東北森林區(qū)與草原區(qū)草本植物群落植硅體組合特征的對比研究
2018-03-26 09:14:11夏瓔凡介冬梅李德暉周沛芳宋麗娜蒙萌蔣雪純
草業(yè)學報 2018年3期
關鍵詞:植物
夏瓔凡,介冬梅,2,3*,李德暉,2,3,周沛芳,宋麗娜,蒙萌,蔣雪純
(1.東北師范大學地理科學學院,吉林 長春 130024;2.國家環(huán)境保護濕地生態(tài)與植被恢復重點實驗室,吉林 長春 130024;3.東北師范大學草地研究所植被生態(tài)科學教育部重點實驗室,吉林 長春 130024)
目前,實現(xiàn)植被類型的判別有多種方式。其中,歸一化植被指數(shù)(normalized difference vegetation index, NDVI)已成為較為通用的、可用于大尺度區(qū)域的植被指數(shù)監(jiān)測方法。但是,這種方法僅適用于宏觀、短時間尺度的植被動態(tài)研究,并不能有效地應用于長時間尺度下森林和草原的判別,無法據(jù)此有效地恢復古植被、古環(huán)境[1]。孢粉分析通過提取地層中的花粉,可以一對一重建過去的植物種類,在古植被、古環(huán)境重建上有一定效果。但是,在以禾本科植物為優(yōu)勢的草原群落,由于草原群落孢粉組合中占優(yōu)勢的蒿屬(Artemisia)和藜科(Chenopodiaceae)等花粉常表現(xiàn)為超代表性,極有可能掩蓋了草原植被真實信息,而且孢粉分析對植物種類的鑒定也只能達到科的水平,亞科之間難以區(qū)分[2]。一般而言,植硅體對草本植物群落中的植物種類可以鑒定到亞科,利用個別特殊形態(tài)可以鑒定到屬[3-4],結合形態(tài)參數(shù)甚至可以達到種[5],這樣的判別能力對科學、準確地判別植物種類,進而重建禾本科為主的區(qū)域古植被、古環(huán)境尤為重要。
植物硅酸體, 簡稱植硅體, 是指植物體內(nèi)的硅通過根系從土壤中吸收, 經(jīng)維管束傳輸, 在細胞內(nèi)腔或細胞之間以水合硅(SiO2·nH2O)的形式出現(xiàn), 在植物體內(nèi)形成難溶的硅酸形態(tài)[6]。在植硅體首次被發(fā)現(xiàn)以來的 150 多年里,學者們對現(xiàn)代幾百種植物植硅體的形態(tài)進行了大量研究,發(fā)現(xiàn)不同植物種產(chǎn)生的植硅體形態(tài)及形態(tài)組合不同[7-12],明確了不同草本植物植硅體的形態(tài)存在一定差異。
草本植物群落所包含的植物種類、各植物種的數(shù)量不盡相同,因此植物群落產(chǎn)生的植硅體組合也會存在差異。著眼于此,本研究針對現(xiàn)代森林區(qū)和草原區(qū)草本植物群落產(chǎn)生的植硅體組合特征,尋找兩者之間的差異,以期為森林區(qū)和草原區(qū)草本植物群落的判別提供一定參考依據(jù)。這一工作不僅可以補充和豐富植硅體形態(tài)學研究內(nèi)容,還可以為禾本科群落分布廣泛的區(qū)域古植被的高精度重建提供科學依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
