反式肉桂醛與溫和加熱結(jié)合對(duì)復(fù)原嬰幼兒牛乳中阪崎克羅諾腸桿菌的抑殺作用
2018-04-24 12:01:28張文婷郭凱倫溫啓吾劉志遠(yuǎn)孫慧慧尹術(shù)華夏效東
食品科學(xué) 2018年7期
關(guān)鍵詞:嬰幼兒
石 超,郭 都,張文婷,郭凱倫,溫啓吾,劉志遠(yuǎn),孫慧慧,陳 珊,孫 正,郭 曉,尹術(shù)華,夏效東*
阪崎克羅諾腸桿菌(Cronobacter sakazakii)是一種周生鞭毛、無(wú)芽孢的棒狀革蘭氏陰性菌[1]。它可引起新生兒腦膜炎、菌血癥和壞死性小腸結(jié)腸炎,并可造成早產(chǎn)兒和低體質(zhì)量新生兒的死亡[2]。對(duì)于感染的嬰幼兒,死亡率高達(dá)40%~80%[3],并且,20%的感染嬰幼兒會(huì)患有嚴(yán)重的神經(jīng)后遺癥,如腦積水、四肢麻痹和智力發(fā)育遲緩等[4-5]。
研究表明,嬰幼兒乳粉是世界范圍內(nèi)嬰幼兒攝取營(yíng)養(yǎng)的主要途徑[6]。然而,嬰幼兒乳粉并不是一類(lèi)無(wú)菌的產(chǎn)品,它含有致病菌并有可能導(dǎo)致疾病[7]。嬰幼兒乳粉是阪崎克羅諾腸桿菌傳播的主要途徑[8]。嬰兒缺乏完善的免疫系統(tǒng)和穩(wěn)定的腸道菌群,因而更易被阪崎克羅諾腸桿菌感染。因此,食用前控制嬰幼兒乳粉的微生物特別是阪崎克羅諾腸桿菌污染尤為重要。
盡管針對(duì)嬰幼兒配方乳粉的優(yōu)良制造標(biāo)準(zhǔn)已發(fā)布,但是保證嬰幼兒乳粉無(wú)菌還是非常困難。為減少嬰幼兒乳粉中的微生物風(fēng)險(xiǎn),世界衛(wèi)生組織(World Health Organization,WHO)在2007年建議嬰幼兒乳粉使用70 ℃的熱水沖調(diào)并快速冷卻至適宜食用的溫度[9]。但是,使用如此高溫度的熱水沖調(diào)乳粉不被父母廣泛接受[10]。高溫會(huì)導(dǎo)致乳粉中的一些熱敏感物質(zhì)損失[11],如對(duì)熱敏感的維生素、乳鐵蛋白等;并且,高溫沖調(diào)也會(huì)增大燙傷的風(fēng)險(xiǎn)并且不方便嬰幼兒直接食用。近年來(lái),部分研究者將目光投向于在食用前的沖調(diào)過(guò)程使用溫和加熱與天然物質(zhì)結(jié)合來(lái)控制殺滅微生物[12-15]。
反式肉桂醛是肉桂樹(shù)皮提取物中的主要活性物質(zhì),已被美國(guó)食品藥品監(jiān)督管理局評(píng)價(jià)為公認(rèn)安全的食品成分(批準(zhǔn)文件序號(hào)21 CFR 182.60)。反式肉桂醛已被報(bào)道對(duì)多種食源性致病菌有抑制作用,研究表明它能夠抑殺蘋(píng)果汁及蘋(píng)果酒中的大腸桿菌O157:H7[16],抑制牛奶中的大腸桿菌并且能夠抗牛奶氧化[17],抑殺西紅柿上不同血清型的沙門(mén)氏菌[18],并且,反式肉桂醛還被報(bào)道有抗癌、抗炎、治療糖尿病、降低細(xì)菌耐藥性等多種生物活性作用[19]。
本研究采用反式肉桂醛與溫和加熱結(jié)合處理復(fù)原嬰幼兒牛乳,探討該方法對(duì)復(fù)原嬰幼兒牛乳中阪崎克羅諾腸桿菌的抑殺效果。并分別利用LIVE/DEAD?細(xì)菌活性檢測(cè)試劑盒和場(chǎng)發(fā)射掃描電子顯微鏡探究二者結(jié)合作用對(duì)阪崎克羅諾腸桿菌細(xì)胞膜完整性及細(xì)胞形態(tài)的影響,試圖闡述其可能的抑菌機(jī)理,為反式肉桂醛與溫和加熱結(jié)合應(yīng)用于復(fù)原嬰幼兒牛乳及其他食品中抑殺阪崎克羅諾腸桿菌提供理論依據(jù)。
1 材料與方法
1.1 菌株與試劑
阪崎克羅諾腸桿菌ATCC 29544購(gòu)于美國(guó)模式菌株收集中心。阪崎克羅諾腸桿菌分離菌株14-15和18-8由西北農(nóng)林科技大學(xué)食品科學(xué)與工程學(xué)院食品微生物研究團(tuán)隊(duì)分離自市售嬰幼兒乳粉。
反式肉桂醛(色譜級(jí),純度≥99%) 美國(guó)Sigma公司;胰蛋白胨大豆瓊脂(tryptone soya agar,TSA)、胰蛋白胨大豆肉湯(tryptone soya broth,TSB) 北京陸橋技術(shù)有限公司;LIVE/DEAD?BacLightTM細(xì)菌活性檢測(cè)試劑盒 美國(guó)賽默飛世爾科技公司;其他所用有機(jī)溶劑均為國(guó)產(chǎn)分析純。
1.2 儀器與設(shè)備
