連續水旱輪作對水稻冷浸田土壤細菌群落結構的影響*
2018-04-13 03:50:10李清華何春梅
土壤學報 2018年2期
方 宇 王 飛 李清華 林 誠 何春梅
(福建省農業科學院土壤肥料研究所,福州 350013)
冷浸田是指山丘谷地受冷水、冷泉浸漬或湖區灘地受地下水浸漬而形成的一種強育性低產水稻田[1]。據統計,我國有冷浸田約400萬hm2,已成為我國糧食增產的主要障礙之一[2]。冷浸田由于長期受冷水浸泡,具有水冷土溫低、還原性強和產量低等特征[3]。然而冷浸田不同于其他的中低產田,其有機質含量豐富,潛在肥力較高,增產潛力巨大。若對冷浸田進行針對性的改良措施,提高農田的生產能力,對保障糧食生產與促進農民增收具有重要的現實意義。
水旱輪作是冷浸田土壤獲得水稻高產的一項有效措施[4]。在水旱輪作條件下,由于季節性的干濕交替使得土壤中的物理和化學性質發生了一系列的變化,這勢必會影響土壤中的微生物。植物根際的土壤微生物群落對不同作物的根系分泌物反應不同[5]。而根際土壤的微生物群落又會對非根際土壤的微生物群系產生影響[6]。微生物群落多樣性與土壤生態系統的結構和功能密切相關,在維持土壤肥力和生態平衡中發揮著重要作用[7],對外界環境較敏感,是土壤變化的重要指示因子之一[8]。各學者關于作物輪作對土壤微生物多樣性影響的研究分歧較大[9-11]。上述輪作研究多集中于障礙特征不明顯的土壤類型,而針對以潛育化為主要特征的冷浸田土壤的報道尚少。水旱輪作下冷浸田土壤的研究主要集中在輪作對冷浸田土壤的理化性狀和微生物數量[12]、活性碳氮磷[13]以及土壤亞鐵的影響[14]。然而,冷浸田土壤細菌群落結構對水旱輪作的響應規律尚未有系統研究。
為了尋求適合冷浸田的輪作體系,提高冷浸田的生產能力,本研究通過冷浸田水旱輪作改制試驗,采用高通量測序技術探討冷浸田由單季稻改為水旱輪作制后對土壤細菌群落結構的影響,旨在為冷浸田治理利用提供理論依據。
1 材料與方法
1.1 研究區概況
試驗設于農業部福建耕地保育科學野外觀測實驗站(閩侯縣白沙鎮溪頭村,119°04'10″E、26°13'31″N)。所處地形為山前傾斜平原交接洼地,土壤類型為淺腳爛泥田,屬潛育化程度較低的冷浸田。成土母質為低丘坡積物。試驗前為單季稻-冬閑。供試土壤基本性狀:有機質34.0 g kg-1、全氮2.01 g kg-1、全磷 0.32 g kg-1、全鉀 13.0 g kg-1、速效氮 173.4 mg kg-1、有效磷 5.5 mg kg-1、速效鉀49.5 mg kg-1,pH 5.3。
1.2 試驗設計
輪作試驗共設5個處理:(1)單季稻-冬閑(CK);(2)油菜-水稻(R-R);(3)春玉米-水稻(C-R);(4)紫云英-水稻(M-R);(5)蠶豆-水稻(B-R)。每處理設3次重復,采用隨機區組排列。每小區面積10 m2(長4 m,寬2.5 m)。各作物品種、種植時間、種植密度、播種量及施肥方式詳見王飛等[12]所述。
試驗自2011年10月上旬單季稻收割后開始,連續種植4茬旱作與4茬單季稻(其中CK處理分別于2012~2015年連續4年種植單季稻,冬季空閑),2015年10月上旬結束。由于冷浸田冬季地下水位高,旱作試驗前,小區內四周開環溝(30 cm×30 cm),以適當降低地下水位。旱作期結合起畦栽培,畦高20 cm。紫云英于盛花期(4月上旬)翻壓,各旱作作物收獲后秸稈全部粉碎回田并翻壓入土,田間保持濕潤狀態,自然腐熟。水稻生育期各處理以淺水灌溉為主。
2015年10月份水稻收獲期采用五點法采集各小區耕層土樣(0~20 cm)約1 kg,部分土樣風干,測定土壤理化性狀;部分土樣-80℃保存,用于高通量測序分析。
1.3 測定方法
土壤理化性質按常規分析方法[15]測定:土壤pH用蒸餾水浸提(土水比1∶2.5)pH計測定,有機質OM采用重鉻酸鉀外加熱法,全氮TN用半微量凱氏定氮法,全磷TP用堿熔-鉬銻抗比色法,全鉀TK用NaOH熔融―火焰光度法,速效氮AN用堿解擴散法,有效磷AP用0.5 mol L-1NaHCO3浸提-鉬銻抗比色法,速效鉀AK用1.0 mol L-1NH4OAc 浸提―火焰光度法測定。
土壤基因組DNA的提取:利用試劑盒提取土壤基因組DNA,采用FastDNA SPIN Kit for Soil(MP Biomedicals公司,美國)試劑盒,使用超微量分光光度計(NanoDrop 2000,美國)檢測DNA的濃度和純度。DNA樣品保存于-20℃冰箱待用。
16S rRNA基因的高通量測序:采用通用引物519F(CAGCMGCCGCGGTAATWC)和907R(CCGTCAATTCMTTTRAGTTT)擴增細菌16S rRNA基因的V4~V5區。正向引物序列包含5 bp的barcode序列以區分樣品。 PCR反應體系為50 μl,包括25 μl的Taq DNA聚合酶(Takara),2 μl的正反向引物(10 μmol L-1),1 μl的DNA模板(約50 ng基因組),20 μl的 ddH2O。擴增條件為95 ℃預變性3 min,95 ℃變性45 s,56 ℃ 退火45 s,72 ℃ 延伸60 s,35個循環。最后72 ℃ 擴增7 min。PCR產物純化后,等物質的量混合后,通過Illumina Miseq平臺(Illumina公司,美國)進行高通量測序(中國科學院南京土壤研究所)。