中國東北地區(qū)位于亞歐大陸東岸,其北界與東界均為國界,西界為大興安嶺,南界為遼寧省南部渤海海岸線。該區(qū)為大陸性季風氣候, 四季分明, 夏季短促而溫暖多雨, 冬季漫長而寒冷, 冬夏之間季風交替。區(qū)域東部與南部分別與日本海和渤海、黃海毗鄰, 受海洋性氣候影響, 水分充足; 北部受西伯利亞寒潮影響, 西部受內(nèi)蒙古冷高壓影響氣溫很低, 具有顯著的冷濕氣候特征[13]。
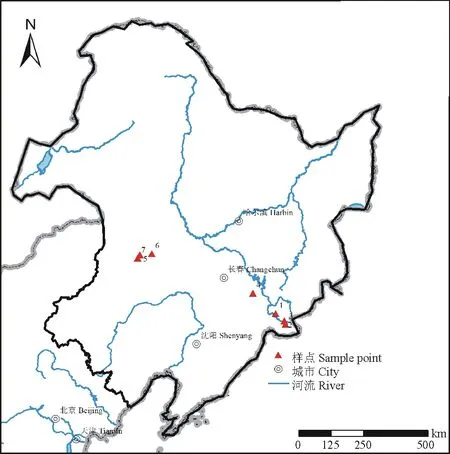
圖1 樣點分布圖Fig.1 Distribution of sample points
年平均氣溫為-4~11 ℃, 7月平均氣溫21~26 ℃, 1月平均氣溫-24~9 ℃, 年降水量350~1100 mm, 其中50%以上的降水發(fā)生在7-9月,從東南向西北有一定的內(nèi)部差異[14]。受距海遠近影響, 由東到西(1400 km)可依次劃分為濕潤、半濕潤和半干旱氣候區(qū), 在大陸尺度上呈現(xiàn)出了森林、森林草原、草甸草原的依次更替,對應的地帶性土壤的更替序列為暗棕壤、黑土、黑鈣土。這為在同一個相對獨立的自然地理區(qū)域內(nèi),對森林區(qū)和草原區(qū)草本植物群落植硅體的形態(tài)及組合特征進行對比分析提供了優(yōu)勢條件。
1.2 采樣點設計
本研究采樣點位于吉林省東部長白山地、內(nèi)蒙古東部大興安嶺東麓(圖1),采樣點位置信息(經(jīng)度、緯度、海拔)記錄于表 1。采樣方法為:在森林區(qū)內(nèi),先在植物原生性較好的森林群落內(nèi),劃出一個 10 m×10 m 的區(qū)域,再在該區(qū)域的四角以及中間,共采集5個 1 m×1 m 草本植物樣方,并按照采集順序依次編號[如,興隆1(XL1)];在草原區(qū)內(nèi),每個樣點采集2個 1 m×1 m 草本植物樣方,并按照采集順序依次編號[如,代欽塔拉1(DQTL1)]。收割樣方內(nèi)所有草本植物,同時記錄樣方信息,包括優(yōu)勢種、不同植物種株數(shù)等,并稱重、裝袋以及編號。本研究共采集了26個草本植物樣方,其中森林區(qū)20個 1 m×1 m 樣方,草原區(qū) 6個 1 m×1 m 樣方。