GHX-9050B-2細(xì)菌培養(yǎng)箱 上海福瑪實(shí)驗(yàn)設(shè)備有限公司;Smart SpecTMplus分光光度計(jì) 美國(guó)Bio-Rad公司;5804R低溫冷凍離心機(jī) 德國(guó)Eppendorf公司;Spectra Max M2多功能微孔板檢測(cè)儀 美國(guó)Molecular Devices公司;S-4800場(chǎng)發(fā)射掃描電子顯微鏡 日本Hitachi公司。
1.3 方法
1.3.1 混合菌懸液的制備
將凍存于-80 ℃的阪崎克羅諾腸桿菌ATCC 29544、14-15和18-8采用劃線法在TSA平板上活化,分別挑取單菌落接種于30 mL TSB中,將3 瓶TSB置于37 ℃、120 r/min振搖培養(yǎng)18 h,培養(yǎng)后的菌液分別離心(5 000×g、10 min、4 ℃)并去除上清液,用pH 7.2磷酸鹽緩沖溶液(phosphate buffered saline,PBS)洗滌菌體沉淀,反復(fù)2 次洗滌后,使用一定量的PBS懸浮菌體沉淀,測(cè)定并調(diào)整菌懸液的OD600nm為0.5,使菌懸液濃度約為108CFU/mL。取相同體積的3 種菌懸液混合制備為混合菌懸液(濃度約為108CFU/mL)。
1.3.2 復(fù)原嬰幼兒牛乳的制備
市售嬰幼兒乳粉購(gòu)買(mǎi)于陜西省咸陽(yáng)市楊凌區(qū)好又多超市。按照商品標(biāo)簽沖調(diào)復(fù)原嬰幼兒乳粉,將25.5 g乳粉溶解于180 mL無(wú)菌水中,搖勻后置于63 ℃水浴30 min進(jìn)行巴氏殺菌[20],冷卻至室溫后備用。
1.3.3 阪崎克羅諾腸桿菌的抑菌作用
將復(fù)原嬰幼兒牛乳加入試管(10 mL),向各組接種100 μL三種阪崎克羅諾腸桿菌的混合菌懸液,使復(fù)原嬰幼兒牛乳中阪崎克羅諾腸桿菌的濃度約為6.5~7.0(lg(CFU/mL)),同時(shí)向樣品中添加反式肉桂醛溶液(溶劑為含0.5% DMSO的無(wú)菌水),使反式肉桂醛溶液的質(zhì)量分?jǐn)?shù)分別為0.4%、0.3%、0.2%和0.1%(下同),將不添加反式肉桂醛的樣品(含有0.5% DMSO)作為對(duì)照組。實(shí)驗(yàn)室前期結(jié)果表明1% DMSO在復(fù)原嬰幼兒牛乳中不會(huì)影響阪崎克羅諾腸桿菌的活性。將接種后的樣品置于4 個(gè)不同的溫度(25、45、50、55 ℃)保溫殺菌。將25 ℃和45 ℃的樣品分別保溫殺菌0、20、40、60、90、120 min,50 ℃和55 ℃的樣品分別保溫殺菌0、10、20、40、60、90 min后,將樣品直接或用PBS 10 倍稀釋后,涂布到TSA培養(yǎng)基上37 ℃恒溫培養(yǎng)24 h,記錄細(xì)菌總數(shù),實(shí)驗(yàn)每組設(shè)置3 個(gè)平行。
1.3.4 阪崎克羅諾腸桿菌細(xì)胞膜完整性的測(cè)定
溫和加熱與反式肉桂醛結(jié)合作用對(duì)阪崎克羅諾腸桿菌細(xì)胞膜完整性影響的測(cè)定,參考Shi Chao等[21]所采用的LIVE/DEAD?細(xì)菌活性試劑盒方法。試劑盒中含有SYTO 9及碘化丙啶(propidium iodide,PI)兩種核酸染料。SYTO 9染料能夠穿透完整及受損的細(xì)胞膜,將所有細(xì)菌染色為綠色。相反,PI僅能穿透受損細(xì)胞膜,將細(xì)胞膜受損的細(xì)胞染色為紅色,并降低SYTO 9的綠色熒光。因此,當(dāng)在細(xì)菌懸液中添加SYTO 9與PI兩種染料時(shí),細(xì)胞膜完整的菌體呈現(xiàn)綠色,細(xì)胞膜不完整的菌體呈現(xiàn)紅色。為探究結(jié)合作用對(duì)阪崎克羅諾腸桿菌的抑制機(jī)理,我們選擇ATCC 29544菌株進(jìn)行后續(xù)的研究。將37 ℃振搖(120 r/min)培養(yǎng)18 h的阪崎克羅諾腸桿菌離心(10 000×g、15 min、4 ℃)棄去培養(yǎng)液,將菌體沉淀溶解于2 mL的0.85% NaCl溶液中。分別取1 mL菌懸液加入20 mL 0.85% NaCl溶液和20 mL 70%異丙醇溶液并置于室溫孵育1 h(每15 min輕柔振搖1次),從而得到活細(xì)胞和死細(xì)胞懸液。隨后將兩管菌懸液離心(10 000×g,10 min),使用0.85% NaCl溶液將活細(xì)胞和死細(xì)胞沉淀重新懸浮并制備為活細(xì)胞和死細(xì)胞混合溶液,使活細(xì)胞的比例為0、10%、50%、90%和100%。