1.4 數據處理
基于QIIME分析平臺對原始FASTQ文件進行過濾、拼接、去除嵌合體[16]。序列比對后按照97%相似性水平劃分操作分類單元(operational taxonomic unit,OTU)。每個OTU的最優勢序列選作代表性序列,通過RDP提供的參考序列和分類系統[17]進行分類。采用Mothur軟件計算多樣性指數(Chao1、Ace、香農指數和辛普森指數)[18]。使用R軟件基于Bray-Curits距離算法進行聚類分析。采用Canoco 5.0 軟件對環境因子與細菌群落結構進行冗余分析(Redundancy analysis,RDA)。采用SPSS 18.0進行統計分析,單因素方差分析(One-way ANOVA)檢驗多處理間均值差異的顯著性。
2 結 果
2.1 水旱輪作模式對冷浸田水稻產量的影響
不同水旱輪作模式對2015年冷浸田水稻產量產生了明顯的影響(圖1)。單季稻-冬閑(CK)處理產量最低,僅為5 737 kg hm-2;春玉米-水稻輪作(C-R)產量最高,為8 266 kg hm-2,其次為蠶豆-水稻輪作(B-R)、油菜-水稻輪作(R-R)和紫云英-水稻輪作(M-R),產量依次為7 737 kg hm-2、7 493 kg hm-2和6 783 kg hm-2。C-R、B-R、R-R和M-R輪作模式的產量分別是CK產量的1.44倍、1.35倍、1.31倍和1.18倍。
2.2 水旱輪作模式對冷浸田土壤理化性質的影響
由表1可知,水旱輪作模式對土壤有機質、全鉀和速效氮含量無顯著影響。B-R處理的土壤全氮含量最高,但是僅顯著高于R-R處理,與其他輪作模式無顯著差異。與CK相比,M-R處理的全磷含量與之無顯著差異;其他輪作模式的全磷含量均得到明顯提升(p< 0.05)。CK處理的有效磷含量最低,僅為14.92±0.72 mg kg-1,水旱輪作后土壤有效磷含量明顯增加(p< 0.05),R-R、C-R、M-R和B-R增幅分別為80.36%、89.48%、25.80%和79.83%。CK和M-R處理的速效鉀含量最低,R-R、C-R和B-R處理的速效鉀含量分別是CK的1.04倍、1.12倍和1.29倍。與CK處理相比,水旱輪作模式的土壤pH值顯著增加,一定程度上緩解了土壤酸化程度。

圖1 不同水旱輪作模式對水稻產量的影響Fig. 1 Effect of paddy-upland rotation on rice yield relative to mode of the rotation

表1 不同水旱輪作模式對冷浸田土壤理化性質的影響Table 1 Effect of paddy-upland rotation on soil physico-chemical properties relative to mode of the rotation
2.3 水旱輪作模式對冷浸田土壤細菌群落豐富度和多樣性的影響
Illumina高通量測序結果顯示,15個土壤樣品共獲得有效序列283 774條,每個樣品序列數為10 782~23 786條,為保證后續α和β多樣性的可比性,對所有樣品隨機抽取10 000條序列。Chao 1指數和OTU數目反映物種豐富度。由表2可知,CK處理土壤細菌Chao 1指數最高,達3 016±135,水旱輪作模式降低了土壤細菌的豐富度,R-R、C-R、M-R和B-R處理Chao1指數較冬閑對照分別降低13.65%、13.68%、4.21%和13.99%。OTU數目與Chao 1指數顯示相同的變化趨勢,即水旱輪作降低了土壤細菌OTU數目。香農和辛普森指數可反映土壤細菌群落的多樣性,表2顯示單季稻-冬閑和水旱輪作模式土壤細菌多樣性無顯著性差異(p> 0.05)。

表2 不同輪作模式下土壤細菌豐富度指數與多樣性指數Table 2 Diversity and richness of the soil bacteria in the soil relative to mode of the rotation
2.4 水旱輪作模式對冷浸田土壤細菌群落結構的影響
在門分類水平,各輪作處理的土壤細菌群落組成相似(相對豐度 >1%)(圖2)。主要包含變形菌門(P r o t e o b a c t e r i a)、酸桿菌門(A c i d o b a c t e r i a)、 擬桿菌門(Bacteroidetes)、綠彎菌門(Chloroflexi)、放線菌門(A c t i n o b a c t e r i a)、厚壁菌門(Firmicutes)、浮霉菌門(Planctomycetes),約占全部細菌豐度的71.88%~80.09%。此外,還含有裝甲菌門(Armatimonadetes)、藍藻門(Cyanobacteria)、迷蹤菌門(Elusimicrobia)、芽單胞菌門(Gemmatimonadetes)、硝化螺旋菌門(Nitrospirae)和疣微菌門(Verrucomicrobia)等相對豐度較低(<1%)的細菌群落。各樣品中還有14.92%~22.89%的菌,屬于目前分類學無法劃分的細菌種類。
