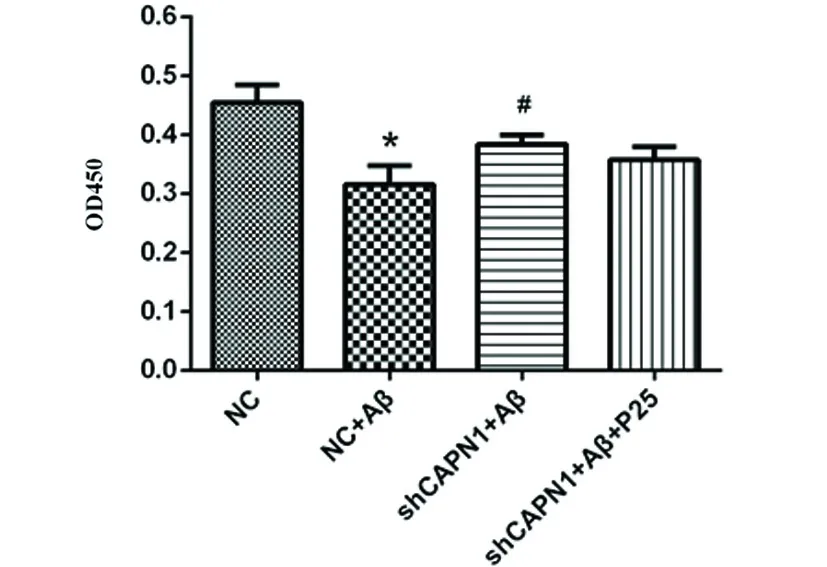
與對照組相比*P<0.05;與模型組相比#P<0.05
圖1 CAPN1 RNAi對神經元細胞活力的影響
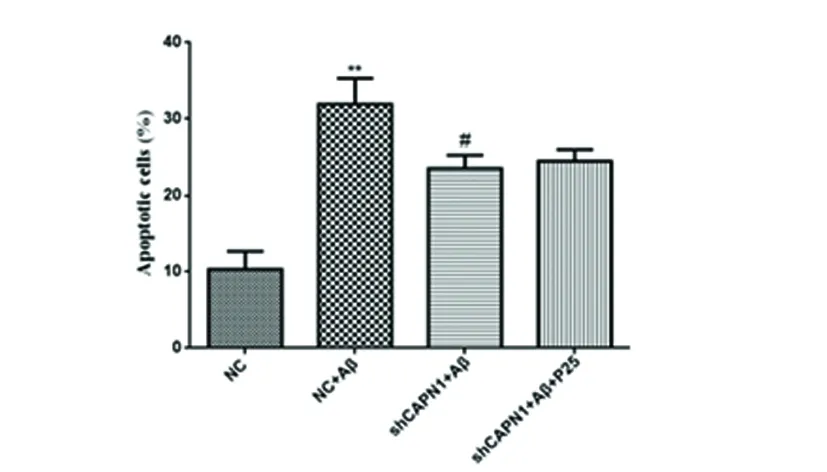

與對照組相比**P<0.01;與模型組相比#P<0.01
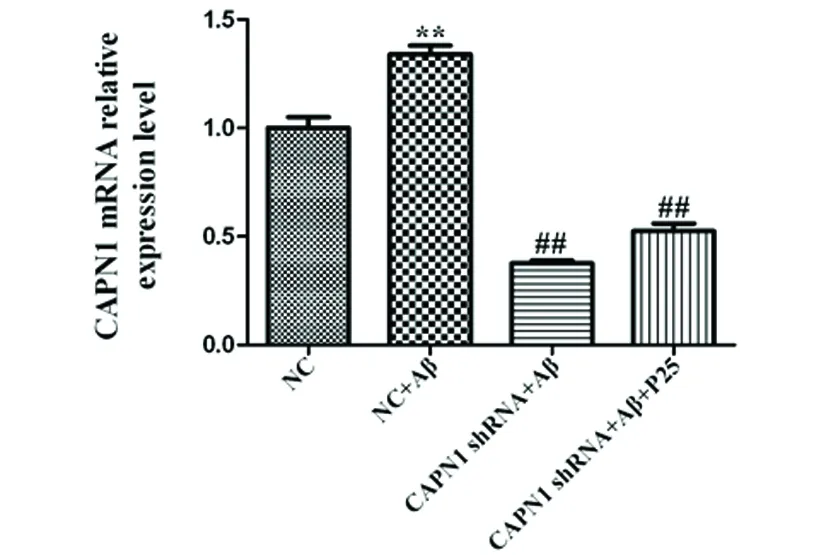
與對照組相比**P<0.01;與模型組相比##P<0.01
圖3 RT-PCR檢測CAPN1 RNAi對神經元細胞CAPN1 mRNA表達水平的影響

與對照組相比**P<0.01;與模型組相比##P<0.01;與shRNA CAPN1+Aβ組相比*P<0.05
圖4 Western blot檢測CAPN1 RNAi對神經元細胞相關蛋白表達水平的影響
3 討 論
阿爾茨海默病是一種以記憶力下降和認知功能障礙為主要表現的神經退行性疾病,以細胞形成大量由β-淀粉樣蛋白組成的淀粉樣斑塊,細胞內神經原纖維纏結及神經元退行性病變。目前多數學者認為AD的病因和發病機制關鍵的因素是Aβ異常代謝和沉積。Aβ具有神經毒性作用,其聚集沉淀后可引起AD樣病變[9],本實驗利用Aβ25-35片段誘導AD細胞模型。研究發現AD與細胞凋亡關系密切,認為細胞凋亡是AD中神經元退行性死亡的重要途徑[10]。干擾細胞凋亡可能成為治療AD新的方法。
研究發現,Ca2+濃度在AD患者腦中升高。近年來有許多研究證據表明,由Ca2+濃度失衡引起的Calpain過度激活直接或間接與AD的大腦病變有關[11]。Calpain系統(包括CalpainⅠ和CalpainⅡ)是一個復雜的、高度調控的蛋白降解體系[12]。在體內,Calpain通過Ca2+激活及自溶而表現出蛋白水解酶活性,并通過鈣蛋白酶抑制蛋白(Calpastatin)抑制Calpain活性。因為Calpain主要作用對象是細胞骨架蛋白,受體蛋白以及蛋白激酶,Calpain可能通過對這些蛋白進行特異的局部降解而對其結構、功能進行調控,在AD疾病中使神經元細胞分解引起凋亡,進而引起神經退行性病變[13]。本研究中發現轉染CAPN1 shRNA后,模型組中CAPN1在mRNA及蛋白水平的表達均顯著下降,與已有報道[14]一致,在AD病變中,calpain過度表達在細胞骨架損傷進展和神經退變中發揮著重要的作用。