森林區(qū)主要草本植物多為雙子葉植物,如歪頭菜(Viciaunijuga)、蕁麻葉龍頭草(Meehaniaurticifolia)、銀線草(Chloranthusjaponicus)、白花碎米薺(Cardamineleucantha)、拉拉藤屬(Galium)等;單子葉植物主要為薹草屬(Carex)等,偶見禾本科(Gramineae),如野青茅(Deyeuxiaarundinacea);蕨類植物有木賊(Equisetumhyemale)等。草原區(qū)雙子葉植物中多為蒿屬(Artemisia),此外還有豬毛菜(Salsolacollina)、野艾蒿(Artemisialavandulifolia)、委陵菜(Potentillachinensis)、野大麻(Cannabissativa)、柴胡屬(Bupleurum)、尖頭葉藜(Chenopodiumacuminatum)等;單子葉植物在草原區(qū)主要以禾本科為主,如狗尾草(Setariaviridis)、羊草(Leymuschinensis)、牛鞭草(Hemarthriasibirica)、燕麥芨芨草(Achnatherumavenoide)、蘆葦(Phragmitesaustralis)等,偶見莎草科(Cyperaceae);蕨類植物中僅見木賊的一個變種——興安木賊(Equisetumvariegatum)。
1.3 實驗方法
本實驗于2016年進行,選擇同一生長期內(nèi)森林區(qū)和草原區(qū)的草本植物樣品,以群落為單位,全株處理樣品。對草本植物群落樣品進行植硅體分析, 共分析草本植物群落樣品26個。
實驗采用濕式灰化法[9,15-16]。實驗步驟如下:
1) 樣本前處理。用超聲波清洗儀將樣方內(nèi)植物反復清洗, 烘干,剪成小段,粉碎成末,并稱重群落粉末樣品1 g,放入干凈的離心管中。
2) 氧化。在離心管中加入濃硝酸, 待有機質在沸水浴環(huán)境中全部被氧化后, 向試管中加入蒸餾水, 用離心機(2500 r·min-1)離心清洗10 min·次-1,并反復離心至少3次,直至溶液接近酸堿中性。
3) 加入孢子片。向試管中加入石松孢子片以及濃鹽酸, 待反應充分后,再次向試管中加入蒸餾水, 用離心機(2500 r·min-1)離心清洗10 min·次-1,并反復離心至少3次,直至溶液接近酸堿中性。
4) 固定制片。加入無水乙醇,用離心機(2500 r·min-1)離心清洗10 min·次-1,并反復離心1~2次,緩慢傾倒上層清液,保留少量液體。將試管中的液體振蕩均勻,用一次性滴管吸取2~3滴,滴在載玻片上,用酒精燈加熱,待無水乙醇蒸發(fā)后,滴上1~2滴加拿大樹膠,蓋上蓋玻片,制成固定片。
5) 鑒定與統(tǒng)計。采用 MOTIC 生物顯微鏡鑒定統(tǒng)計樣品, 每個樣品統(tǒng)計植硅體不少于300粒,共統(tǒng)計10170粒植硅體。本研究提取的植硅體樣品和制作的玻片保存在東北師范大學地理科學學院實驗室。
1.4 數(shù)據(jù)分析
