丹參酮ⅡA磺酸鈉抑制低氧誘導的大鼠肺動脈平滑肌細胞增殖及其機制
2018-06-08 09:07:36楊磊鄭金旭史小東錢海孫金玲
江蘇大學學報(醫學版) 2018年3期
楊磊, 鄭金旭, 史小東, 錢海, 孫金玲
(1. 江蘇大學附屬醫院呼吸內科, 江蘇 鎮江 212001; 2. 江蘇大學醫學院, 江蘇 鎮江 212013)
肺動脈高壓是多因素所致的一種病理生理狀態,包含5種臨床亞組,其預后不良。慢性低氧所致肺動脈高壓(hypoxia-induced pulmonary hypertension,HPH)是肺動脈高壓中最常見的亞組之一,其發病機制與肺動脈中層平滑肌細胞異常增殖引起的肺血管重塑有關[1]。研究揭示,HIF-2α、Wnt、TGF和MAPKs等多種信號通路參與肺動脈平滑肌細胞(pulmonary arterial smooth muscle cells, PASMC)增殖和肺血管重塑,但針對HPH仍缺乏有效的治療藥物[2-5]。近來文獻報道mTOR信號途徑也與PASMC增殖有關,慢性低氧啟動mTOR活化,通過磷酸化激活下游關鍵調節因子eIF2α,進一步增加c-myc表達,從而促進血管平滑肌細胞過度增殖[6]。
丹參酮ⅡA磺酸鈉(sodium tanshinone ⅡA sulfonate,STS)是丹參酮ⅡA的水溶性衍生物,是中藥丹參的主要有效成分之一。研究提示STS可改善肺血管重塑,降低肺動脈壓力,從而治療慢性低氧肺動脈高壓模型大鼠[7]。也有臨床研究結果表明,STS可改善肺動脈高壓患者的臨床癥狀,降低肺動脈壓力[8],但其具體作用機制尚未明確。故本研究通過建立PASMC缺氧模型并予STS干預,檢測細胞內mTOR和eIF2α蛋白的表達,觀察其對低氧誘導的大鼠PASMC增殖的影響,進一步探討STS治療肺動脈高壓的可能機制。
1 材料與方法
1.1 材料
大鼠PASMC購于賽齊(上海)生物工程有限公司(貨號CBR130556);STS凍干粉(成都瑞芬思生物科技有限公司);雷帕霉素(RAM),CCK-8試劑盒(上海生工生物工程有限公司);mTOR抗體、p-mTOR抗體及c-myc抗體(美國Cell Signaling Technology);eIF2α抗體、p-eIF2α抗體(英國Abcam公司);β-肌動蛋白抗體(北京博奧森生物技術有限公司);蛋白裂解液及蛋白酶抑制劑苯甲基磺酰氟(上海博彩生物公司);Trizol(美國Invitrogen公司);DEPC原液(上海生工生物工程有限公司);PrimeScriptTMRT Reagent Kit(日本TaKaRa公司)。倒置光學顯微鏡,CO2培養箱,細胞缺氧培養箱(美國Thermo公司);DU800核酸蛋白定量儀(美國Beckman公司);DNA Engine普通PCR儀及CFX96熒光定量PCR儀(美國Bio-Rad公司)。
1.2 細胞分組及給藥
取PASMC傳代后用于后續實驗,實驗中施加干預時用含10%胎牛血清的DMEM高糖培養基作為基質。在驗證STS抑制低氧誘導的PASMC增殖實驗中,將細胞分為常氧組,常氧+STS組(5、10、20 ng/mL)、低氧組(3%O2)、低氧+STS組(5、10、20 ng/mL)。在STS作用機制實驗(細胞通路實驗)中,行熒光定量PCR檢測時,將PASMC分為常氧組、低氧組、低氧+STS組(10 ng/mL);行蛋白質印跡檢測時,將PASMC分為常氧組、低氧組、低氧+STS組(10 ng/mL)、低氧+RAM組(20 nmol/L)。
1.3 CCK-8法檢測細胞增殖能力
待PASMC生長至80%~90%時,將其消化重懸,以5×103/mL密度接種于48孔板內培養;每組設5個復孔(分組見“1.2”驗證實驗)。培養60 h后用CCK-8試劑盒測定波長450 nm處各孔D值,代表細胞的相對增殖率。
1.4 熒光定量PCR檢測大鼠PASMC中mTOR、eIF2α和c-myc mRNA的表達