本實驗中還發現,轉染CAPN1 shRNA后,神經元細胞的活力明顯增加,凋亡情況明顯改善,并且CDK5、GSK3β、p-tau蛋白表達水平均顯著下調,在加入P25后,CDK5、GSK3β、p-tau的蛋白表達顯著升高。已有研究表明,tau蛋白的過度磷酸化可導致微管功能喪失,是AD分子病理的重要環節,過度磷酸化的tau蛋白通過與正常tau蛋白競爭性結合管蛋白,從而阻斷微管蛋白的組裝,抑制微管聚集,使微管解體及細胞骨架破壞[15]。CDK5是引起tau蛋白磷酸化的重要的蛋白激酶[16]。有多種調節蛋白參與此過程,其中以p35的作用最為重要。在一些損傷因素的作用下,p35可經中性蛋白酶(calpain)作用,降解為p25[17],從而形成p25/CDK5復合體,更大地發揮激活CDK5的作用,進一步對其底物tau蛋白磷酸化,從而引發神經退行性改變[18]。糖原合成酶激酶(GSK)-3β是一種通過磷酸化作用的糖原合成的關鍵調節酶[19]。GSK-3β是通過磷酸化而失活的蛋白激酶之一,磷酸化后其活性明顯降低,GSK-3β被認為參與了有淀粉樣前體蛋白(APP)生成Aβ以及tau蛋白磷酸化過程。近來研究結果顯示,GSK-3β能使tau蛋白多個位點磷酸化,有研究結果顯示:GSK-3β極有可能是AD腦中引起tau蛋白異常磷酸化的最關鍵的激酶之一[20]。本實驗沉默CAPN1基因后,calpain1表達下調,致使GSK-3β表達降低。該結果為阿爾茨海默癥的治療靶點提供了理論基礎。
綜上所述,本研究證明CAPN1 RNAi可以顯著改善Aβ 25-35誘導的原代神經細胞的神經毒性及凋亡,其作用機制可能是通過下調CDK5、GSK3β、p-tau蛋白表達水平來實現的。
[參考文獻]
[1]Palop JJ,Mucke L.Amyloid-beta-induced neuronal dysfunction in Alzheimer’s disease:from synapses toward neural networks[J].Nat Neurosci,2010,13(7):812-818.