數(shù)據(jù)分析采用定性與定量相結合的方法。首先根據(jù)森林區(qū)與草原區(qū)草本植物植硅體的形態(tài)特征,將植硅體類型分為15種。而后,利用SPSS 19.0軟件對森林區(qū)與草原區(qū)草本植物群落相同類型植硅體的數(shù)量進行對比,從方差分析和聚類分析兩個角度,明確了森林區(qū)與草原區(qū)相同類型草本植物群落植硅體數(shù)量的相似程度與差異程度。再后,利用前人建立的Ic指數(shù)[17]對森林區(qū)與草原區(qū)草本植物群落植硅體指數(shù)進行對比,明確了森林區(qū)與草原區(qū)草本植物群落的不同環(huán)境特征。
2 結果與分析
通過對26個草本植物群落中的10170粒植硅體進行鑒定、統(tǒng)計,可將其分為15種植硅體類型(不包括硅化組織)。其中,各個草本植物群落植硅體的組合類型不盡相同,相同類型植硅體的數(shù)量也存在差異。
2.1 森林區(qū)與草原區(qū)草本植物植硅體組合類型對比
就細胞起源而言:啞鈴型、鞍型、齒型、帽型為短細胞植硅體;棒型為長細胞植硅體;毛發(fā)狀和尖型為毛狀細胞植硅體;邊緣彎曲扁平狀和硅化表皮細胞屬于表皮細胞植硅體;還有硅化氣孔(器)、硅化導管、硅質突起、扇型、塊狀等形態(tài)。此外,還見到了一些尚未在文獻中定名描述的植硅體,本研究統(tǒng)稱其為不規(guī)則型。
1) 短細胞植硅體。森林區(qū)和草原區(qū)都產(chǎn)生啞鈴型植硅體(圖2a~e),森林區(qū)在亞類上多為A型鈴(圖2a),而草原區(qū)在亞類上多為C型啞鈴型(圖2b)和兩個半鈴型(圖2c);此外,只在草原區(qū)發(fā)現(xiàn)了十字型植硅體(圖2d)。鞍型植硅體在森林區(qū)和草原區(qū)均有出現(xiàn)(圖2f~g)。森林區(qū)和草原區(qū)均產(chǎn)生齒型植硅體(圖2h~i),但從總體來看,森林區(qū)產(chǎn)量略高于草原區(qū),且多為普通齒型(圖2h)。帽型植硅體(圖2j~k)在森林區(qū)的含量幾乎為0,而在草原區(qū)有一定產(chǎn)出,且主要表現(xiàn)為平頂帽型(圖2j)和尖頂帽型(圖2k)。從總體來看,森林區(qū)的短細胞植硅體產(chǎn)量遠低于草原區(qū)。
2) 長細胞植硅體(棒型植硅體)。森林區(qū)和草原區(qū)均可產(chǎn)生棒型植硅體(圖2l~u)。其中,平滑棒型(圖2l)和扁平棒型(圖2m)較為多見,森林區(qū)的產(chǎn)量高于草原區(qū)。牛角棒型(圖2n)和樹突棒型(圖2o),在森林區(qū)和草原區(qū)都產(chǎn)生得較少,幾乎為0。草原區(qū)多產(chǎn)生較典型的刺狀棒型(圖2p)、刺狀突起棒型(圖2q)。此外,還可觀察到一些其他類型的棒型植硅體,如來源于含有木賊的群落的特殊刺狀突起棒型(圖2r);僅產(chǎn)生于森林區(qū)的三棱柱型(圖2s);可彎曲的棒型(圖2t)等。從總體來看,森林區(qū)的長細胞植硅體產(chǎn)量高于草原區(qū)。
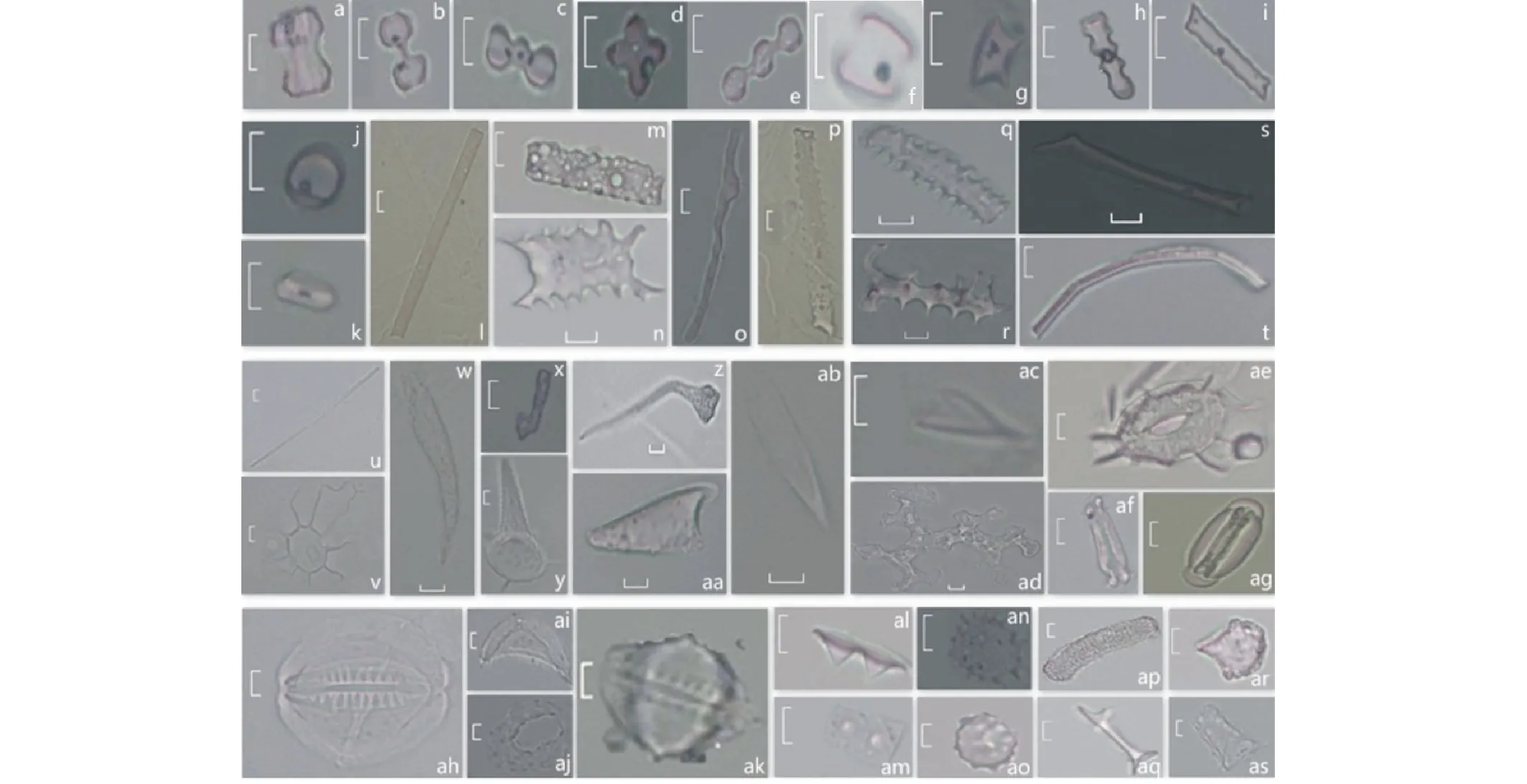
