3個玉米WRKY轉錄因子在非生物脅迫下的表達分析
2018-04-08 06:37:47決登偉桑雪蓮劉麗琴王一承石勝友
西北農(nóng)業(yè)學報 2018年3期
決登偉,桑雪蓮,舒 波,劉麗琴,王一承,石勝友
(中國熱帶農(nóng)業(yè)科學院 南亞熱帶作物研究所,農(nóng)業(yè)部熱帶果樹生物學重點實驗室,廣東湛江 524091)
轉錄因子(transcription factor,TF)是一類能與真核基因啟動子區(qū)域中的順式作用元件特異性結合的蛋白分子。通過它們之間及與其他蛋白間的相互作用,實現(xiàn)對基因激活或者抑制轉錄,從而保證目的基因以特定的強度、在特定的時間與空間表達[1]。植物體內(nèi)含有大量的轉錄因子,根據(jù)序列特征及功能的差異,可以將它們分為很多不同的類型或家族。比如WRKY、AP2/ERF和MYB等。這其中,WRKY是植物體中最大的轉錄因子家族之一,它們在植物的生長發(fā)育及響應脅迫過程中起到重要作用。第一個WRKY轉錄因子(SPF1)在1994年從甘薯中鑒定出來[2]。隨后,在眾多植物中都出現(xiàn)報道,且其數(shù)目從低等植物到高等植物呈現(xiàn)上升趨勢。比如,在油菜 (BrassicanapusL.)中有46個WRKY轉錄因子成員[3],黃瓜(Cucumissativus)中有57個[4],麻風樹(Jatrophacurcas)中有58個[5],蓖麻(RicinuscommunisL.)中有58個[6],擬南芥(Arabidopsisthaliana)中有72個[7],白梨(Pyrusbretschneideri)中有103個[8],白楊(Populustrichocarpa)中有105個[9],谷子(Setariaitalica)中有105個[10],水稻(Oryzasativa)中有109個[11],在棉花(Gossypium)中有109~112個[12],玉米(ZeamayL.)中有136個[13],大豆(Glycinemax)中有197個[14]。
WRKY蛋白在結構上有一個顯著特點:都包含一個或者兩個WRKY結構域,包括N-端高度保守的WRKYGQK七肽及C-端的鋅指結構(Cx4-7Cx22-23HxH/C)。根據(jù)WRKY結構域的數(shù)目及鋅指結構的特點,可將WRKY蛋白劃分為3個主要類型。第Ⅰ類有兩個WRKY結構域,鋅指結構類型為C2H2。第Ⅱ和Ⅲ類只含有一個WRKY結構域。其中,第Ⅲ類成員的鋅指結構類型為C2HC,而第Ⅱ類為C2H2。根據(jù)進化關系及WRKY結構域中某些氨基酸基序不同,第Ⅱ類WRKY轉錄因子又可細分為5個小類(a~e)[15]。研究表明,WRKY轉錄因子參與植物多種生理生化過程,如應對生物和非生物脅迫,參與葉片衰老,發(fā)育和次生代謝等[16]。在擬南芥中,AtWRKY25和AtWRKY33受鹽的促進表達,且異源表達這兩個基因,增強植物對NaCl的耐性[17-18]。 AtWRKY70既可以激活SA介導的抗病信號途徑,同時也能抑制JA介導的信號途徑,從而實現(xiàn)對擬南芥抗病反應的調(diào)控[19]。在水稻中,過表達 OsWRKY45促進擬南芥對干旱的耐性, OsWRKY8異源表達促進擬南芥對滲透的耐性, OsWRKY72的異源表達則會影響根的生長和逆境耐性[20]。另外, OsWRKY89的過表達促進植株對紫外線的抵御[21],過表達 OsWRKY11增強轉基因植株對高溫的耐性[18]。在白梨的103 WRKY基因中,44個PbWRKY在干旱處理下呈上調(diào)表達[8]。高粱(PanicummiliaceumL.)中,10個PmWRKY基因的表達受到干旱脅迫誘導,16個PmWRKY基因的表達受到低溫脅迫誘導[22]。玉米(ZeamaysL.)是當今世界重要的糧食作物之一,亦是重要的飼料和工業(yè)原料作物, 在世界糧食總產(chǎn)量中處于第一位[23]。中國作為世界第二大玉米生產(chǎn)國,近30多年來,玉米生產(chǎn)發(fā)展非常迅速,總產(chǎn)量也呈逐年上升趨勢。世界糧農(nóng)組織統(tǒng)計數(shù)據(jù)庫(FAOSTAT)顯示在2012 年,玉米產(chǎn)量超過稻谷產(chǎn)量,成為中國第一大糧食作物[24]。