變形菌門是土壤中最為優勢的菌群,占41.61%~44.82%。變形菌門的4個亞群的分布在各輪作處理間發生了變化(圖3)。與C K相比,水旱輪作土壤α-變形菌和γ-變形菌相對含量呈增加趨勢(p< 0.05),分別增加了49.42%~82.58%和38.92%~61.48%。而δ-變形菌相對豐度從CK處理的14.95%下降至水旱輪作處理的8.95%~11.29%。屬于α-變形菌的根瘤菌目(Rhizobiales)輪作后豐度顯著增加(p<0.05)。酸桿菌門(Acidobacteria)是冷浸田土壤中第2大優勢菌群,其豐度在CK處理中最低為14.18%,水旱輪作模式明顯增加了酸桿菌的豐度(p< 0.05),R-R、C-R、M-R和B-R分別較CK處理增加了62.36%、48.84%、33.48%和51.78%,表明水旱輪作的土壤環境更有利于酸桿菌門的生長。
在門分類水平還有部分細菌菌群的相對豐度也發生了顯著變化。例如,放線菌門(Actinobacteria)、硝化螺旋菌門(Nitrospirae)和綠彎菌門(Chloroflexi)在CK、R-R、C-R、M-R和B-R中的相對豐度分別為1.10%、1.40%、1.55%、2.01%、1.62%;0.35%、0.75%、0.98%、0.75%、0.81%和7.61%、4.59%、5.33%、4.84%、4.96%。與CK相比,水旱輪作土壤放線菌門和硝化螺旋菌門相對豐度分別增加27.27%~82.73%和114.3%~180.0%。而綠彎菌門(Chloroflexi)相對豐度則減少29.96%~39.68%。
土壤細菌群落在屬水平的分布也發生了改變(圖4)。各水旱輪作處理中土壤GP1、GP3、GP6、GP18、Sideroxydans、u n c l a s s i f i e d A l p h a p r o t e o b a c t e r i a、u n c l a s s i f i e d B e t a p r o t e o b a c t e r i a、u n c l a s s i f i e d D e l t a p r o t e o b a c t e r i a、unclassified Rhodospirillaceae、unclassified Rhizobiales、Pseudolabrys、Bradyrhizobium、Rhizomicrobium、unclassified Chloroflexi、unclassif ied Anaerolineaceae、unclassified Bacteroidetes的相對豐度> 1%。其中酸桿菌門包含的菌屬最多,GP1、GP3、GP6和GP18占全部菌屬的9.79%~17.19%。在不同輪作處理中有一些豐度發生顯著變化的屬,例如GP1、亞硝化螺菌屬(Nitrosospira)、慢生根瘤菌屬(Bradyrhizobium)、Rhizomicrobiu m在CK處理中含量最低,水旱輪作后相對豐度分別增加61.77%~265.3%、67.76%~113.6%、4.78%~60.75%和99.18%~194.6%;鐵氧化屬細菌(Sideroxydans)相對豐度在CK處理中最高,輪作后相對豐度減少33.68%~57.85%。

圖2 各水旱輪作處理土壤細菌群落在門水平上的組成和相對豐度Fig. 2 Composition and relative abundance of the bacterial community at the phylum level in the soil relative to mode of the rotation

圖3 各水旱輪作處理土壤變形菌亞群分布情況Fig. 3 Distribution of proteobacterial subgr oups in the soil relative to mode of the rotation
2.5 聚類分析
對樣品進行聚類分析,如圖5所示。樣品可以分為2大類,第1類:CK和M-R;第2類:R-R、C-R和B-R。單季稻-冬閑和紫云英-水稻輪作之間土壤細菌群落結構無明顯差異。玉米、蠶豆和油菜這3種旱作作物與水稻輪作對土壤細菌群落結構無明顯影響。
2.6 環境因子對水旱輪作冷浸田水稻土壤細菌群落結構的影響

圖4 各水旱輪作處理土壤細菌群落在屬水平上的組成和相對豐度Fig. 4 Composition and relative abundance of the bacterial community at the genus level in the soil relative to mode of the rotation
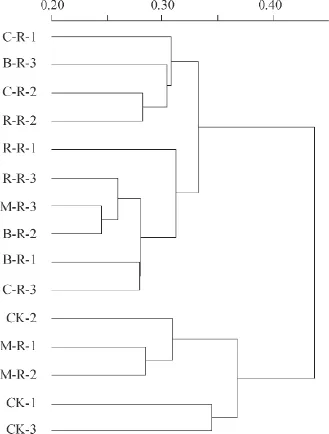
圖5 不同水旱輪作處理的土壤細菌群落結構相似度樹狀圖Fig.5 Dendrogram of hierarchical cluster analysis of the bacterial community in the soil relative to mode of the rotation