圖2 東北森林區(qū)與草原區(qū)草本植物植硅體形態(tài)Fig.2 The type of herbaceous phytolith in forest region and grassland region in Northeast China a~e:啞鈴型Bilobate;f~g:鞍型Saddle;h~i:齒型Trapeziform;j~k:帽型Rondel;l~r,t~u:棒型Elongate;s:有轉折的三棱柱型Triangular turn;v:毛發(fā)底座Hair base;w~z:毛發(fā)狀Hair;aa~ac:尖型Lanceolate;ad:有穴狀紋飾的邊緣彎曲扁平狀Tabular stellate scrobiculate;ae~ak:硅化氣孔Silicified stomata and silicified stomatal apparatus;al~an:硅質突起Papillate;ao:表面硅質突起的球型Globular papillate;ap:絮狀長條型Oblong reticulate;aq:不規(guī)則型Uncertain type;ar:扇型Cuneiform bulliform cell;as:塊狀Parallepipedal bulliform cell (標尺均為10 μm Scalar is 10 μm).
3) 毛狀細胞植硅體。森林區(qū)主要產(chǎn)生毛發(fā)狀植硅體(圖2v~z)和尖型植硅體(圖2aa~ac),而在草原區(qū)多產(chǎn)生尖型植硅體,也產(chǎn)生了一些毛發(fā)狀植硅體,其中還有毛發(fā)狀植硅體中較為特殊的小毛發(fā)狀植硅體(圖2x)。從總體來看,森林區(qū)的毛狀細胞植硅體產(chǎn)量低于草原區(qū)。
4) 表皮細胞植硅體。森林區(qū)和草原區(qū)都產(chǎn)生表皮細胞植硅體,多為邊緣彎曲扁平狀(圖2ad),還可觀察到一些硅化不完全的表皮細胞。從總體來看,森林區(qū)的表皮細胞植硅體產(chǎn)量略高于草原區(qū),但產(chǎn)量都不算高。
5) 硅化氣孔(器)。森林區(qū)產(chǎn)生的硅化氣孔(圖2ae~ak)形態(tài)較為豐富,涵蓋了蕨類植物、單子葉植物、雙子葉植物的氣孔類型。其中,木賊科植物的硅化氣孔(器)形態(tài)特殊但容易破碎,因此經(jīng)常見到的是木賊科植物氣孔器的各個構件,包括硅化氣孔(圖2ah)、硅化保衛(wèi)細胞(圖2ai)、硅化氣孔窩(圖2aj),完整的硅化氣孔器偶爾可以見到。草原區(qū)也存在木賊的某一變種產(chǎn)生的完整硅化氣孔器(圖2ak)。從總體來看,森林區(qū)的硅化氣孔(器)產(chǎn)量高于草原區(qū)。
6) 硅質突起。硅質突起(圖2al~an)多來源于莎草科植物,是森林區(qū)的優(yōu)勢植硅體類型(本研究定義植硅體組合中百分含量最大的植硅體類型為優(yōu)勢類型)。硅質突起的形態(tài)豐富,包括錐狀(圖2al)、珠狀(圖2am)等。此外,在突起數(shù)量、有無紋飾上也存在差異。草原區(qū)多產(chǎn)生刺狀硅質突起(圖2an)。從總體來看,森林區(qū)的硅質突起產(chǎn)量遠高于草原區(qū)。