2014年,中國玉米常年種植面積達3 715.04 萬hm2,總產(chǎn)量約2.16 億t,占同年中國糧食總產(chǎn)量(6.07億t)的1/3。但在實際生產(chǎn)中,玉米常因受到諸多不利環(huán)境因素的影響而大面積減產(chǎn),比如干旱、鹽脅迫和低溫等。目前,已有關于WRKY家族基因的研究文章[13],但還沒有關于 ZmWRKY1-like、 ZmWRKY4-like、 ZmWRKY21-like基因在植物響應非生物脅迫中作用的報道。本研究采用生物信息學技術從玉米的基因組發(fā)現(xiàn)3個WRKY家族基因: ZmWRKY1-like、 ZmWRKY4-like、 ZmWRKY21-like。利用熒光定量PCR技術,探討這3個基因在玉米不同組織、干旱、鹽脅迫及低溫脅迫的表達模式。該研究可以為挖掘玉米抗逆基因及豐富WRKY轉錄因子在不同作物中的功能研究提供一定的理論和試驗依據(jù)。
1 材料與方法
1.1 試驗材料及處理
玉米自交系B73種子保存于南亞熱帶作物所。玉米組織根(root)、莖(stem)、葉(leaf)、穗(tassel)、幼果(young seed)和穗絲(silk)取自大田種植的玉米自交系B73植株,采樣時期為乳熟期。所用植物RNA提取試劑盒購自北京華越洋生物公司(Huayueyang Bio Co., Ltd, Beijing, China);反轉錄試劑及Realtime PCR試劑盒購自大連寶生物工程有限公司(TaKaRa Bio, Inc., Dalian, China);引物由北京賽百盛基因技術有限公司合成;其他試劑購自上海生物工程有限公司。逆境處理所用材料為B73種子萌發(fā)而來的幼苗,種子萌發(fā)后,轉入1/2 Hoagland 培養(yǎng)液,在培養(yǎng)箱中繼續(xù)培養(yǎng),生長條件為(28±2) ℃,14 h光照,10 h黑暗。3周后,選擇長勢均一的幼苗進行下步試驗。每個處理具體方法如下:①鹽脅迫處理,將幼苗放入含200 mol/L NaCl的1/2 Hoagland培養(yǎng)液中培養(yǎng),按0、1、6和24 h時間點取樣;②干旱脅迫處理,將幼苗放入含200 g/L PEG6000的1/2 Hoagland培養(yǎng)液中培養(yǎng),按0、1、6和24 h時間點取樣;③低溫脅迫處理,將幼苗放入溫度設置為4 ℃的培養(yǎng)箱中培養(yǎng),按0、1、6和24 h時間點取樣。設3次重復,取樣部位為葉片。所有取樣剪成2 cm左右的小段,立即放入液氮速凍并轉入-80 ℃冰箱中保存,備用。
1.2 試驗方法
1.2.1 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like序列的獲得及分析 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like3個基因序列從Phytozome (http://bioinformatics.psb.ugent.be/plaza/versions/plaza/)數(shù)據(jù)庫中獲得,并在玉米基因組數(shù)據(jù)庫(MaizeGDB, http://www. maizegdb.org/)中進行驗證。3個基因的ORF、氨基酸長度和染色體定位信息從Phytozome數(shù)據(jù)庫獲得。應用在線軟件SMART (http://smart.emblheidelberg.de/)預測3個蛋白的結構域;通過ExPASy (http://expasy.org/tools/)分析3個蛋白的等電點和分子質(zhì)量;通過Plant-mPLoc (http://www.csbio.sjtu.edu.cn/cgibin/ PlantmPLoc.cgi)預測3個蛋白的亞細胞定位情況;采用Gene Structure Display Server (GSDS) (http://gsds.cbi.pku.edu. cn/)分析3個基因的外顯子和內(nèi)含子結構;用Clustal X進行不同序列的多重序列對比,同時利用MEGA 6軟件進行氨基酸序列同源性分析及系統(tǒng)發(fā)育分析,構建Neighbor-Joining進化樹,1 000 次重復,其他均為默認設置。