在無(wú)菌的離心管中加入活細(xì)胞樣品,按照處理?xiàng)l件不同將菌懸液分為4 組:對(duì)照組(25 ℃處理)、單純反式肉桂醛組(0.2%反式肉桂醛+25 ℃處理)、單純溫和加熱組(50 ℃處理)和結(jié)合作用實(shí)驗(yàn)組(0.2%反式肉桂醛+50 ℃處理),每組3 個(gè)平行。將樣品處理后離心(11 000×g、1 min)并使用0.85% NaCl重懸浮。將100 μL標(biāo)準(zhǔn)樣品及實(shí)驗(yàn)樣品加入黑色96 孔酶標(biāo)板中,將SYTO 9和PI混合染料加入每孔并輕柔混合。將樣品置于25 ℃避光孵育15 min,隨后使用多功能酶標(biāo)儀檢測(cè)熒光強(qiáng)度。綠色熒光染料SYTO 9的激發(fā)、發(fā)射波長(zhǎng)分別為485、542 nm,紅色熒光染料PI的激發(fā)、發(fā)射波長(zhǎng)分別為485、610 nm。通過(guò)檢測(cè)不同活細(xì)菌比例標(biāo)準(zhǔn)樣品(0、10%、50%、90%和100%)的綠色熒光強(qiáng)度,構(gòu)建綠色熒光強(qiáng)度與細(xì)胞膜完整細(xì)菌(活細(xì)菌)百分比之間的線性關(guān)系。檢測(cè)實(shí)驗(yàn)組的綠色熒光強(qiáng)度,根據(jù)本研究測(cè)定的綠色熒光強(qiáng)度與細(xì)胞膜完整細(xì)菌百分比之間的擬合方程,計(jì)算出細(xì)胞膜完整細(xì)菌百分比。
1.3.5 阪崎克羅諾腸桿菌細(xì)胞形態(tài)的觀察
反式肉桂醛與溫和加熱結(jié)合對(duì)阪崎克羅諾腸桿菌細(xì)胞形態(tài)的影響參照Li Guanghui等[22]利用場(chǎng)發(fā)射掃描電子顯微鏡的方法。將37 ℃、120 r/min振搖培養(yǎng)18 h的阪崎克羅諾腸桿菌離心(5 000×g、15 min、4 ℃)并棄去TSB培養(yǎng)液,使用TSB將菌體沉淀重新懸浮。將菌懸液分為4 組,同1.3.4節(jié),將樣品分別培養(yǎng)1 h或2 h。隨后將菌體離心并使用PBS清洗兩次,使用含有0.25%戊二醛的PBS 4 ℃固定細(xì)胞。12 h后將菌體離心(5 000×g、10 min、4 ℃)并使用PBS及無(wú)菌水分別清洗菌體1次,隨后使用含有1%鋨酸的PBS于4 ℃固定細(xì)胞5 h后,將菌體離心(5 000×g、10 min、4 ℃),并使用不同體積分?jǐn)?shù)的無(wú)水乙醇(30%、50%、70%、80%、90%和100%)梯度洗脫細(xì)胞,每次10 min。將菌體滴加至潔凈的玻片表面進(jìn)行干燥后噴金處理,使用場(chǎng)發(fā)射掃描電子顯微鏡觀測(cè)細(xì)胞形態(tài)。
1.4 數(shù)據(jù)統(tǒng)計(jì)分析
所有實(shí)驗(yàn)重復(fù)3 次,結(jié)果以 ±s表示。使用SPSS 19.0軟件,對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析。采用ANOVA對(duì)結(jié)果間的顯著性進(jìn)行比較,不同處理間P<0.05認(rèn)為差異顯著,P<0.01則認(rèn)為差異極顯著。
2 結(jié)果與分析
2.1 阪崎克羅諾腸桿菌的抑菌效果
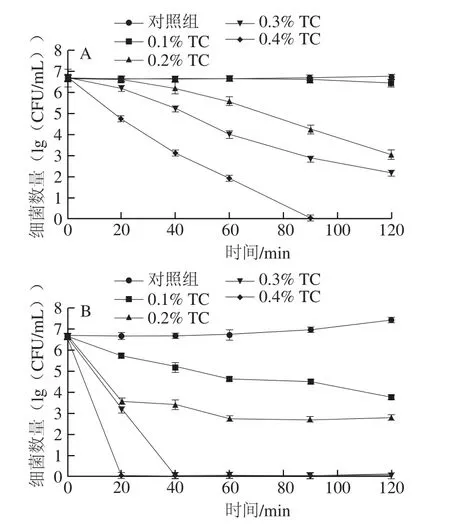
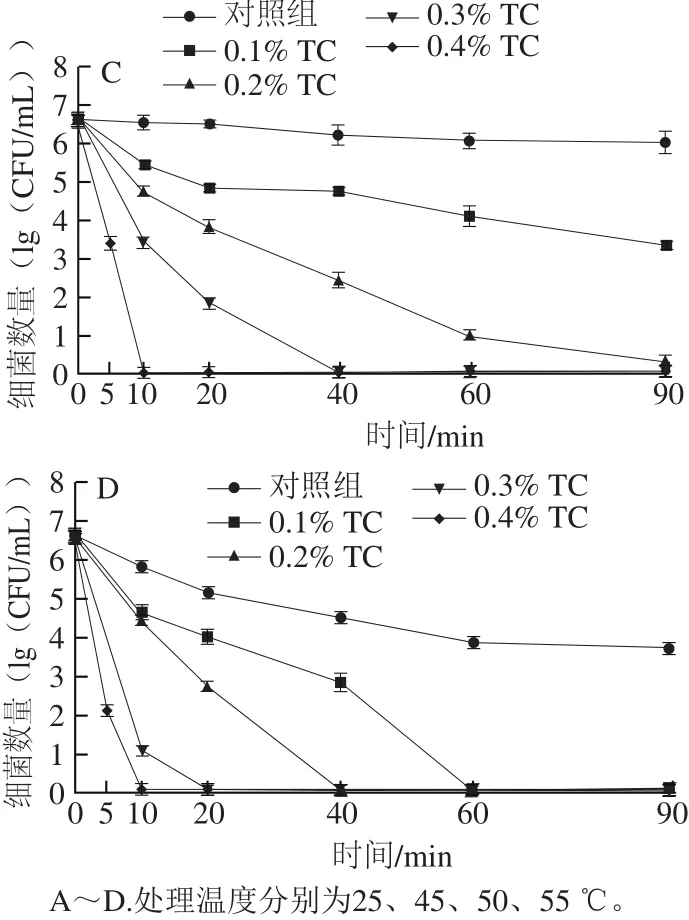
圖1 反式肉桂醛結(jié)合溫和加熱對(duì)復(fù)原嬰幼兒牛乳中阪崎克羅諾腸桿菌數(shù)量的影響Fig. 1 Effect of combined treatment with trans-cinnamaldehyde and mild heating on C. sakazakii populations in reconstituted infant formula
復(fù)原嬰幼兒牛乳樣品中阪崎克羅諾腸桿菌的起始濃度約為6.6(lg(CFU/mL))。由圖1A可知,對(duì)照組在120 min內(nèi)細(xì)菌總數(shù)與起始值沒(méi)有顯著性差異(P>0.05)。經(jīng)0.1%反式肉桂醛處理120 min后,阪崎克羅諾腸桿菌總數(shù)下降約0.3(lg(CFU/mL))。經(jīng)0.2%、0.3%和0.4%反式肉桂醛處理60 min后,細(xì)菌總數(shù)分別下降約1.1、2.7、4.7(lg(CFU/mL))。處理90 min后,0.4%反式肉桂醛使阪崎克羅諾腸桿菌細(xì)菌總數(shù)下降至檢出限以下。