土壤理化因子和細菌群落多樣性相關性分析見表3。OTU數目、Chao1指數和理化因子間有著密切的相關性。其中OTU數目與TP、AP呈顯著負相關(p< 0.05);Chao1指數與TP和AP呈極顯著負相關(p< 0.01);OTU數目、Chao1指數與TK呈顯著正相關(p< 0.05)。用Spearman’s系數對土壤優勢菌(門水平)、變形菌亞群和土壤理化性質進行相關性分析(表4)。結果顯示α-變形菌的相對豐度與TP、pH呈極顯著正相關(p<0.01);δ-變形菌門與TP、AP呈顯著負相關(p< 0.05),與pH呈極顯著負相關(p< 0.01);γ-變形菌門相對豐度與AP呈極顯著正相關(p<0.01),與pH呈顯著正相關(p< 0.05);酸桿菌門的相對豐度與TP呈顯著正相關(p< 0.05),與pH呈極顯著正相關(p< 0.01),與TK呈顯著負相關(p< 0.05);綠彎菌門相對豐度與pH呈顯著負相關(p< 0.05)。

表3 土壤性質與細菌群落多樣性指數的相關性Table 3 Relationships between soil properties and bacterial diversity index

表4 土壤性質與細菌優勢菌的相關性Table 4 Relationships between soil properties and dominant bacterial taxa (RA >3% in all samples)
通過CANOCO軟件對環境因子和土壤細菌群落進行RDA分析。由圖6可知,第一排序軸和第二排序軸分別解釋了細菌群落變化的54.92%和10.75%,兩者之和達65.67%,表明本研究選擇的環境因子能較好地解釋土壤細菌群落。M-R和CK處理位于一、四象限,B-R、C-R和R-R處理位于二、三象限。經過蒙特卡羅檢驗,土壤pH(F=7.9,P= 0.002)、AP(F= 6.2,P= 0.002)和TP(F= 5.2,P= 0.018)對細菌群落結構的影響達到顯著水平。
3 討 論
單季稻-冬閑(CK)處理水稻產量最低,水旱輪作后產量呈增加趨勢:M-R< R-R< B-R< C-R。輪作增加水稻產量這一結論與Xuan等[11]的結果相一致。然而由于輪作作物種類和施入肥料的差異,水稻產量的增幅存在差異。C-R處理水稻產量最高為8 266 kg hm-2,較CK產量高44.12%。產量的增加可能與旱作期收獲物帶走的養分量低于施肥量和秸稈還田養分量有關[12]。水旱輪作后土壤部分理化因子也發生了變化。與CK相比,水旱輪作后土壤TP、AP、AK和pH明顯增加,這可能是由于旱作植物秸稈還田后可以當做肥料使用的原因。值得注意的是,紫云英-水稻輪作處理中種植紫云英時沒有施用氮肥,然而土壤TN和AN含量與其他輪作處理間并無顯著差異,這是由于綠肥的高效固氮能力和快速生物積累特性造成的[19]。CK處理土壤pH最低,水旱輪作后pH明顯增加,這在一定程度上緩解了土壤的酸化現象,這可能與旱作秸稈還田下鹽基離子釋放有關。
本研究采用高通量測序技術對冷浸田不同水旱輪作模式下土壤細菌群落多樣性進行了分析研究。結果顯示,與CK相比,水稻與紫云英、蠶豆、玉米和油菜輪作后土壤細菌的豐富度降低,各輪作處理間豐富度沒有顯著差異。水旱輪作后土壤細菌多樣性指數與冬閑處理無顯著差異(表2)。以上結果表明水稻產量與土壤的細菌多樣性之間并沒有明顯的因果關系[11]。較低的土壤細菌多樣性未必表示差的生態系統,較低的多樣性也未必暗示著它們是不穩定或不健康的微生物群落[20]。
相比水稻-冬閑處理,水旱輪作并未顯著改變冷浸田土壤的細菌群落結構,只是部分類別的細菌相對豐度發生變化。在門分類水平,土壤細菌以變形菌門為絕對優勢菌,占細菌群落的41.61%~44.82%。酸桿菌門廣泛存在于自然界的各種環境中,是土壤微生物的重要成員,約占土壤細菌類群的5%~46%[21]。本研究中酸桿菌門的相對豐度為18.92%~23.02%,僅次于變形菌門,這與Fang等[22]的研究相一致。土地種植方式的改變對土壤微生物群落結構會產生顯著影響。例如,輪作后土壤變形菌的亞群分布發生了明顯變化,α-變形菌和γ-變形菌從CK處理的8.67%、3.32%增加至輪作后的12.92%~15.79%和4.61%~5.37%。Rousk等[23]報道α-變形菌和γ-變形菌的相對豐度隨著pH的增加而增多,本研究結果也顯示輪作后土壤pH增加,同時α-變形菌和γ-變形菌的相對豐度也得到明顯提升(表1和圖3)。α-變形菌中的慢生根瘤菌屬(Bradyrhizobium)在輪作土壤中豐度顯著增加,它們是水稻生態系統中活躍的固氮細菌[24]。而δ-變形菌相對含量在CK處理中含量最高(14.95%),輪作后豐度明顯降低(8.95%~11.29%)。水旱輪作后酸桿菌豐度明顯增加,其相對含量與土壤全磷含量呈顯著正相關(p< 0.05),而輪作處理(除M-R處理)能夠顯著增加土壤全磷含量。Eichorst等[25]報道酸桿菌能夠降解復雜的碳化合物例如纖維素和木聚糖等。輪作處理中旱作植物秸稈還田增加了土壤中纖維素等復雜碳化合物的輸入,導致酸桿菌的增多。同時本研究發現酸桿菌與土壤pH呈極顯著正相關,而Mannisto[26]和Griffiths[27]等的研究結果表明酸桿菌豐度與pH呈顯著負相關。也有研究表明,酸桿菌相對豐度與土壤pH相關性不顯著[28]。由于一些酸桿菌的基因序列也在中性、甚至堿性的環境中被檢測出來[29],所以酸桿菌相對含量和pH的關系還有待進一步研究。與CK相比,水旱輪作后土壤放線菌和硝化螺旋菌相對豐度明顯增加。放線菌能夠降解復雜的木質素[30]和纖維素[31]。旱作植物秸稈還田能夠增加木質素和纖維素的輸入,從而刺激了放線菌的生長。硝化螺旋菌可將亞硝酸鹽氧化成硝酸鹽,提高土壤氮素的利用率,從而促進水稻的生長,增加其產量。與CK相比,水旱輪作增加了慢生根瘤菌屬以及亞硝化螺菌屬的豐度,這些菌屬與土壤氮循環(固氮作用和硝化作用第一步)均有著密切的聯系。