7) 其他型。在研究中還發(fā)現(xiàn)了許多特殊形態(tài)植硅體,如“表面硅質突起的球型”(圖2ao)、“絮狀長條型”(圖2ap),還發(fā)現(xiàn)了一些尚未被定名的植硅體形態(tài),稱之為“不規(guī)則型”(圖2aq)。此外,還有一些數(shù)量較少的植硅體,如扇型(圖2ar)、塊狀(圖2as)等。以上形態(tài)均被劃到其他型中。
對以上描述過的植硅體形態(tài)作總結,發(fā)現(xiàn)森林區(qū)和草原區(qū)草本植物產(chǎn)生的植硅體類型中,在形態(tài)、數(shù)量、紋飾等方面存在一定差異(表2),可以認為這樣的植硅體具有植被指示意義。
2.2 森林區(qū)與草原區(qū)草本植物群落相同類型植硅體數(shù)量對比
通過對植硅體類型進行單因素方差分析(表3),其中毛發(fā)狀植硅體(P=0.173)、尖型植硅體(P=0.509)、硅化導管(P=0.852)、邊緣彎曲扁平狀(P=0.444)和硅化氣孔(P=0.204)未通過顯著性檢驗,但硅質突起(P=0.015)和硅化組織(P=0.024)的平均百分含量在森林區(qū)和草原區(qū)有顯著差異,短細胞有極顯著差異(P=0.000)。可以認為,在森林區(qū)與草原區(qū)的草本植物群落中,毛狀細胞植硅體、尖型植硅體、硅化導管和硅化氣孔的平均百分含量相差不大,但硅化組織、硅質突起平均百分含量存在顯著差別,森林區(qū)高于草原區(qū),而短細胞植硅體平均百分含量存在極大差別,草原區(qū)遠遠高于森林區(qū)。

表2 森林區(qū)與草原區(qū)草本植物具有指示意義的植硅體類型Table 2 The phytolith of herbaceous plant with indicative significance in forest region and grassland region
注:*表示“有”,—表示“無”,*的數(shù)量表示兩區(qū)內(nèi)同種植硅體類型平均百分含量的比值,如,4*表示“森林區(qū)齒型平均百分含量是草原區(qū)的4倍”。
Note:* shows “Have”, — shows “No”. The number of “*” shows the ratio of percentage content of the same phytolith in two regions.Eg, “4*” shows that “the percentage content of trapeziform in forest region is four times than grassland region.”