由圖1B可知,對(duì)照組在120 min細(xì)菌總數(shù)增加約0.8(lg(CFU/mL)),這說(shuō)明阪崎克羅諾腸桿菌對(duì)45 ℃有一定的耐受能力。經(jīng)0.1%、0.2%和0.3%反式肉桂醛處理20 min后,阪崎克羅諾腸桿菌細(xì)菌總數(shù)下降約0.9、3.1、3.5(lg(CFU/mL)),經(jīng)0.4%反式肉桂醛處理20 min及0.3%反式肉桂醛處理40 min后,細(xì)菌總數(shù)下降至檢出限以下。
由圖1C可知,對(duì)照組在作用60 min后,阪崎克羅諾腸桿菌總數(shù)下降約0.5(lg(CFU/mL)),但作用90 min與60 min沒(méi)有顯著性差異(P>0.05)。經(jīng)0.1%、0.2%和0.3%反式肉桂醛結(jié)合50 ℃處理20 min后,阪崎克羅諾腸桿菌總數(shù)分別下降約1.9、2.7、4.8(lg(CFU/mL))。經(jīng)0.1%和0.2%反式肉桂醛結(jié)合50 ℃處理90 min后,細(xì)菌總數(shù)分別下降約3.4、6.2(lg(CFU/mL))。0.3%反式肉桂醛在作用40 min及0.4%的反式肉桂醛作用在作用10 min時(shí)將阪崎克羅諾腸桿菌的總數(shù)降低至檢出限以下。
由圖1D可知,反式肉桂醛和55 ℃溫和加熱結(jié)合與45 ℃和50 ℃相比對(duì)阪崎克羅諾腸桿菌有較強(qiáng)的抑殺效果。對(duì)照組在作用90 min后,阪崎克羅諾腸桿菌總數(shù)下降約3.0(lg(CFU/mL))。經(jīng)0.1%反式肉桂醛處理40 min后,阪崎克羅諾腸桿菌總數(shù)下降約3.8(lg(CFU/mL)),處理60 min后,細(xì)菌總數(shù)降低至檢出限以下。經(jīng)0.2%反式肉桂醛處理20 min后,阪崎克羅諾腸桿菌總數(shù)下降約3.8(lg(CFU/mL)),處理40 min后,細(xì)菌總數(shù)降低至檢出限以下。經(jīng)0.3%反式肉桂醛處理10 min后,阪崎克羅諾腸桿菌總數(shù)下降約5.7(lg(CFU/mL)),處理20 min后,細(xì)菌總數(shù)降低至檢出限以下。經(jīng)0.4%反式肉桂醛處理5 min后,阪崎克羅諾腸桿菌總數(shù)下降約4.6(lg(CFU/mL)),處理10 min后,細(xì)菌總數(shù)降低至檢出限以下。
2.2 對(duì)阪崎克羅諾腸桿菌細(xì)胞膜完整性的影響
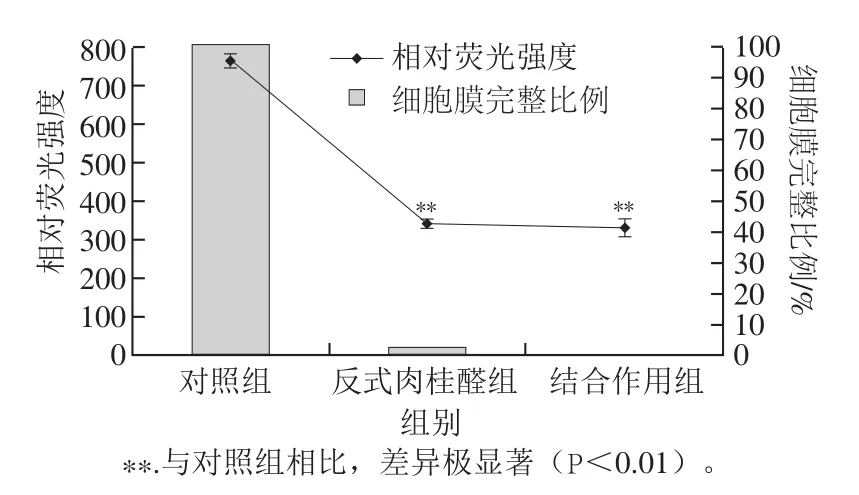
圖2 不同處理組對(duì)阪崎克羅諾腸桿菌ATCC 29544細(xì)胞膜熒光強(qiáng)度的影響Fig. 2 Fluorescence of C. sakazakii ATCC 29544 subjected to various treatments
實(shí)驗(yàn)首先構(gòu)建了綠色熒光與細(xì)胞膜完整細(xì)菌百分比的線性關(guān)系,得到綠色熒光強(qiáng)度與細(xì)胞膜完整細(xì)菌百分比的線性擬合方程:y=431.3x+329.5(R2=0.988)。由圖2可知,未經(jīng)反式肉桂醛和溫和加熱處理的對(duì)照組,細(xì)胞膜完整的細(xì)菌百分比為100%,經(jīng)0.2%反式肉桂醛處理20 min后,細(xì)胞膜完整的細(xì)胞比例為2.73%,與對(duì)照組有極顯著差異(P<0.01)。經(jīng)0.2%反式肉桂醛與50 ℃結(jié)合作用處理20 min,細(xì)胞膜完整的細(xì)胞比例僅為0.65%,與對(duì)照組有極顯著差異(P<0.01)。
2.3 對(duì)阪崎克羅諾腸桿菌細(xì)胞形態(tài)的影響
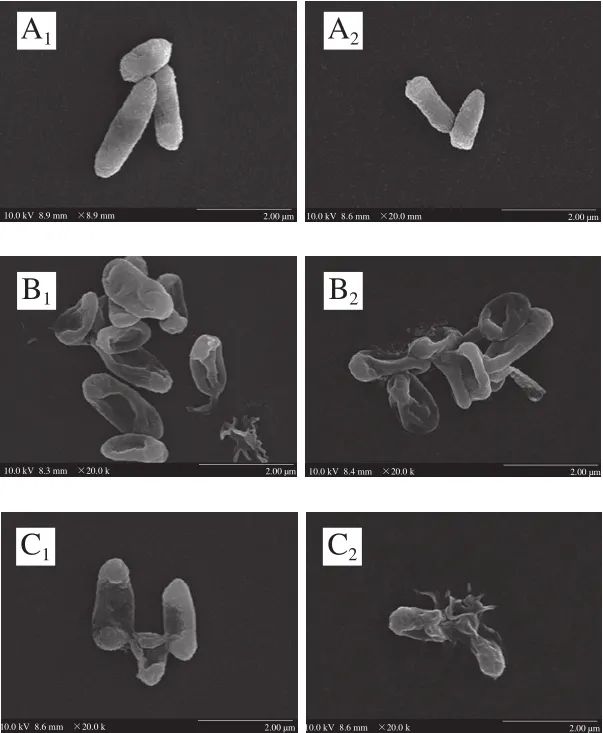
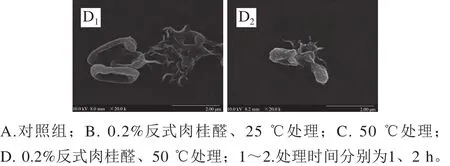
圖3 場(chǎng)發(fā)射掃描電子顯微鏡下阪崎克羅諾腸桿菌的形態(tài)Fig. 3 Scanning electron micrographs of C. sakazakii
場(chǎng)發(fā)射掃描電子顯微鏡觀測(cè)發(fā)現(xiàn),未經(jīng)溫和加熱和反式肉桂醛處理的阪崎克羅諾腸桿菌呈棒狀,菌體飽滿(mǎn),表面光滑(圖3A、E)。經(jīng)0.2%反式肉桂醛作用后,阪崎克羅諾腸桿菌菌體從中部干癟,細(xì)胞扁平呈橢圓狀,且部分細(xì)胞破碎,出現(xiàn)碎片(圖3B、F)。經(jīng)50 ℃處理的阪崎克羅諾腸桿菌菌體出現(xiàn)皺縮,并有部分細(xì)菌破裂(圖3C、G)。反式肉桂醛與溫和加熱結(jié)合作用后阪崎克羅諾腸桿菌的形態(tài)如圖3D、H所示,結(jié)合作用后1 h,部分細(xì)胞干癟,部分細(xì)胞破裂。結(jié)合作用后2 h,大部分菌體破裂且瓦解為碎片。結(jié)合作用比反式肉桂醛單獨(dú)作用和溫和加熱單獨(dú)作用造成的菌體皺縮干癟程度更大,破碎菌體比例更大。