1.2.2植物總RNA提取及Realtime PCR分析用北京華越洋生物公司的植物RNA提取試劑盒提取玉米不同組織材料的RNA,然后用TaKaRa公司的PrimeScript RT試劑盒反轉錄成cDNA后作為模板,具體操作步驟參照說明書。根據(jù)玉米基因組中 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like3個基因的CDS序列設計Realtime PCR引物,并以玉米的 Actin 2(NM_001154731)基因作為內(nèi)參基因,具體引物序列見表1。

表1 ZmWRKY基因定量PCR所用引物Table 1 Primers used in quantitative RT-PCR of ZmWRKY gene
Realtime PCR反應在Roche的LightCycler480儀器中進行,PCR反應酶為TaKaRa公司的SYBR Green Master Mix。反應體系為20 mL,其中模板cDNA 40 ng,上、下游引物各250 nmol/L,SYBR Green Master Mix 10 μL,用ddH2O 補齊。反應程序:94 ℃預變性5 min;94 ℃ 10 s,59 ℃ 20 s,72 ℃ 30 s,40 個循環(huán)后作熔解曲線(95 → 65 ℃,0.1 ℃/s)。利用2-ΔΔCt計算基因的相對表達量。所有樣品進行3次重復,均設陰性對照。在分析基因的表達時,上調(diào)或者下調(diào)大于2倍時才認為存在差異。
2 結果與分析
2.1 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like3個基因的獲得及生物信息學分析
以WRKY轉錄因子保守區(qū)氨基酸序列在Phytozome和MaizeGDB進行BLASTP搜索,通過分析獲得3個玉米同源性較高的WRKY轉錄因子家族基因,分別為 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like。3個基因的基因id、locus、染色體定位等信息見表2和圖1。其中, ZmWRKY1-like基因序列開放閱讀框為1 194 bp,編碼的蛋白具有397個氨基酸,其分子質(zhì)量為42.80 ku,理論等電點為9.98,位于玉米第Ⅰ號染色體; ZmWRKY4-like基因序列開放閱讀框為1 701 bp,編碼的蛋白具有566個氨基酸,其分子質(zhì)量為61.71 ku,理論等電點為6.20,位于玉米第Ⅳ號染色體; ZmWRKY21-like基因序列開放閱讀框為1 065 bp,編碼的蛋白具有354個氨基酸,其分子質(zhì)量為38.04 ku,理論等電點為9.81,位于玉米第Ⅰ號染色體。預測表明3個基因都位于細胞核。對3個基因的氨基酸序列進行分析,發(fā)現(xiàn) ZmWRKY4-like含有兩個WRKY結構域,根據(jù)WRKY轉錄因子分類原則,歸為第Ⅰ類。 ZmWRKY1-like和 ZmWRKY21-like都含有1個WRKY結構域,鋅指結構同為C-X5-C-X23-H-X1-H型,屬于第Ⅱd類WRKY轉錄因子。

表2 ZmWRKY基因信息Table 2 Information of ZmWRKY genes
2.2 ZmWRKY1-like、ZmWRKY4-like和ZmWRKY21-like的系統(tǒng)進化分析
采用MEGA 6軟件將玉米ZmWRKY1-like、ZmWRKY4-like和ZmWRKY21-like蛋白序列與擬南芥、水稻、小米、小麥的部分WRKY蛋白序列進行分析并構建進化樹(圖 2),發(fā)現(xiàn)ZmWRKY1-like和ZmWRKY21-like都聚到Ⅱd分支中,與小米的SiWRKY1及小麥的TaWRKY16同源性最高,與同分支擬南芥及水稻的WRKY蛋白同源性稍低。ZmWRKY4-like屬于Ⅰ分支,與同分支中小麥的TaWRKY45同源性最高,與擬南芥的AtWRKY25、AtWRKY26、AtWRKY33,水稻的OsWRKY24、OsWRKY53、OsWRKY70同源性較低(圖 2)。