取PASMC,去除培養基;用Trizol法提取總RNA,在紫外分光光度計下定量;反轉錄獲得cDNA。采用SYBR-Green法行熒光定量PCR。大鼠eIF2α引物正義鏈序列:5′-GGACAAATGGAAGTATGGGATG-3′,反義鏈序列5′-CAAGAGAGAGCCAGTGTAATGC-3′。大鼠mTOR引物正義鏈序列: 5′-ATCCAGACCCTGACCCAAAC-3′,反義鏈序列:5′-TCCACCCACTTCCTCATCTC-3′。c-myc引物正義鏈序列:5′-TGTCCGTTCAAGCAGATGAG-3′,反義鏈序列:5′-GGGTCAGTTTATGCACCAGA-3′。反應條件:預變性95 ℃ 3 min,95 ℃變性5 s,60 ℃退火15 s,72 ℃延伸15 s,擴增40個循環;運用熔解曲線程序以鑒定擴增產物的特異性。數據采用相對定量的ΔCt法處理,以β-肌動蛋白對mTOR、eIF2α和c-myc mRNA進行標準化分析。
1.5 免疫印跡法測定p-mTOR、mTOR、p-eIF2α、eIF2α、c-myc蛋白的表達
取PASMC,加入預冷PBS洗滌3次;充分吸盡殘余液體,加入100 ∶1配置的蛋白裂解液及蛋白酶抑制劑,搖床搖勻置于冰上裂解30 min;刮取細胞碎片,于4 ℃以12 000×g離心30 min;取上清液進行蛋白定量;加入相同體積的2×SDS-上樣緩沖液,100 ℃煮沸10 min;再超聲破碎細胞10 s。取40 μg蛋白樣品,10%SDS-PAGE電泳1.5 h;200 mA轉移90 min至PVDF薄膜;封閉液中封閉1 h;加入一抗,mTOR、p-mTOR、eIF2α、p-eIF2α及c-myc抗體(稀釋比均1 ∶1 000)4 ℃孵育過夜;洗膜3次,10 min/次;加入堿性磷酸酶標記的抗IgG抗體(1 ∶5 000)室溫孵育1 h;顯色液顯色,半定量分析顯影條帶。
1.6 統計學處理
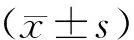
示,多組比較采用單因素方差分析,進一步兩兩比較采用LSD-t檢驗。以P<0.05表示差異有統計學意義。
2 結果
2.1 各組細胞增殖率的比較
與常氧組相比,低氧組細胞增殖率明顯升高(P<0.05);與低氧組相比,低氧+STS(5、10、20 ng/mL)細胞增殖率呈濃度依賴性降低(P<0.05),尤其藥物濃度達20 ng/mL時,細胞增殖水平明顯降低。與常氧組相比,常氧+STS(5、10 ng/mL)細胞增殖水平無明顯降低(P>0.05),但濃度達20 ng/mL時,細胞增殖水平顯著下降(P<0.05),考慮為細胞毒性作用,故選用10 ng/mL STS用于后續實驗。見圖1。
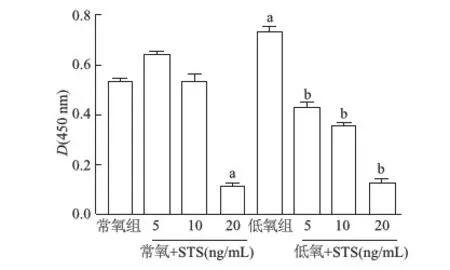
a:P<0.05,與常氧組比較;b:P<0.05,與低氧組比較
2.2 各組細胞mTOR、eIF2α、c-myc mRNA的表達
實時熒光定量PCR結果顯示,與常氧組相比,低氧組mTOR、eIF2α和c-myc mRNA相對表達量明顯升高(P<0.05),低氧+STS組3種mRNA相對表達量較低氧組明顯降低(P<0.05);由此表明,STS處理對于PASMC的mTOR、eIF2α、c-myc mRNA表達有顯著抑制作用。見圖2。

a:P<0.05,與常氧組比較;b:P<0.05,與低氧組比較
2.3 各組細胞中mTOR、p-mTOR、eIF2α、p-eIF2α、c-myc蛋白的表達
免疫印跡結果顯示,與常氧組相比,低氧組細胞中p-mTOR、mTOR、p-eIF2α、eIF2α和c-myc蛋白表達均顯著增加(P<0.05);與低氧組相比,低氧+STS組和低氧+RAM組細胞中p-mTOR、mTOR、p-eIF2α、eIF2α和c-myc蛋白相對表達量均明顯降低(P<0.05)。低氧+STS組與低氧+RAM組間各蛋白表達均無明顯差異(P>0.05)。由此表明,STS干預能夠抑制PASMC中mTOR、eIF2α、c-myc的表達,與雷帕霉素的效果類似,見圖3。
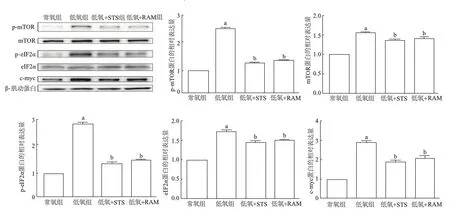