3 討 論
研究表明,與腸桿菌科的其他屬種相比,阪崎克羅諾腸桿菌有著較強(qiáng)的耐干燥的能力[23],能夠在干燥的乳粉中存活2 年[24]。Breeuwer等[25]發(fā)現(xiàn)阪崎克羅諾腸桿菌能夠通過(guò)維持細(xì)胞膜蛋白和磷脂的穩(wěn)定從而保護(hù)自身應(yīng)對(duì)高滲透壓環(huán)境。這說(shuō)明阪崎克羅諾腸桿菌能夠在乳粉的貨架期中長(zhǎng)期存活,為抑殺阪崎克羅諾腸桿菌從而降低其引起的感染,食用前的沖調(diào)乳粉過(guò)程尤為重要。盡管WHO已推薦消費(fèi)者使用70 ℃的熱水沖調(diào)乳粉迅速降低至適宜的溫度,但由于該方法存在操作繁瑣,導(dǎo)致乳液感官品質(zhì)及營(yíng)養(yǎng)改變、易造成燙傷等多種缺點(diǎn)而不被消費(fèi)者廣泛接受。溫和加熱結(jié)合天然物質(zhì)對(duì)食品中食源性致病菌的抑殺作用成為近年來(lái)的研究熱點(diǎn)。Espina等[26]研究結(jié)果表明200 μL/L柑橘精油與54 ℃結(jié)合作用與單獨(dú)加熱相比能夠顯著縮短作用時(shí)間,提高蘋(píng)果汁中大腸桿菌O157:H7的抑殺效率。與美國(guó)食品藥品管理局推薦的71.11~82.22 ℃處理果汁的方法相比,能夠減少能耗并維持果汁良好的感官性狀。Gabriel等[27]研究結(jié)果表明,香草醛及甘草根提取物與55 ℃結(jié)合能夠有效降低椰汁中大腸桿菌O157:H7的數(shù)量。Choi等[28]研究結(jié)果表明20 mmol/L辛酸和30 mmol/L香草醛結(jié)合45 ℃能在5 min將復(fù)原嬰幼兒牛乳中的鼠傷寒沙門(mén)氏菌降低至檢出限以下(起始菌濃度約為7.0(lg(CFU/mL))。
在實(shí)驗(yàn)中,0.3%反式肉桂醛處理的復(fù)原嬰幼兒牛乳在50 ℃和55 ℃能夠顯著降低阪崎克羅諾腸桿菌的數(shù)量,在作用20 min使阪崎克羅諾腸桿菌總數(shù)至檢出限以下,然而在相同的作用時(shí)間內(nèi),反式肉桂醛在50 ℃和45 ℃對(duì)阪崎克羅諾腸桿菌的抑殺作用較弱(分別下降4.8、3.5(lg(CFU/mL)))。類(lèi)似的,Jang等[11]研究表明與50 ℃和55 ℃相比,辛酸在45 ℃下對(duì)阪崎腸桿菌抑制作用較弱。Char等[29]發(fā)現(xiàn)當(dāng)溫和加熱溫度升高時(shí),香草醛在橙汁中的殺菌作用更強(qiáng)。反式肉桂醛隨溫度升高抑殺作用增強(qiáng)可能歸因于升高的溫度使反式肉桂醛在復(fù)原嬰幼兒牛乳中的溶解度升高。同時(shí),猜測(cè)溫和加熱作用能夠造成細(xì)菌細(xì)胞膜受損從而增強(qiáng)反式肉桂醛的抑殺作用。
為闡明反式肉桂醛與溫和加熱結(jié)合對(duì)阪崎克羅諾腸桿菌細(xì)胞形態(tài)的改變及細(xì)胞膜的損傷,利用場(chǎng)發(fā)射掃描電子顯微鏡對(duì)未處理、經(jīng)過(guò)反式肉桂醛處理、經(jīng)過(guò)溫和加熱處理和經(jīng)過(guò)反式肉桂醛與溫和加熱結(jié)合處理的阪崎克羅諾腸桿菌進(jìn)行了觀測(cè)。結(jié)果表明單獨(dú)溫和加熱作用使細(xì)胞皺縮,延長(zhǎng)作用時(shí)間,菌體出現(xiàn)了破碎,呈現(xiàn)作用時(shí)間依賴(lài)性。這是由于溫和加熱影響了細(xì)胞膜蛋白的空間結(jié)構(gòu)松散、伸展,造成細(xì)胞膜的穿孔和裂隙,并且使膜蛋白更容易被蛋白酶水解。單獨(dú)反式肉桂醛作用使阪崎克羅諾腸桿菌菌體皺縮及破裂。Pasqua等[30]通過(guò)測(cè)定處理后的細(xì)菌細(xì)胞膜不飽和脂肪酸含量,得出肉桂醛的主要作用靶點(diǎn)在于細(xì)胞膜,肉桂醛能夠增大菌體細(xì)胞膜的面積,改變細(xì)胞膜的結(jié)構(gòu),并能夠使金黃色葡萄球菌細(xì)胞膜破裂,同時(shí),肉桂醛也能夠穿透細(xì)胞膜進(jìn)入細(xì)胞,造成菌體死亡。Helander等[31]研究推測(cè)反式肉桂醛是通過(guò)OM轉(zhuǎn)運(yùn)孔蛋白進(jìn)入細(xì)胞的。本研究中,溫和加熱和反式肉桂醛結(jié)合作用使菌體破裂數(shù)量增多,瓦解破碎程度增大,這說(shuō)明結(jié)合作用對(duì)細(xì)胞膜的破壞程度大于反式肉桂醛和溫和加熱單獨(dú)作用。