A.保守結構域蛋白序列對比,黑框中為WRKY結構Alignment of conserved motifs of ZmWRKYS.The WRKY conserved motifis are bordered(black rectangles);B.基因結構,藍色為外顯子,黑色為內(nèi)含子Gene structure of ZmWRKYs is presented by blue exons and gray intron between the blue box
圖1ZmWRKY1-like、ZmWRKY4-like和ZmWRKY21-like保守結構域蛋白序列對比及基因結構
Fig.1AlignmentofconservedmotifsandgenestructureofZmWRKYs
2.3 玉米不同組織 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like的表達分析
以大田種植的B73植株為材料,采樣時期為乳熟期。用Realtime PCR技術分析 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like3個基因在玉米幼苗的根(root)、莖(stem)、葉(leaf)、穗(tassel)、幼果(young seed)和穗絲(silk)中的表達情況。圖3結果顯示,3個基因在玉米不同組織中都表達,但存在差異。其中, ZmWRKY1-like和 ZmWRKY21-like基因在穗絲中的表達量最低, ZmWRKY4-like基因在根中的表達量最低。3個基因在幼果中的表達量都顯著高于在其他組織, ZmWRKY1-like和 ZmWRKY21-like在幼果中的表達量分別是根的65和13倍; ZmWRKY4-like在幼果中的表達量是穗絲的27倍。說明 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like這3個基因可能參與玉米植株不同組織器官的發(fā)育過程,特別是玉米籽粒的發(fā)育過程。
2.4 不同非生物逆境脅迫下 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like的表達分析
以3周苗齡的B73玉米幼苗為材料,用Realtime PCR技術分析 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like3個基因在鹽、干旱和低溫處理下表達情況。如圖4、5、6所示,鹽脅迫下, ZmWRKY1-like基因在6 h時出現(xiàn)下調(diào)表達,為對照的0.43倍,在24 h時恢復到對照水平; ZmWRKY4-like基因的表達在檢測時間內(nèi)未出現(xiàn)變化; ZmWRKY21-like基因在24 h時呈現(xiàn)上調(diào)表達,為對照的2.2倍。干旱脅迫下, ZmWRKY1-like基因在6和24 h出現(xiàn)下調(diào)表達,分別為對照的0.3和0.4倍; ZmWRKY4-like基因的表達在檢測時間內(nèi)未出現(xiàn)變化; ZmWRKY21-like基因在6 h時出現(xiàn)下調(diào)表達,為對照的0.4倍,在24 h時恢復到對照水平。低溫脅迫下,3個基因的表達都沒有出現(xiàn)顯著變化。上述結果表明, ZmWRKY1-like和 ZmWRKY21-like基因可能分別參與植物對干旱和鹽脅迫的響應。
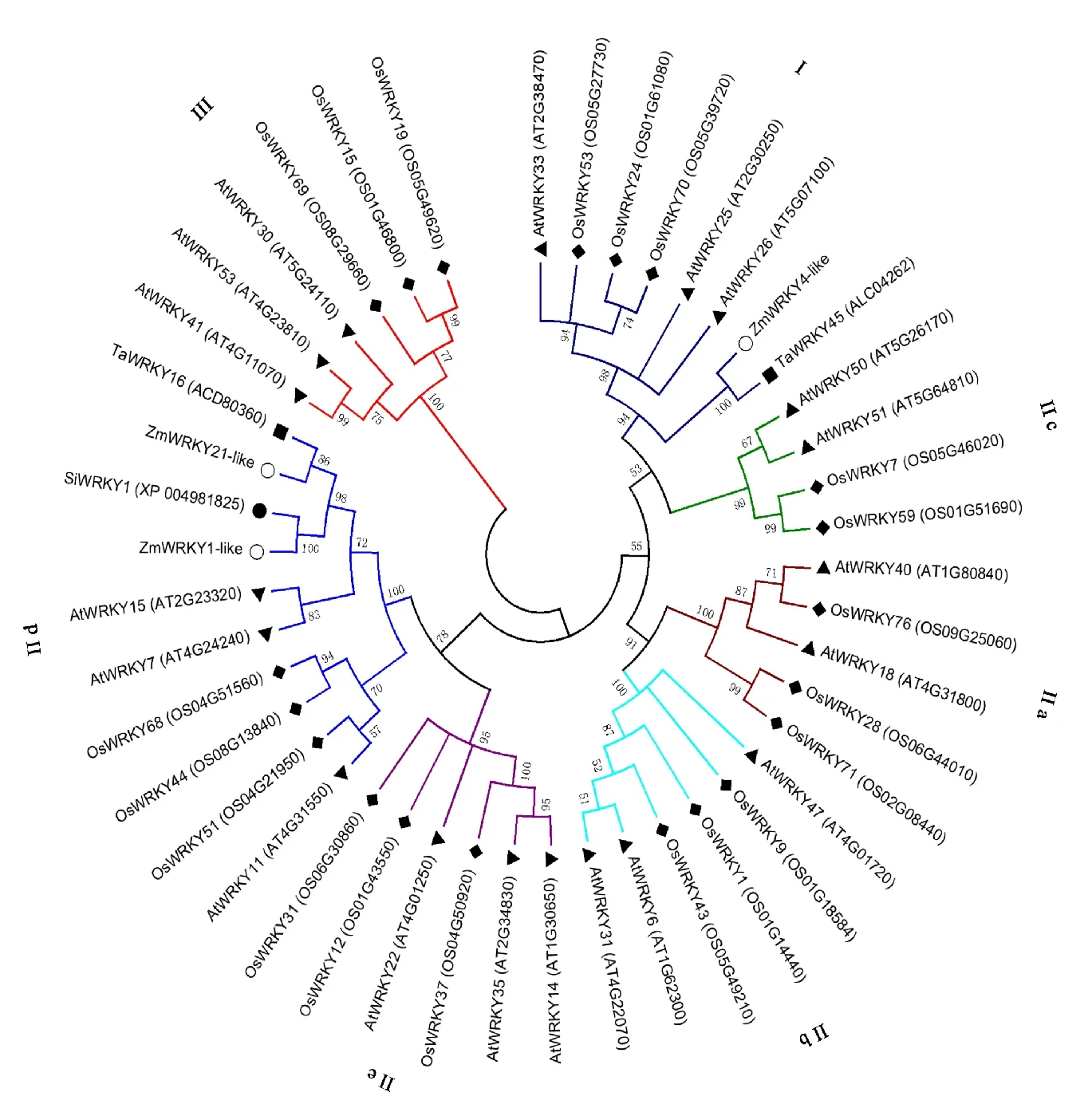
○玉米 Zea mays;▲擬南芥 Arabidopsis thaliane; ● 小米 Setaria italica;◆水稻 Oryza sativa ;■小麥 Triticum aestivum

圖3 ZmWRKY1-like、ZmWRKY4-like和ZmWRKY21-like基因在玉米不同組織中的表達水平Fig.3 Expression level of ZmWRKY1-like, ZmWRKY4-like and ZmWRKY21-like gene in different maize tissues

圖4 玉米 ZmWRKY1-like基因的逆境表達分析Fig.4 Stress expression analysis of ZmWRKY1-like gene

圖5 玉米 ZmWRKY4-like基因的逆境表達分析Fig.5 Stress expression analysis of ZmWRKY4-like gene

