葛根素對MC3T3-E1成骨前體細胞增殖能力的影響及其機制
2018-06-15 04:34:02馮倩俞鵬程嵩奕曾祥偉楊瑞楊婧趙鳳鳴朱家鵬詹秀琴南京中醫藥大學醫學與生命科學學院南京003南京中醫藥大學第一附屬醫院
山東醫藥 2018年16期
關鍵詞:信號
馮倩,俞鵬,程嵩奕,曾祥偉,楊瑞,楊婧,趙鳳鳴,朱家鵬,詹秀琴(南京中醫藥大學醫學與生命科學學院,南京003;南京中醫藥大學第一附屬醫院)
葛根素是提取自中藥葛根中的一種異黃酮類物質,具有抗癌及改善心腦血管疾病、糖尿病及骨質疏松等作用[1]。研究發現,低濃度葛根素可能通過下調miR-204及其宿主基因TRPM3表達等多種途徑提高成骨細胞增殖能力[2,3]。絲裂原活化蛋白激酶(AMPK)是細胞內的一類絲氨酸/蘇氨酸蛋白激酶,目前已知的AMPK信號通路有三條,包括經典的AMPK信號途徑、ERK信號通路和非經典的AMPK信號途徑(JNK/SAPK途徑和p38/AMPK途徑),三種信號通路往往同時發揮作用,與細胞增殖、分化以及能量代謝密切相關[4]。因AMPK多在磷酸化后發揮生物學效應,故磷酸化的AMPK(p-AMPK)表達水平對信號通路的調控具有重要意義。研究發現,激活AMPK/微管相關蛋白1輕鏈3(LC3)信號通路可通過誘導細胞自噬而促進肺動脈平滑肌細胞、鼻黏膜上皮細胞及多巴胺能神經元[5~7]增殖。目前葛根素對成骨細胞增殖的影響及其是否與AMPK/LC3信號通路有關鮮見報道,為此我們于2017年9~11月進行了如下研究。
1 材料
細胞:MC3T3-E1成骨前體細胞購于中國科學院上海細胞生物學研究所。細胞常規培養于含有10%胎牛血清的α-MEM完全培養基中,5% CO2、37 ℃孵箱中培養,待細胞融合度達90%時,0.25%胰酶消化,按1∶3比例進行傳代培養。主要試劑:α-MEM 完全培養基(上海中喬新舟生物科技有限公司),葛根素標準品(中國藥品生物制品檢定所),β-雌二醇(國藥集團化學試劑有限公司),CCK-8 試劑盒(日本同仁化學研究所),兔抗鼠p-AMPK、LC3多克隆抗體、內參β-actin一抗、Anti-Rabbit IgG二抗(美國Cell Signaling Technology公司),BCA 試劑盒(北京索萊寶科技有限公司),顯色劑(美國BIO-RAD公司),siRNA-AMPK質粒(上海吉瑪制藥技術有限公司)。主要儀器:超凈工作臺(蘇州佳寶凈化工程設備有限公司),多功能酶標儀(美國BioTek公司),凝膠成像儀(美國BIO-RAD公司)。
2 方法與結果
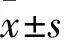
2.1 葛根素促進細胞增殖能力的最佳作用濃度及作用時間篩選 取對數生長期MC3T3-E1細胞,以5×103個/孔接種于96孔板中,37 ℃、5% CO2細胞培養箱中培養。待細胞貼壁后,將細胞隨機分為空白對照組、葛根素組和雌激素組,空白對照組每孔加入α-MEM完全培養基200 μL,葛根素組每孔加入葛根素與α-MEM完全培養基混合液200 μL,調整葛根素終濃度分別為0.1、1、10 μmol/L,雌激素組每孔加入雌激素與α-MEM完全培養基混合液200 μL,調整雌激素終濃度為0.01 μmol/L,每組設置6個復孔。分別于作用24、36、48 h時每孔加入10 μL CCK-8試劑,將細胞板避光置于細胞培養箱中培養30 min,采用多功能酶標儀檢測450 nm波長處的光密度(OD)值。結果顯示,葛根素組不同濃度作用24、36、48 h的細胞增殖能力均高于空白對照組、低于雌激素組(P均<0.05);0.1 μmol/L葛根素作用48 h細胞增殖能力均高于同濃度作用24、36 h及1、10 μmol/L葛根素作用48 h(P均<0.05)。見表1。后續實驗葛根素組均選擇葛根素濃度為0.1 μmol/L、作用時間為48 h。