本實(shí)驗(yàn)探究了反式肉桂醛與溫和加熱結(jié)合對(duì)復(fù)原嬰幼兒牛乳中阪崎克羅諾腸桿菌的抑殺效果,結(jié)果表明這種結(jié)合作用對(duì)阪崎克羅諾腸桿菌有良好的抑殺效果。反式肉桂醛的抑殺活性隨溫度的升高而增大,0.4%反式肉桂醛與55 ℃結(jié)合在10 min內(nèi)使阪崎克羅諾腸桿菌全部失活。通過(guò)LIVE/DEAD?細(xì)菌活性檢測(cè)試劑盒檢測(cè)結(jié)合作用對(duì)細(xì)胞膜完整性的影響,結(jié)果表明與對(duì)照組相比單獨(dú)反式肉桂醛作用與結(jié)合作用使細(xì)胞膜完整性極顯著降低(P<0.01)。利用場(chǎng)發(fā)射掃描電子顯微鏡探究對(duì)細(xì)胞形態(tài)的影響,結(jié)果表明反式肉桂醛與溫和加熱結(jié)合增大了細(xì)胞形態(tài)的皺縮干癟程度,并且使細(xì)胞破裂比例增大。以上結(jié)果表明反式肉桂醛有潛力作為抑菌劑在嬰幼兒乳粉中應(yīng)用,反式肉桂醛在乳粉復(fù)原沖調(diào)時(shí)與溫水或溫和加熱結(jié)合,能夠降低通過(guò)污染的嬰幼兒乳粉造成阪崎克羅諾腸桿菌感染的風(fēng)險(xiǎn),這種結(jié)合的方法有潛力替代常規(guī)使用高溫度水沖調(diào)乳粉的方式。然而,現(xiàn)行的GB 2760—2014《食品安全國(guó)家標(biāo)準(zhǔn) 食品添加劑使用標(biāo)準(zhǔn)》中,僅規(guī)定了肉桂醛作為防腐劑用途在表面處理的鮮水果的殘留量,對(duì)復(fù)原嬰幼兒牛乳中的限量問(wèn)題沒(méi)有規(guī)定。因此,反式肉桂醛作為食品添加劑在復(fù)原嬰幼兒乳粉中的添加量及與溫和加熱結(jié)合對(duì)復(fù)原嬰兒牛乳的感官品質(zhì)的影響,需要在推薦使用前深入探究。
參考文獻(xiàn):
[1] KIM T J, SILVA J L, WENG W L, et al. Inactivation of Enterobacter sakazakii by water-soluble muscadine seed extracts[J]. International Journal of Food Microbiology, 2009, 129(3): 295-299. DOI:10.1016/j.ijfoodmicro.2008.12.014.
[2] BAR-OZ B, PREMINGER A, PELEG O, et al. Enterobacter sakazakii infection in the newborn[J]. Acta Paediatrica, 2001, 90(3): 356-358.DOI:10.1080/080352501300067857.
[3] BOWEN A B, BRADEN C R. Invasive Enterobacter sakazakii disease in infants[J]. Emerging Infectious Diseases, 2006, 12(8): 1185-1189.
[4] FORSYTHE S J. Enterobacter sakazakii and other bacteria in powdered infant milk formula[J]. Maternal and Child Nutrition, 2005, 1(1): 44-50.DOI:10.1111/j.1740-8709.2004.00008.x.
[5] LAI K K. Enterobacter sakazakii infections among neonates, infants, children, and adults: case reports and a review of the literature[J]. Medicine, 2001, 80(2): 113-122. DOI:10.1097/00005792-200103000-00004.
[6] DRUDY D, MULLANE N R, QUINN T, et al. Enterobacter sakazakii:an emerging pathogen in powdered infant formula[J]. Clinical Infectious Diseases, 2006, 42(7): 996-1002. DOI:10.1086/501019.