圖6 土壤細菌群落結構和土壤理化因子的RDA分析Fig.6 Redundancy analysis (RDA) of soil bacterial community structure and soil properties
冷浸田土壤由于其自身的生態特征,使之區別于高產田。王秀呈[32]報道水稻-綠肥輪作下紅壤水稻土細菌類群主要為變形菌門、酸桿菌門、放線菌門、厚壁菌門、擬桿菌門、綠彎菌門、疣微菌門、浮霉菌門以及芽單胞菌門。本研究中冷浸田土壤細菌主要類群與之類似,然而某些類別的細菌豐度存在明顯差異。例如冷浸田輪作土壤放線菌豐度明顯低于紅壤水稻土,可能是由于放線菌一般是好氣性微生物,而冷浸田土壤長期處于還原性條件,不利于放線菌的生長。
許多研究表明環境因子能夠影響微生物群落結構[33-34]。本研究發現細菌的某些優勢菌和變形菌亞群與土壤pH高度相關。許多研究已經證實土壤pH是影響細菌群落結構的重要因子[35-36]。此外,土壤的其他理化性質如TP、AP和細菌的某些優勢菌和變形菌亞群也呈顯著相關(表4)。例如,α-變形菌與土壤TP呈極顯著正相關,這就解釋了CK和M-R處理中含有較低豐度的α-變形菌。由RDA分析可知,土壤pH、全磷和有效磷是影響水旱輪作土壤細菌群落結構的主要因子。秦紅等[37]研究結果也表明全磷和有效磷的含量是影響細菌群落結構的重要因素。B-R、C-R和R-R處理的土壤全磷和有效磷含量均顯著高于CK和M-R處理。這可能是由于B-R、C-R和R-R輪作模式下磷肥的盈余(收獲物帶走的磷低于施肥量)高于CK和M-R處理[12],從而導致了土壤細菌群落結構的變化。
4 結 論
與冬閑對照相比,水旱輪作通過提高土壤全磷、有效磷、速效鉀含量,緩解土壤酸化程度而增加水稻產量。水旱輪作土壤細菌豐富度降低,多樣性指數與對照沒有顯著差異。變形菌門和酸桿菌門為研究區土壤中的優勢細菌類群。輪作后酸桿菌、放線菌、硝化螺旋菌、慢生根瘤菌以及亞硝化螺菌等參與碳氮循環的菌群豐度顯著增加。土壤pH、AP和TP是影響水旱輪作冷浸田土壤細菌群落分布的主要因子。
[ 1 ] Dickopp J,Kazda M,?í?ková H. Differences in rhizome aeration ofPhragmitesaustralisin a constructed wetland. Ecological Engineering,2011,37(11):1647—1653
[ 2 ] 曾燕,黃敏,蔣鵬,等. 冷浸田條件下不同類型品種的表現和高產栽培方式研究. 作物研究,2010,24(3):140—144 Zeng Y,Huang M,Jiang P,et al. Studies on yield performance and high yielding cultivation methods of different types of cultivars in cold water paddy field(In Chinese). Crop Research,2010,24(3):140—144
[ 3 ] 柴娟娟,廖敏,徐培智,等. 我國主要低產水稻冷浸田養分障礙因子特征分析. 水土保持學報,2012,26(2):284—288 Chai J J,Liao M,Xu P Z,et al. Feature analysis on nutrient’s restrictive factors of major low productive waterlogged paddy soil in China(In Chinese).Journal of Soil Water Conservation,2012,26(2):284—288
[ 4 ] 龔子同,張效樸,韋啟璠. 我國潛育性水稻土的形成、特性及增產潛力. 中國農業科學,1990,23(1):45—53 Gong Z T,Zhang X P,Wei Q F. The formation,characterization and potential of gleyic paddy soils in China(In Chinese). Scientia Agricultura Sinica,1990,23(1):45—53
[ 5 ] Costa R,G?tz M,Mrotzek N,et al. Effects of site and plant species on rhizosphere community structure as revealed by molecular analysis of microbial guilds.FEMS Microbiology Ecology,2006,56(2):236—249