a:P<0.05,與常氧組比較;b:P<0.05,與低氧組比較
3 討論
丹參酮ⅡA作為丹參中的一種提取成分,其水溶性STS在臨床中多用于治療心血管疾病,如高血壓、動脈粥樣硬化等[9]。丹參酮ⅡA具有抑制血小板黏附、聚集,抗血栓作用,還具有抗動脈粥樣硬化、抗心肌缺氧、改善血管平滑肌功能等作用[10]。目前認為肺動脈高壓發病與遠端肺動脈血管中層的PASMC異常增殖導致的肺血管重塑有關[1]。Jiang等[11]發現STS干預慢性低氧性肺動脈高壓大鼠模型,能夠抑制PASMC增殖,改善肺血管重塑,降低肺動脈壓力;Wang等[12]通過構建低氧性肺動脈高壓大鼠和野百合堿誘導肺動脈高壓模型大鼠,并予STS干預,結果發現遠端肺動脈平滑肌經典瞬時受體電位蛋白(TRPC)1、6表達下調,而平滑肌細胞內基礎鈣離子濃度及鈣池操縱性鈣內流增加,導致PASMC增殖及遷移抑制,且右心室收縮壓、右心室平均壓、右室壁厚度與左室壁+室間隔厚度之比均下降,肺血管重塑明顯改善。上述結果表明,STS能夠改善肺血管重塑,降低肺動脈壓力,對肺動脈高壓具有治療作用。本研究結果顯示,與常氧組相比,低氧組細胞增殖水平顯著升高,說明細胞低氧模型構建成功。與常氧組相比,STS+低氧組細胞增殖水平降低,由此表明,通過構建細胞缺氧模型,發現低氧可誘導大鼠PASMC增殖,而STS能夠抑制其增殖。
mTOR是一種絲氨酸/蘇氨酸激酶,在外源性信號(絲裂原、營養、能量)刺激下,具有調控細胞代謝、生長、增殖及生存的作用[13]。mTOR有兩種不同的復合體,即mTORC1和mTORC2。mTORC1是細胞生長增殖的主要激活物,可通過磷酸化活化p70s6激酶1(S6K1)與4E結合蛋白1(4E-BP1),進一步激活核糖體蛋白6和eIF2E,促進蛋白質合成及細胞增殖[14-15]。關于mTOR信號通路功能的相關研究主要集中在腫瘤領域[16]。也有文獻報道在特發性肺動脈高壓患者和動物肺動脈高壓模型中,mTOR具有促進PASMC增殖的作用[17-18]。另有研究顯示在低氧誘導的肺動脈高壓模型大鼠及低氧誘導的PASMC模型中,mTOR明顯活化,呈過表達,且eIF2α、c-myc也呈高表達;再通過雷帕霉素和siRNA敲除mTOR及eIF2α處理PASMC可抑制PASMC增殖及c-myc表達[6]。本研究通過細胞低氧模型檢測mTOR mRNA及蛋白表達,結果顯示低氧誘導體外培養的PASMC中mTOR活化。由此提示,mTOR在PASMC增殖中過度活化,這可能是低氧性肺動脈高壓肺血管重塑的機制之一。為進一步證實STS是否具有抑制mTOR活性的作用,本研究設計了低氧+雷帕霉素組作為陽性對照,結果顯示STS與雷帕霉干預后mTOR表達無顯著差異。因此,本研究揭示STS可通過降低mTOR表達,進而抑制PASMC增殖。
本研究結果提示,eIF2α作為mTOR的下游靶分子,可能在低氧性肺動脈高壓的PASMC增殖過程中發揮重要的調節作用。給予STS處理并以雷帕霉素作陽性對照后發現,STS能夠抑制PASMC增殖的同時,還減少了p-eIF2α的表達。當然,除了eIF2α,其他起始延長因子eIF也與mTOR信號通路相關,如mTOR結合eIF3可以促進s6k的活化[15, 19];因此,是否還有其他eIF參與還有待進一步研究。c-myc活化是血管平滑肌細胞增殖的起始之一,血管損傷時其表達異常[20]。本研究中,低氧誘導PASMC中c-myc表達上調,而STS通過抑制mTOR或eIF2α進一步下調c-myc的表達。綜上所述,丹參酮可以抑制PASMC的增殖,其機制可能是通過抑制低氧誘導的mTOR/eIF2α信號通路,進一步抑制c-myc的表達而發揮作用。
[ 1 ] Sakao S, Tatsumi K. Vascular remodeling in pulmonary arterial hypertension: multiple cancer-like pathways and possible treatment modalities[J]. Int J Cardiol, 2011, 147(1):4-12.