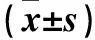
表1 各組細胞增殖能力
2.2 葛根素對細胞自噬的影響 取對數生長期MC3T3-E1細胞,以5×105個/孔接種于6孔板中。待細胞貼壁后,分為空白對照組、葛根素組和雌激素組,各組每孔均加入α-MEM完全培養基2 mL。葛根素組每孔加入葛根素,調整葛根素終濃度為0.1 μmol/L;雌激素組每孔加入雌激素,調整雌激素終濃度為0.01 μmol/L;每組設置3個復孔。作用48 h,每孔加入1 mL胰蛋白酶消化并吸至1.5 mL EP管中,1 000 r/min離心5 min,去上清;每管加入500 μL戊二醛,置于4 ℃冰箱中固定12 h;鋨酸再次固定,醋酸鈾染色,乙醇梯度脫水,樹脂包埋,0.1 μm厚度切片。檸檬酸鉛染色,透射電鏡下觀察各組細胞器結構。結果顯示,空白對照組細胞器、細胞核和染色體形態正常;葛根素組和雌激素組細胞質內可見較多環狀結構物質,該物質具有自噬體特異性的雙層膜結構。表明葛根素與雌激素可誘導MC3T3-E1細胞發生自噬。
2.3 葛根素對細胞p-AMPK、LC3-Ⅱ/LC3Ⅰ表達的影響 取對數生長期MC3T3-E1細胞,參照2.2的方法進行接種及分組處理,提取總蛋白并采用BCA法進行蛋白定量。取樣本25 μg進行SDS-PAGE凝膠電泳,轉膜并剪膜后,加封閉液室溫封閉1 h。TBST漂洗5 min×3次。分別加入p-AMPK、LC3一抗(稀釋比例均為1∶1 000)4 mL,4 ℃過夜,TBST漂洗5 min×3次。加入膜與HRP結合的二抗(稀釋比例均為1∶2 000),室溫下置于搖床孵育1 h,TBST漂洗5 min×3次。將顯色液加于濾膜正面并顯影,調整曝光時間,選擇最佳條帶。采用Bandscan分析軟件分析條帶灰度并進行半定量分析。以目的基因條帶灰度值與管家基因β-actin條帶灰度的比值計算蛋白相對表達量,并計算LC3-Ⅱ/LC3-Ⅰ。結果顯示,葛根素組、雌激素組p-AMPK及LC3-Ⅱ/LC3-Ⅰ均明顯高于空白對照組,雌激素組均明顯高于葛根素組(P均<0.01)。見表2。
表2 各組細胞p-AMPK、IC3-Ⅰ、IC3-Ⅱ、LC3-Ⅱ/LC3-Ⅰ比較
注:與空白對照組比較,*P<0.01;與雌激素組比較,#P<0.01。
2.4 干擾AMPK對葛根素作用后MC3T3-E1細胞自噬及LC3-Ⅱ/LC3-Ⅰ表達的影響
2.4.1 細胞分組處理 取對數生長期MC3T3-E1細胞,以5×105個/孔接種于6孔板,每組3個復孔。細胞融合至70%時,隨機分為空白對照組、陰性對照組、葛根素組和AMPK干擾組。各組每孔均加入轉染試劑與α-MEM完全培養基混合液2 mL。陰性對照組采用Lipofectamine2000脂質體轉染法轉染無意義陰性序列;葛根素組及AMPK干擾組每孔加入葛根素,調整終濃度為0.1 μmol/L,AMPK干擾組采用Lipofectamine2000脂質體轉染法轉染siRNA-AMPK質粒。各組均培養4 h,更換為α-MEM完全培養基。
2.4.2 細胞自噬情況 各組轉染48 h,參照2.2的方法觀察細胞自噬情況。結果顯示,空白對照組和陰性對照組細胞器、細胞核、染色體形態正常;葛根素組細胞質內可見較多環狀結構物質,該物質具有自噬體特異性的雙層膜結構;AMPK干擾組僅見一處自噬體特異性結構,較空白對照組和陰性對照組無明顯區別。表明干擾AMPK表達可抑制葛根素介導的MC3T3-E1細胞自噬。