[ 6 ] Kent A D,Triplett E W. Microibal communities and their interactions in soil and rhizosphere ecosystems.Annual Review of Microbiology,2002,56(1):211—236
[ 7 ] Jacobsen C S,Hjelms? M H. Agricultural soils,pesticides and microbial diversity. Current Opinion in Biotechnology,2014,27(5):15—20
[ 8 ] 張薇,魏海雷,高洪文,等. 土壤微生物多樣性及其環境影響因子研究進展. 生態學雜志,2005,24(1):48—52 Zhang W,Wei H L,Gao H W,et al. Advances of studies on soil microbial diversity and environmental impact factors(In Chinese). Chinese Journal of Ecology,2005,24(1):48—52
[ 9 ] Yin C T,Jones K L,Peterson D E,et al. Members of soil bacterial communities sensitive to tillage and crop rotation. Soil Biology & Biochemistry,2010,42:2111—2118
[10] Navarro—Noya Y E,Gómez—Acata S,Montoya—Ciriaco N,et al. Relative impacts of tillage,residue management and crop—rotation on soil bacterial communities in a semi—arid agroecosystem. Soil Biology & Biochemistry,2013,65:86—95
[11] Xuan D T,Guong V T,Rosling A,et al. Different crop rotation systems as drivers of change in soil bacterial community structure and yield of rice,Oryzasativa. Biology and Fertility of Soils,2012,48(2):217—225
[12] 王飛,李清華,林誠,等. 冷浸田水旱輪作對作物生產及土壤特性的影響. 應用生態學報,2015,26(5):1469—1476 Wang F,Li Q H,Lin C,et al. Influences of paddy rice—upland crop rotation of cold—waterlogged paddy field on crops production and soil characteristics(In Chinese). Chinese Journal of Applied Ecology,2015,26(5):1469—1476
[13] 李清華,王飛,林誠,等. 水旱輪作對冷浸田土壤碳、氮、磷養分活化的影響. 水土保持學報,2015,29(6):113—117 Li Q H,Wang F,Lin C,et al. Effects of paddy—upland rotation on the nutrient activation of soil carbon,nitrogen and phosphorus in cold waterlogged paddy field(In Chinese). Journal of Soil and Water Conservation,2015,29(6):113—117
[14] 林海波,陳正剛,張欽,等. 不同耕作措施對冷浸田亞鐵和水稻產量的影響. 廣東農業科學,2015(8):1—4 Lin H B,Chen Z G,Zhang Q,et al. Effects of different tillage practices on ferrous and rice yield in cold waterlogged paddy field(In Chinese).Guangdong Agricultural Sciences,2015(8):1—4
[15] 魯如坤. 土壤農化分析方法. 北京:中國農業科學技術出版社,2000 Lu R K. Analytical methods for soil and agrochemistry(In Chinese). Beijing:China Agricultural Science and Technology Press,2000