通過單因素方差分析可知,利用森林區(qū)與草原區(qū)草本植物群落中平均百分含量明顯不同的硅化組織、硅質突起和短細胞植硅體可將兩區(qū)的群落區(qū)別開來,但是對于在森林區(qū)與草原區(qū)平均百分含量相差不大的共有植硅體類型,僅通過數(shù)量進行描述,并不能明確區(qū)分森林和草原,因此可以采用形態(tài)參數(shù)、分形維數(shù)、不變矩等方法,進一步系統(tǒng)地、量化地描述草本植物群落中的植硅體。
進一步對植硅體組合類型進行聚類分析,26個群落被分為森林區(qū)群落和草原區(qū)群落兩個集合,說明森林區(qū)和草原區(qū)兩類草本植物群落植硅體組合之間存在一定差異性,但森林區(qū)的20個草本植物群落植硅體組合具有一定相似性,草原區(qū)6個草本植物群落植硅體組合具有一定相似性。為了進一步研究兩區(qū)內(nèi)部群落的相似性,又可將森林區(qū)草本植物群落分為4組,草原區(qū)草本植物群落分為2組。
從兩大區(qū)植硅體組合的平均百分含量來看(表4),森林區(qū)草本植物群落植硅體組合中,硅質突起、硅化氣孔、齒型、啞鈴型、其他棒型、尖型、平滑棒型、刺棒型、扁棒型的累積百分含量達到90.46%,硅質突起為優(yōu)勢植硅體類型(43.78%)。此外,還有少量鞍型、毛發(fā)狀、硅化組織、帽型、硅化導管、邊緣彎曲扁平狀、牛角棒型、樹突棒型和其他類型植硅體。草原區(qū)草本植物群落植硅體組合中,啞鈴型、帽型、硅質突起、鞍型、尖型、硅化氣孔、齒型、刺棒型的累積百分含量達到90.51%,其中,啞鈴型為優(yōu)勢植硅體類型(54.68%)。此外,還有少量毛發(fā)狀、平滑棒型、硅化導管、扁平棒型和其他類型植硅體。