2.4.3 細胞LC3-Ⅱ/LC3-Ⅰ表達 各組轉染48 h,參照2.3的方法檢測細胞LC3-Ⅱ/LC3-Ⅰ。結果顯示,葛根素組LC3-Ⅱ/LC3-Ⅰ明顯高于空白對照組、陰性對照組、AMPK干擾組(P均<0.01),空白對照組、陰性對照組、AMPK干擾組LC3-Ⅱ/LC3-Ⅰ比較差異無統計學意義(P>0.05)。見表3。
表3 各組細胞LC3-Ⅰ、LC3-Ⅱ及LC3-Ⅱ/LC3-Ⅰ比較
注:與空白對照組及陰性對照組比較,*P<0.01;與葛根素組比較,#P<0.01。
3 討論
骨質疏松癥是以單位體積內骨組織量減少為特征的一類骨代謝病,成骨細胞增殖能力下降是誘發骨質疏松的重要原因[8]。骨質疏松癥在老年人和絕經后婦女人群中高發,多與雌激素缺乏有關[9]。近年研究發現,葛根素具有類雌激素樣作用,在促進成骨細胞增殖、改善骨質疏松方面具有良好的調節作用,但是其具體機制尚未明了[10]。本研究結果顯示,不同濃度葛根素均可促進MC3T3-E1細胞增殖,并呈時間依賴性,其中以0.1 μmol/L葛根素作用48 h效果最佳。
自噬是將自身細胞器包裹,并通過溶酶體降解的過程,生理狀態下可以發揮提供能量、維持細胞器穩態的作用。一旦損傷超過了細胞的承受能力,細胞將通過自噬進行調控性自殺[11]。LC3參與細胞自噬過程,細胞中新合成的LC3被加工成細胞質可溶形式的LC3-Ⅰ,泛素化修飾后與細胞膜上的磷脂酰乙醇胺結合,轉化為細胞膜結合形式的LC3-Ⅱ。LC3-Ⅱ定位于前自噬體和自噬體,是自噬標志物,而LC3-Ⅱ/LC3-Ⅰ與自噬水平呈正相關,可反映細胞自噬水平[12]。目前自噬對成骨細胞增殖水平的影響存在爭議。自噬相關蛋白7 (Atg7) 是一類自噬組成蛋白,可促使LC3-Ⅰ與磷脂酰乙醇胺共價結合,形成LC3-Ⅱ。有研究發現,Atg7敲除小鼠較正常小鼠的骨量降低,成骨細胞增殖能力下降[13]。此外,在成骨細胞分化的過程中,形態學轉變可導致細胞內質網和線粒體數量逐漸減少,而抑制細胞自噬后則不會,表明成骨細胞自噬在一定程度上有助于骨骼穩態和骨形成。本研究結果顯示,MC3T3-E1細胞經0.1 μmol/L葛根素作用48 h后,透射電鏡下可見明顯有自噬體特異性的雙層膜結構,表明低濃度葛根素可通過促進成骨細胞自噬而增強其細胞增殖能力。
成骨細胞的自噬過程涉及多條信號通路。有研究顯示,抑制ERK信號通路可降低大鼠成骨細胞的自噬水平,延緩糖尿病誘導的骨質疏松癥,推測自噬對成骨細胞的不同影響可能與激活自噬的途徑有關[14]。AMPK/LC3通路是近年來自噬相關通路的研究熱點。研究發現,外源性H2S可以通過AMPK/LC3通路誘導自噬,降低糖尿病心肌病患者的氧化應激水平[15]。血府逐瘀湯可以通過AMPK/LC3通路上調缺氧-復氧心肌細胞自噬水平,從而增強細胞增殖能力,減少心肌細胞凋亡,發揮抗缺血再灌注損傷作用[16]。葛根素是否通過AMPK/LC3通路誘導細胞自噬從而產生保護作用目前鮮見報道。本研究結果顯示,MC3T3-E1細胞經0.1 μmol/L葛根素作用48 h后LC3-Ⅱ/LC3-Ⅰ及p-AMPK蛋白表達水平上升,而干擾AMPK表達后的成骨細胞電鏡下自噬體數量減少、LC3-Ⅱ/LC3-Ⅰ明顯降低。表明在葛根素調控自噬參與成骨細胞增殖的過程中,AMPK/LC3通路可能發揮了正向調控作用。但細胞自噬是一個動態過程,且成骨細胞的增殖、分化、礦化對骨質疏松均會產生影響[17],需進一步深入研究其中的分子機制。
綜上所述,葛根素可增強MC3T3-E1細胞增殖能力;激活AMPK/LC3信號通路介導的自噬水平是其可能的機制之一,可為臨床治療骨質疏松癥提供新的治療靶點。
參考文獻:
[1] 曾祥偉,馮倩,張瑩瑩,等.葛根素對炎癥相關疾病研究進展[J].中國藥理學通報,2018,34(1):8-11.