[16] Zhou J,Guan D W,Zhou B K,et al. Influence of 34—years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China.Soil Biology and Biochemistry,2015,90:42—51
[17] Wang Q,Garrity G M,Tiedje J M,et al. Na?ve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied and Environmental Microbiology,2007,73(16):5261—5267
[18] Schloss P D,Gevers D,Westcott S L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA—based studies. PLoS ONE,2011,DOI:10.1371/journal.pone.0027310
[19] Lee C H,Park K D,Jung K Y,et al. Effect of Chinese milk vetch(AstragalussinicusL.)as a green manure on rice productivity and methane emission in paddy soil. Agriculture,Ecosystems and Environment,2010,138(3/4):343—347
[20] Shade A. Diversity is the question,not the answer.The ISME Journal,2017,11(1):1—6
[21] Ellis R J,Morgan P,Weightman A J,et al.Cultivation—dependent and independent approaches for determining bacterial diversity in heavy—metal contaminated soil. Applied and Environmental Microbiology,2003,69(6):3223—3230
[22] Fang Y,Wang F,Chen J J,et al. Microbial responses in a cold waterlogged paddy field to different distances from a drainage ditch. Chilean Journal of Agricultural Research,2017,77(1):87—94
[23] Rousk J,Baath E,Brookes P C,et al. Soil bacterial and fungal communities across a pH gradient in an arable soil. The ISME Journal,2010,4(10):1340—1351
[24] C h a i n t r e u i l C,G i r a u d E,P r i n Y,e t a l.PhotosyntheticBradyrhizobiaare natural endophytes of the African wild riceOryzabreviligulata. Applied and Environmental Microbiology,2000,66(12):5437—5447
[25] Eichorst S A,Kuske C R,Schmidt T M. Inf l uence of plant polymers on the distribution and cultivation of bacteria in the phylumAcidobacteria. Applied and Environmental Microbiology,2011,77(2):586—596
[26] Mannisto M K,Tiirola M,Haggblom M M. Bacterial communities in Arctic fields of Finnish Lapland are stable but highly pH—dependent. FEMS Microbiology Ecology,2007,59(2):452—465
[27] Griffiths R I,Thomson B C,James P,et al. The bacterial biogeography of British soils. Environmental Microbiology,2011,13(6):1642—1654
[28] Liu J J,Sui Y Y,Yu Z H,et al. Soil carbon content drives the biogeographical distribution of fungal communities in the black soil zone of Northeast China.Soil Biology & Biochemistry,2015,83:29—39