[2]Workgroup AA.Calcium Hypothesis of Alzheimer’s disease and brain aging:A framework for integrating new evidence into a comprehensive theory of pathogenesis[J].Alzheimers Dement,2017,13(2):178-182.
[3]Shiraha H,Glading A,Chou J,et al.Activation of m-calpain (calpain II) by epidermal growth factor is limited by protein kinase A phosphorylation of m-calpain[J].Mol Cell Biol,2002,22(8):2716-2727.
[4]Ray SK.Currently evaluated calpain and caspase inhibitors for neuroprotection in experimental brain ischemia[J].Curr Med Chem,2006,13(28):3425-3440.
[5]Ono Y,Saido TC,Sorimachi H.Calpain research for drug discovery:challenges and potential[J].Nat Rev Drug Discov,2016,15(12):854-876.
[6]Biesecker KR,Srienc AI,Shimoda AM,et al.Glial cell calcium signaling mediates capillary regulation of blood flow in the retina[J].J Neurosci,2016,36(36):9435-9445.
[7]Karlsson JO,Blennow K,Janson I,et al.Increased proteolytic activity in lymphocytes from patients with early onset Alzheimer’s disease[J].Neurobiol Aging,1995,16(6):901-906.
[8]謝守嬪,羅 佳,李海龍,等.當歸含藥血清對Aβ-(25-35)誘導PC12細胞凋亡的保護作用研究[J].西部中醫藥,2015,28(6):13-16.
[9]Kudo W,Lee HP,Zou WQ,et al.Cellular prion protein is essential for oligomeric amyloid-beta-induced neuronal cell death[J].Hum Mol Genet,2012,21(5):1138-1144.
[10]Leissring MA,Akbari Y,Fanger CM,et al.Capacitative calcium entry deficits and elevated luminal calcium content in mutant presenilin-1 knockin mice[J].J Cell Biol,2000,149(4):793-798.
[11]Baudry M,Bi X.Calpain-1 and Calpain-2:The Yin and Yang of synaptic plasticity and neurodegeneration[J].Trends Neurosci,2016,39(4):235-245.
[12]Liu Q,Chen Y,Auger-Messier M,et al.Interaction between NFkappaB and NFAT coordinates cardiac hypertrophy and pathological remodeling[J].Circ Res,2012,110(8):1077-1086.
[13]Hou ST,Jiang SX,Desbois A,et al.Calpain-cleaved collapsin response mediator protein-3 induces neuronal death after glutamate toxicity and cerebral ischemia[J].J Neurosci,2006,26(8):2241-2249.
[14]Yu Y,Run X,Liang Z,et al.Developmental regulation of tau phosphorylation,tau kinases,and tau phosphatases[J].J Neurochem,2009,108(6):1480-1494.
[15]Liu SL,Wang C,Jiang T,et al.The role of cdk5 in Alzheimer’s disease[J].Mol Neurobiol,2016,53(7):4328-4342.
[16]Cruz JC,Tseng HC,Goldman JA,et al.Aberrant Cdk5 activation by p25 triggers pathological events leading to neurodegeneration and neurofibrillary tangles[J].Neuron,2003,40(3):471-483.
[17]Zhou M,Huang T,Collins N,et al.APOE4 induces site-specific tau phosphorylation through calpain-CDK5 signaling pathway in EFAD-Tg mice[J].Curr Alzheimer Res,2016,13(9):1048-1055.
[18]Jiang H,Guo W,Liang X,et al.Both the establishment and the maintenance of neuronal polarity require active mechanisms:critical roles of GSK-3 beta and its upstream regulators[J].Cell,2005,120(1):123-135.
[19]Javitt DC.Glutamate as a therapeutic target in psychiatric disorders[J].Mol Psychiatry,2004,9(11):984-997.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