[7] TORLAK E, SERT D. Inactivation of Cronobacter by gaseous ozone in milk powders with different fat contents[J]. International Dairy Journal,2013, 32(2): 121-125. DOI:10.1016/j.idairyj.2013.05.013.
[8] ZHOU X F, FU S Z, GAO J X, et al. Enterobacter sakazakii: an emerging foodborne pathogenic bacterium[J]. Annals of Microbiology, 2012,62(1): 1-5. DOI:10.1007/s13213-011-0274-x.
[9] World Health Organization. Safe preparation, storage and handling of powdered infant formula: guidelines[A/OL]. [2017-12-10]. http://www.who.int/iris/handle/10665/43659.
[10] CARLETTI C, CATTANEO A. Home preparation of powdered infant formula: is it safe?[J]. Acta Paediatrica, 2008, 97(8): 1131-1132.DOI:10.1111/j.1651-2227.2008.00846.x.
[11] JANG H I, RHEE M S. Inhibitory effect of caprylic acid and mild heat on Cronobacter spp. (Enterobacter sakazakii) in reconstituted infant formula and determination of injury by flow cytometry[J]. International Journal of Food Microbiology, 2009, 133(1/2): 113-120. DOI:10.1016/j.ijfoodmicro.2009.05.009.
[12] AIT-OUAZZOU A, ESPINA L, GARCIA-GONZALO D, et al.Synergistic combination of physical treatments and carvacrol for Escherichia coli O157:H7 inactivation in apple, mango, orange, and tomato juices[J]. Food Control, 2013, 32(1): 159-167. DOI:10.1016/j.foodcont.2012.11.036.
[13] GAYAN E, SERRANO M J, MONFORT S, et al. Combining ultraviolet light and mild temperatures for the inactivation of Escherichia coli in orange juice[J]. Journal of Food Engineering, 2012,113(4): 598-605. DOI:10.1016/j.jfoodeng.2012.07.018.
[14] PAN X J, MAK L M, NAKANO H. Efficacy of essential oils on inactivation of Escherichia coli O157:H7 in vegetable juice[J].Food Science and Technology Research, 2014, 20(5): 1043-1049.DOI:10.3136/Fstr.20.1043.
[15] SILVA F V M, TAN E K, FARID M. Bacterial spore inactivation at 45-65 ℃ using high pressure processing: study of Alicyclobacillus acidoterrestris in orange juice[J]. Food Microbiology, 2012, 32(1): 206-211. DOI:10.1016/J.Fm.2012.04.019.