圖6 玉米 ZmWRKY21-like基因的逆境表達分析Fig.6 Stress expression analysis of ZmWRKY21-like gene
3 結論與討論
WRKY是植物中一類重要的轉錄因子家族,也是植物轉錄因子中最大的家族之一[25]。本研究從Phytozome和MaizeGDB數(shù)據(jù)庫中得到3個玉米WRKY家族基因: ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like。對3個基因的序列特征和進化關系進行分析,并采用熒光定量PCR檢測這3個基因在玉米不同組織和不同逆境脅迫處理下的表達情況。
分析表明ZmWRKY1-like、ZmWRKY4-like和ZmWRKY21-like都具有WRKY保守結構域和鋅指結構,分別為第Ⅱd、Ⅰ和Ⅱd類WRKY轉錄因子。石紅梅等[26]的研究表明,PsWRKY基因主要在牡丹的葉片和心皮中表達,在其他器官中的表達量非常低。煙草的164個WRKY基因中,大部分在根、莖、葉中都有表達,但不同基因間的表達模式存在差異。比如: NtWRKY7、NtWRKY11、NtWRKY32、NtWRKY82、NtWRKY92、NtWRKY105和 NtWRKY147在根、莖、葉中不表達或表達量較低; NtWRKY26和 NtWRKY78在根中表達量最高; NtWRKY21和 NtWRKY114在莖中表達量最高; NtWRKY28、 NtWRKY81、 NtWRKY163在葉片中表達量最高[27]。與這些研究結果類似,本研究中 ZmWRKY1-like、 ZmWRKY4-like和 ZmWRKY21-like3個基因在檢測的玉米組織中都有表達,呈多種表達模式。其中, ZmWRKY1-like和 ZmWRKY21-like基因在穗絲中的表達量最低, ZmWRKY4-like基因在根中的表達量最低。3個基因在幼果中的表達量都顯著高于在其他組織中的表達量,特別是 ZmWRKY1-like在幼果中的表達量達到根中表達量的65倍。該結果說明這3個基因可能參與玉米植株不同組織器官的發(fā)育過程,特別是玉米籽粒的發(fā)育過程。
大量研究表明,WRKY蛋白參與植物對逆境脅迫的響應過程[16]。比如在擬南芥中,有近2 000 個干旱響應基因,其中就包含不少WRKY基因[28]。干旱和高鹽處理時, AtWRKY25和 AtWRKY33 的表達量會提高,而它們的異源表達增強轉基因植株對NaCl耐性[29]。高溫脅迫誘導 AtWRKY25和 AtWRKY26的表達,而抑制 AtWRKY33 的表達[30]。過表達 OsWRKY45可以增強轉基因植株對鹽和干旱的耐受性[31]。過表達 GmWRKY21可以增強轉基因擬南芥植株對低溫的耐受性[32]。在本研究中,高鹽處理下, ZmWRKY55-like基因出現(xiàn)上調(diào)表達,24 h為對照的2.2倍。 ZmWRKY1-like基因在干旱脅迫處理的24 h時表達量下調(diào)為對照的0.4倍。該結果表明, ZmWRKY1-like和 ZmWRKY21-like基因可能分別參與玉米植株對干旱和鹽脅迫的響應。但具體的分子機制仍需要進一步研究分析。
參考文獻Reference:
[1]劉強,張貴友.植物轉錄因子的結構與調(diào)控作用[J].科學通報,2000,45(14):1465-1474.
LIU Q,ZHANG G Y.Structure and regulation of plant transcription factors [J].ChineseScienceBulletin,2000,45(14):1465-1474.
[2]ISHIGURO S,NAKAMURA K.Characterization of a cDNA encoding a novel DNA-binding protein,SPF1,that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and β-amylase from sweet potato[J].MolecularandGeneralGeneticsMGG,1994,244(6):563-571.