[ 2 ] Raghavan A, Zhou G, Zhou Q, et al. Hypoxia-induced pulmonary arterial smooth muscle cell proliferation is controlled by forkhead box M1[J]. Am J Respir Cell Mol Biol, 2012, 46(4):431-436.
[ 3 ] de Jesus Perez V, Yuan K, Alastalo TP, et al. Targeting the Wnt signaling pathways in pulmonary arterial hypertension[J]. Drug Discov Today, 2014, 19(8): 1270-1276.
[ 4 ] Christopher L, Michael R, Zhong J, et al. The interaction of endothelin-1 and TGF-β1mediates vascular cell remodeling[J]. PLoS One, 2013, 8(8): e73399.
[ 5 ] Karelina K, Liu Y, Alzate-Correa D, et al. Mitogen and stress-activated kinases 1/2 regulate ischemia-induced hippocampal progenitor cell proliferation and neurogenesis[J]. Neuroscience, 2015, 285: 292-302.
[ 6 ] Wang A, Li X, Yang Y, et al. A critical role of the mTOR/eIF2α pathway in hypoxia-induced pulmonary hypertension[J]. PLoS One, 2015, 10(6): e0130806.
[ 7 ] Zheng L, Liu M, Wei M, et al. Tanshinone ⅡA attenuates hypoxic pulmonary hypertension via modulating KV currents[J]. Respir Physiol Neurobiol, 2015, 205: 120-128.
[ 8 ] Wang J, Lu W, Wang W, et al. Promising therapeutic effects of sodium tanshinone ⅡA sulfonate towards pulmonary arterial hypertension in patients[J]. J Thorac Dis, 2013, 5(2): 169-172.
[ 9 ] Xu S, Liu P. Tanshinone Ⅱ-A: new perspectives for old remedies[J]. Expert Opin Ther Pat, 2013, 23(2): 149-153.
[10] Gao S, Liu Z, Li H, et al. Cardiovascular actions and therapeutic potential of tanshinone ⅡA[J]. Atherosclerosis, 2012, 220(1): 3-10.
[11] Jiang Q, Lu W, Yang K, et al. Sodium tanshinone ⅡA sulfonate inhibits hypoxia-induced enhancement of SOCE in pulmonary arterial smooth muscle cells via the PKG-PPAR-γ signaling axis[J]. Am J Physiol Cell Physiol, 2016, 311(1): C136-C149.
[12] Wang J, Jiang Q, Wan L, et al. Sodium tanshinone ⅡA sulfonate inhibits canonical transient receptor potential expression in pulmonary arterial smooth muscle from pulmonary hypertensive rats[J]. Am J Respir Cell Mol Biol, 2013, 48(1): 125-134.
[13] Laplante M, Sabatini DM. mTOR signaling in growth control and disease[J]. Cell, 2012, 149(2): 274-293.
[14] Liu T, Yacoub R, Taliaferrosmith LD, et al. Combinatorial effects of lapatinib and rapamycin in triple-negative breast cancer cells[J]. Mol Cancer Ther, 2011, 10(8): 1460-1469.
[15] Eckerdt F, Beauchamp E, Bell J, et al. Regulatory effects of a Mnk2-eIF4E feedback loop during mTORC1 targeting of human medulloblastoma cells[J]. Oncotarget, 2014, 5(18): 8442-8451.
[16] Evangelisti C, Ricci F, Tazzari P, et al. Targeted inhibition of mTORC1 and mTORC2 by active-site mTOR inhibitors has cytotoxic effects in t-cell acute lymphoblastic leukemia[J]. Leukemia, 2011, 25(5): 781-791.
[17] Wang W, Liu J, Ma A, et al. mTORC1 is involved in hypoxia-induced pulmonary hypertension through the activation of Notch3[J]. J Cell Physiol, 2014, 229(12): 2117-2125.
[18] Houssaini A, Abid S, Mouraret N, et al. Rapamycin reverses pulmonary artery smooth muscle cell proliferation in pulmonary hypertension[J]. Am J Respir Cell Mol Biol, 2013, 48(5): 568-577.
[19] Ahlemann M, Zeidler R, Lang S, et al. Carcinoma-associated eIF3i overexpression facilitates mTOR-dependent growth transformation[J]. Mol Carcinog, 2006, 45(12): 957-967.
[20] Sun LQ, Cairns MJ, Gerlach WL, et al. Suppression of smooth muscle cell proliferation by a c-myc RNA-cleaving deoxyribozyme[J]. J Biol Chem, 1999, 274(24): 17236-17241.