[29] Xiong J B,Liu Y Q,Lin X G,et al. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environmental Microbiology,2012,14(9):2457—2466
[30] Lynd L R,Weimer P J,Zyl W H V,et al. Microbial cellulose utilization:Fundamentals and biotechnology.Microbiology and Molecular Biology Reviews,2002,66(3):506—577
[31] Pankratov T A,Ivanova A O,Dedysh S N,et al. Bacterial populations and environmental factors controlling cellulose degradation in an acidicsphagnumpeat. Environmental Microbiology,2011,13(7):1800—1814
[32] 王秀呈. 稻—稻—綠肥長期輪作對水稻土壤及根系細菌群落的影響. 北京:中國農業科學院,2015 Wang X C. The influence of long term rice—rice—green manure rotation on rice bulk soil and root—associated bacteria(In Chinese). Beijing:Chinese Academy of Agricultural Sciences,2015
[33] 沈冰潔,祝貞科,袁紅朝,等. 不同種植方式對亞熱帶紅壤微生物多樣性的影響. 環境科學,2015,36(10):3839—3844 Shen B J,Zhu Z K,Yuan H C,et al. Effects of different plantation type on the abundance and diversity of soil microbes in subtropical red soils(In Chinese).Environmental Science,2015,36(10):3839—3844
[34] Jiang Y J,Liang Y T,Li C M,et al. Crop rotations alter bacterial and fungal diversity in paddy soils across East Asia. Soil Biology & Biochemistry,2016,95:250—261
[35] Lauber C L,Hamady M,Knight R,et al.Pyrosequencing—based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Applied and Environmental Microbiology,2009,75(15):5111—5120
[36] Chu H,Fierer N,Lauber C L,et al. Soil bacterial diversity in the Arctic is not fundamentally different from that found in other biomes. Environmental Microbiology,2010,12(11):2998—3006
[37] 秦紅,李昌曉,任慶水. 不同土地利用方式對三峽庫區消落帶土壤細菌和真菌多樣性的影響. 生態學報,2017,37(10):3494—3504 Qin H,Li C X,Ren Q S. Effects of different land use patterns on soil bacterial and fungal biodiversity in the hydro—fluctuation zone of the Three Gorges Reservoir region(In Chinese). Acta Ecologica Sinica,2017,37(10):3494—3504
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
中華詩詞(2020年1期)2020-09-21 09:24:52
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
小學生作文(中高年級適用)(2018年5期)2018-06-11 01:22:56
數學小靈通·3-4年級(2017年10期)2017-11-08 08:42:59
中學生數理化·七年級數學人教版(2017年11期)2017-04-23 07:18:00