[3]YANG B,JIANG Y,RAHMAN M H,etal.Identification and expression analysis of WRKY transcription factor genes in canola(BrassicanapusL.) in response to fungal pathogens and hormone treatments[J].BMCPlantBiology,2009,9(1):68.
[4]LING J,JIANG W,ZHANG Y,etal.Genome-wide analysis of WRKY gene family inCucumissativus[J].BMCGenomics,2011,12(1):471.
[5]XIONG W,XU X,ZHANG L,etal.Genome-wide analysis of the WRKY gene family in physic nut(JatrophacurcasL.)[J].Gene,2013,524(2):124-132.
[6]LI H L,ZHANG L B,GUO D,etal.Identification and expression profiles of the WRKY transcription factor family inRicinuscommunis[J].Gene,2012,503(2):248-253.
[7]DONG J,CHEN C,CHEN Z.Expression profiles of theArabidopsisWRKY gene superfamily during plant defense response[J].PlantMolecularBiology,2003,51(1):21-37.
[8]HUAG X,LI K,XU X,etal.Genome-wide analysis of WRKY transcription factors in white pear(Pyrusbretschneideri) reveals evolution and patterns under drought stress[J].BMCGenomics,2015,16(1):1104.
[9]HE H,DONG Q,SHAO Y,etal.Genome-wide survey and characterization of the WRKY gene family inPopulustrichocarpa[J].PlantCellReports,2012,31(7):1199-1217.
[10]MUTHAMILARASAN M,BONTHALA V S,KHANDELWAL R,etal.Global analysis of WRKY transcription factor superfamily inSetariaidentifies potential candidates involved in abiotic stress signaling[J].FrontiersinPlantScience,2015,6:910.
[11]ROSS C A,LIU Y,SHEN Q J.The WRKY gene family in rice(Oryzasativa)[J].JournalofIntegrativePlantBiology,2007,49(6):827-842.
[12]CAI C,NIU E,DU H,etal.Genome-wide analysis of the WRKY transcription factor gene family inGossypiumraimondiiand the expression of orthologs in cultivated tetraploid cotton[J].TheCropJournal,2014,2(2):87-101.
[13]WEI K F,CHEN J,CHEN Y F,etal.Molecular phylogenetic and expression analysis of the complete WRKY transcription factor family in maize[J].DNAResearch,2012:dsr048.