[2] Zhan XQ, Zeng XW, Zhang YY, et al. Puerarin promotes the viability and differentiation of MC3T3 E1 cells by miR 204 regulated Runx2 upregulation[J]. Mol Med Rep, 2017,16(5):6262-6268.
[3] 張瑩瑩,周建斌,曾祥偉,等.葛根素對成骨細胞增殖能力及靶向Runx2的miRNA的影響[J].中國藥理學通報,2016,32(10):1457-1462.
[4] 馮倩,曾祥偉,楊瑞,等.骨髓間充質干細胞成骨分化相關信號通路及其與BMP-2關系的研究進展[J].山東醫藥,2017,57(48):97-99.
[5] Zhou Y, Wang Y, Wang X, et al. The protective effects of Κ-Opioid receptor stimulation in hypoxic pulmonary hypertension involve inhibition of autophagy through the AMPK-MTOR pathway[J]. Cell Physiol Biochem, 2017,44(5):1965-1979.
[6] Wang XH, Zhang ZH, Cai XL, et al. Lipopolysaccharide induces autophagy by targeting the AMPK-mTOR pathway in Human Nasal Epithelial Cells[J]. Biomed Pharmacother, 2017,96:899-904.
[7] Wen Z, Zhang J, Tang P, et al. Overexpression of miR 185 inhibits autophagy and apoptosis of dopaminergic neurons by regulating the AMPK/mTOR signaling pathway in Parkinson′s disease[J]. Mol Med Rep, 2018,17(1):131-137.
[8] Diepenhorst N, Rueda P, Cook AE, et al. G protein-coupled receptors as anabolic drug targets in osteoporosis[J]. Pharmacol Ther, 2017,7258(17):30265-30266.
[9] Almeida M, Laurent MR, Dubois V, et al. Estrogens and androgens in skeletal physiology and pathophysiology[J]. Physiol Rev, 2017,97(1):135-187.
[10] An J, Yang H, Zhang Q, et al. Natural products for treatment of osteoporosis: the effects and mechanisms on promoting osteoblast-mediated bone formation[J]. Life Sci, 2016,147:46-58.
[11] Keulers TG, Schaaf MBE, Rouschop KMA. Autophagy-dependent secretion: contribution to tumor progression[J]. Front Oncol, 2016,6:251.
[12] Dancourt J, Melia TJ. Lipidation of the autophagy proteins LC3 and GABARAP is a membrane-curvature dependent process[J]. Autophagy, 2014,10:1470-1471.
[13] Piemontese M, Onal M, Xiong J, et al. Low bone mass and changes in the osteocyte network in mice lacking autophagy in the osteoblast lineage[J]. Sci Rep, 2016,6:24262.
[14] Zhang WL, Meng HZ, Yang RF, et al. Melatonin suppresses autophagy in type 2 diabetic osteoporosis[J]. Oncotarget, 2016,7(32):52179-52194.
[15] Yang F, Zhang L, Gao Z, et al. Exogenous H2S protects against diabetic cardiomyopathy by activating autophagy via the AMPK/mTOR Pathway[J]. Cell Physiol Biochem, 2017,43(3):1168-1187.
[16] Shi X, Zhu H, Zhang Y, et al. XuefuZhuyu decoction protected cardiomyocytes against hypoxia/reoxygenation injury by inhibiting autophagy[J]. BMC Complement Altern Med, 2017,17(1):325.
[17] 曾祥偉,馮倩,張瑩瑩,等.葛根素對MC3T3-E1細胞增殖、分化和礦化及TRPM3 mRNA表達的影響[J].中國藥理學通報, 2017,33(7):977-982.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