[16] BASKARAN S A, AMALARADJOU M A R, HOAGLAND T, et al.Inactivation of Escherichia coli O157:H7 in apple juice and apple cider by trans-cinnamaldehyde[J]. International Journal of Food Microbiology,2010, 141(1/2): 126-129. DOI:10.1016/j.ijfoodmicro.2010.04.002.
[17] CHUN J Y, KIM K B, SHIN J B, et al. Effect of trans-cinnamaldehyde and high pressure treatment on physico-chemical and microbial properties of milk during storage periods[J]. Korean Journal for Food Science of Animal Resources, 2013, 33(1): 16-23. DOI:10.5851/kosfa.2013.33.1.16.
[18] MATTSON T E, JOHNY A K, AMALARADJOU M A R, et al.Inactivation of Salmonella spp. on tomatoes by plant molecules[J].International Journal of Food Microbiology, 2011, 144(3): 464-468.DOI:10.1016/j.ijfoodmicro.2010.10.035.
[19] ZHANG L Q, ZHANG Z G, FU Y, et al. Trans-cinnamaldehyde improves memory impairment by blocking microglial activation through the destabilization of iNOS mRNA in mice challenged with lipopolysaccharide[J]. Neuropharmacology, 2016, 110: 503-518.DOI:10.1016/j.neuropharm.2016.08.013.
[20] AMALARADJOU M A R, HOAGLAND T A,VENKITANARAYANAN K. Inactivation of Enterobacter sakazakii in reconstituted infant formula by trans-cinnamaldehyde[J]. International Journal of Food Microbiology, 2009, 129(2): 146-149. DOI:10.1016/j.ijfoodmicro.2008.11.016.
[21] SHI Chao, SONG Kaikuo, ZHANG Xiaorong, et al. Antimicrobial activity and possible mechanism of action of citral against Cronobacter sakazakii[J]. PLoS ONE, 2016, 11(7): e0159006. DOI:10.1371/journal.pone.0159006.
[22] LI Guanghui, XU Yunfeng, WANG Xin, et al. Tannin-rich fraction from pomegranate rind damages membrane of Listeria monocytogenes[J].Foodborne Pathogens and Disease, 2014, 11(4): 313-319. DOI:10.1089/fpd.2013.1675.
[23] CAUBILLA-BARRON J, HURRELL E, TOWNSEND S, et al.Genotypic and phenotypic analysis of Enterobacter sakazakii strains from an outbreak resulting in fatalities in a neonatal intensive care unit in France[J]. Journal of Clinical Microbiology, 2007, 45(12): 3979-3985.DOI:10.1128/Jcm.01075-07.
[24] BARRON J C, FORSYTHE S J. Dry stress and survival time of Enterobacter sakazakii and other Enterobacteriaceae in dehydrated powdered infant formula[J]. Journal of Food Protection, 2007, 70(9):2111-2117.
[25] BREEUWER P, LARDEAU A, PETERZ M, et al. Desiccation and heat tolerance of Enterobacter sakazakii[J]. Journal of Applied Microbiology,2003, 95(5): 967-973. DOI:10.1046/j.1365-2672.2003.02067.x.
[26] ESPINA L, SOMOLINOS M, OUAZZOU A A, et al. Inactivation of Escherichia coli O157:H7 in fruit juices by combined treatments of citrus fruit essential oils and heat[J]. International Journal of Food Microbiology,2012, 159(1): 9-16. DOI:10.1016/j.ijfoodmicro.2012.07.020.
[27] GABRIEL A A, PINEDA J K F. Influences of vanillin and licorice root extract supplementations on the decimal reduction times of Escherichia coli O157:H7 in mildly heated young coconut liquid endosperm[J]. Food Control, 2014, 38: 136-141. DOI:10.1016/j.foodcont.2013.10.011.
[28] CHOI M J, KIM S A, LEE N Y, et al. New decontamination method based on caprylic acid in combination with citric acid or vanillin for eliminating Cronobacter sakazakii and Salmonella enterica serovar Typhimurium in reconstituted infant formula[J]. International Journal of Food Microbiology, 2013, 166(3): 499-507. DOI:10.1016/j.ijfoodmicro.2013.08.016.
[29] CHAR C D, GUERRERO S N, ALZAMORA S M. Mild thermal process combined with vanillin plus citral to help shorten the inactivation time for Listeria innocua in orange juice[J]. Food and Bioprocess Technology,2010, 3(5): 752-761. DOI:10.1007/s11947-008-0155-x.
[30] PASQUA R D, BETTS G, HOSKINS N, et al. Membrane toxicity of antimicrobial compounds from essential oils[J]. Journal of Agricultural and Food Chemistry, 2007, 55(12): 4863-4870. DOI:10.1021/Jf0636465.
[31] HELANDER I M, ALAKOMI H L, LATVA-KALA K, et al.Characterization of the action of selected essential oil components on Gram-negative bacteria[J]. Journal of Agricultural and Food Chemistry,1998, 46(9): 3590-3595. DOI:10.1021/Jf980154m.
猜你喜歡
幸福家庭(2023年8期)2023-08-17 10:06:50
紡織標(biāo)準(zhǔn)與質(zhì)量(2022年1期)2022-07-12 06:01:12
家庭醫(yī)學(xué)(下半月)(2020年2期)2020-05-11 02:07:32
華人時(shí)刊(2019年23期)2019-05-21 03:31:38
健康管理(2017年4期)2017-05-20 08:17:06
媽媽寶寶(2017年4期)2017-02-25 07:00:48
媽媽寶寶(2017年2期)2017-02-21 01:21:04
中國(guó)當(dāng)代醫(yī)藥(2015年20期)2015-03-01 02:04:41
中國(guó)醫(yī)療美容(2015年5期)2015-02-03 03:02:00
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年17期)2014-03-01 04:29:21