[14]YIN G,XU H,XIAO S,etal.The large soybean(Glycinemax) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups[J].BMCPlantBiology,2013,13(1):148.
[15]EULGEM T,RUSHTON P J,ROBATZEK S,etal.The WRKY superfamily of plant transcription factors[J].TrendsinPlantScience,2000,5(5):199-206.
[16]PANDEY S P,SOMSSICH I E.The role of WRKY transcription factors in plant immunity[J].PlantPhysiology,2009,150(4):1648-1655.
[17]ZHENG Z,MOSHER S L,FAN B ,etal.Functional analysis ofArabidopsisWRKY25 transcription factor in plant defense againstPseudomonassyringae[J].BMCPlantBiology,2007,7(1):2.
[18]ZHENG Z,QAMAR S A,CHEN Z,etal.ArabidopsisWRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens[J].ThePlantJournal,2006,48(4):592-605.
[19]LI J,BRADER G,PALVA E T.The WRKY70 transcription factor:a node of convergence for jasmonate-mediated and salicylate-mediated signals in plant defense[J].ThePlantCell,2004,16(2):319-331.
[20]QIU Y,YU D.Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance inArabidopsis[J].EnvironmentalandExperimentalBotany,2009,65(1):35-47.
[21]WANG H,HAO J,CHEN X,etal.Overexpression of rice WRKY89 enhances ultraviolet B tolerance and disease resistance in rice plants[J].PlantMolecularBiology,2007,65(6):799-815.
[22]YUE H,WANG M,LIU S,etal.Transcriptome-wide identification and expression profiles of the WRKY transcription factor family in Broomcorn millet(PanicummiliaceumL.)[J].BMCGenomics,2016,17(1):343.
[23]齊濤.中國玉米國際競爭力研究[D].陜西楊凌:西北農(nóng)林科技大學,2011.
QI T.Study on the intrnational competitiveness of corn in china [D].Yangling Shaanxi:Northwest A&F University,2011.
[24]劉巍.中國玉米及其深加工產(chǎn)品國際競爭力研究[D].北京:對外經(jīng)濟貿(mào)易大學,2009.
LIU W.Study on international competitiveness of Chinese corn and its deep processing products [D].Beijing:University of International Business and Economics,2009.
[25]顏君,郭興啟,曹學成.WRKY轉錄因子的基因組水平研究現(xiàn)狀[J].生物技術通報,2015,31(11):9-17.
YAN J,GUO X Q,CAO X CH.Progress of genome-wide researches on WRKY transcription factors[J].BiotechnologyBulletin,2015,31(11):9-17.
[26]石紅梅,戰(zhàn)新梅,管世銘,等.牡丹PsWRKY基因的克隆和表達特性分析[J].植物生理學報,2015,51(10):1743-1748.
SHI H M,ZHAN X M,GAN SH M,etal.Clong and expression analysis ofPsWRKYin tree peony(Paeoniasuffruticosa)[J].PlantPhysiologyJournal,2015,51(10):1743-1748.
[27]向小華,吳新儒,晁江濤,等.普通煙草WRKY基因家族的鑒定及表達分析[J].遺 傳,2016,38(9):840-856.
XIANG X H,WU X R,CHAO J T,etal.Genome-wide identification and expression analysis of the WRKY gene family in common tobacco(NicotianatabacumL.)[J].Hereditas,2016,38(9):840-856.
[28]HUANG D,WU W,ABRAMS S R,etal.The relationship of drought-related gene expression inArabidopsisthalianato hormonal and environmental factors[J].JournalofExperimentalBotany,2008,59(11):2991-3007.
[29]JIANG Y,DEYHOLOS M K.Functional characterization ofArabidopsisNaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses[J].PlantMolecularBiology,2009,69(1/2):91-105.
[30]LI S,FU Q,CHEN L,etal.ArabidopsisthalianaWRKY25,WRKY26 and WRKY33 coordinate induction of plant thermo tolerance[J].Planta,2011,233(6):1237-1252.
[31]QIU Y,YU D.Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance inArabidopsis[J].EnvironmentalandExperimentalBotany,2009,65(1):35-47.
[32]ZHOU Q Y,TIAN A G,ZOU H F,etal.Soybean WRKY-type transcription factor genes,GmWRKY13, GmWRKY21 and GmWRKY54,confer differential tolerance to abiotic stresses in transgenicArabidopsisplants[J].PlantBiotechnologyJournal,2008,6(5):486-503.
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