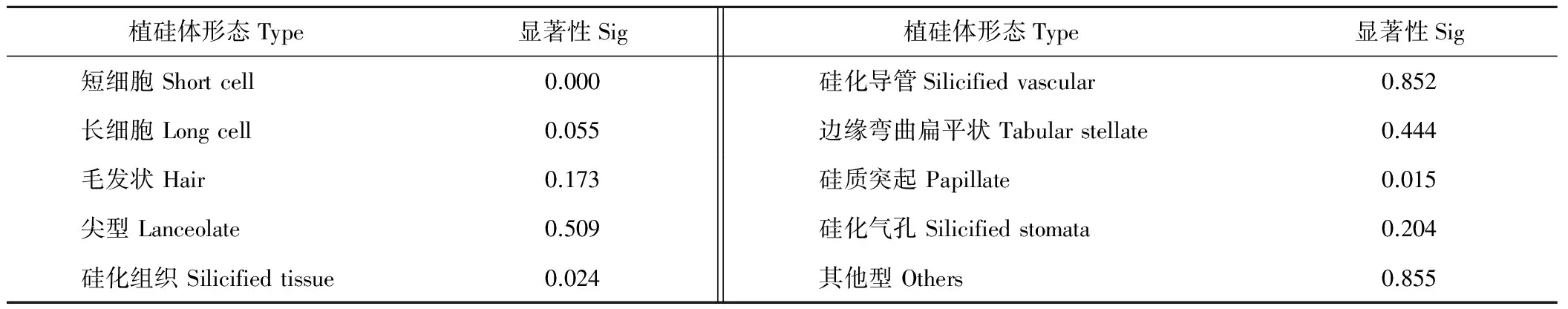
表3 植硅體形態(tài)單因素方差分析Table 3 The One-Way ANOVA of type
通過將森林區(qū)與草原區(qū)草本植物群落的植硅體組合對比可以發(fā)現(xiàn)(表4),森林區(qū)的優(yōu)勢植硅體類型——硅質突起,約為草原區(qū)的5.96倍。硅化氣孔形態(tài)豐富,且含量較高(15.52%),約為草原區(qū)的4.93倍。長細胞植硅體約為草原區(qū)的2.5倍。森林區(qū)的短細胞植硅體含量較低(17.6%),只有草原區(qū)的1/4。森林區(qū)毛狀細胞植硅體含量低于草原區(qū),約為后者的3/5,這是因為草原區(qū)草本植物群落中以禾本科植物為主,而禾本科植物能夠產(chǎn)生大量尖型植硅體,因此草原區(qū)尖型植硅體平均百分含量(5.92%)遠高于森林區(qū)(3.94%)。表皮細胞植硅體在研究中發(fā)現(xiàn)多為邊緣彎曲扁平狀和硅化表皮細胞,在森林區(qū)有少量可見,在草原區(qū)并未發(fā)現(xiàn)(圖3和圖4)。
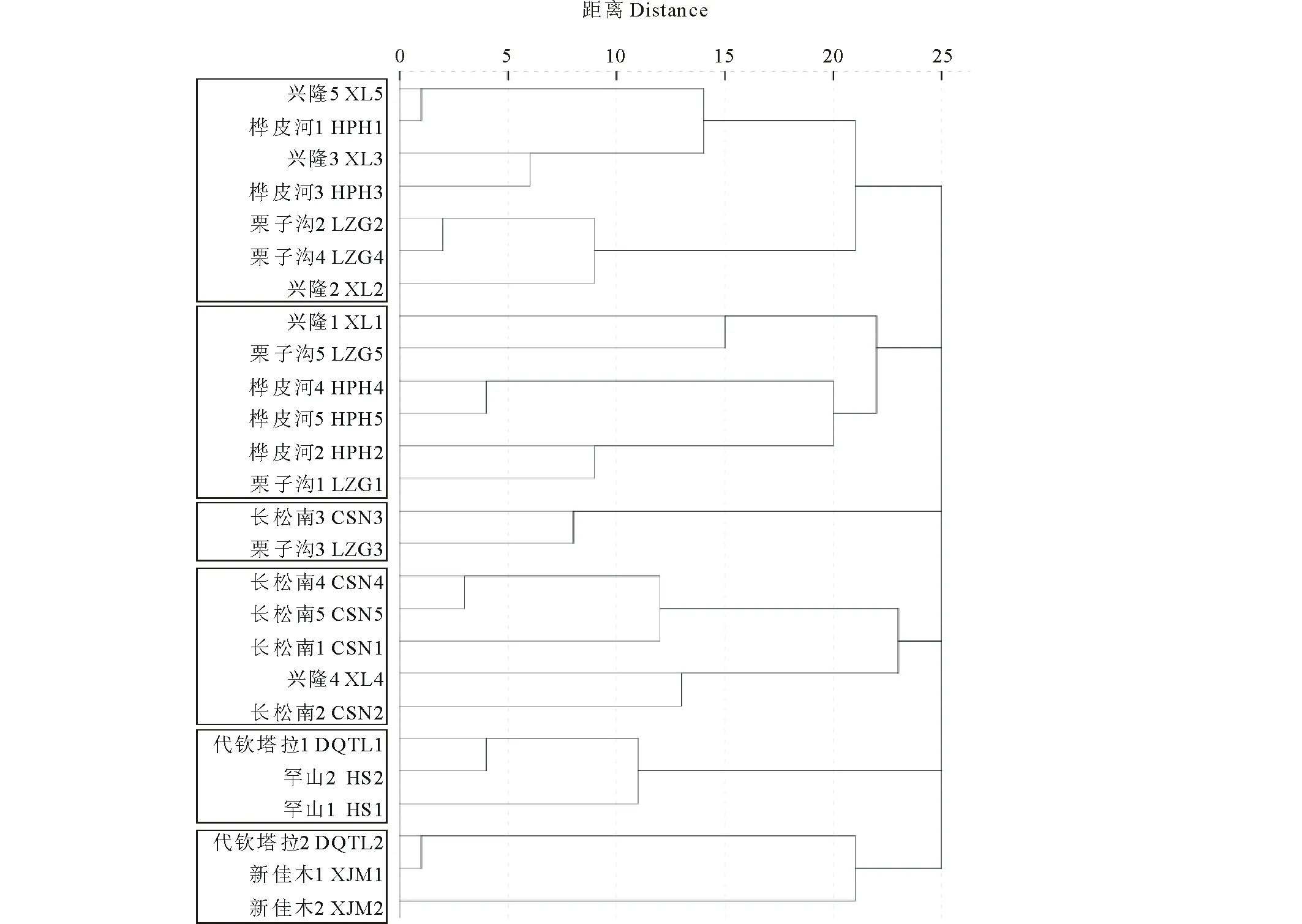
圖3 草本植物群落植硅體組合聚類分析Fig.3 The clustering analysis of herbaceous phytolith assemblages 樣點名稱后的數(shù)字代表該樣方在其樣點中的編號,如,“興隆5”代表“在興隆樣點中的第5個樣方”。The number after sample point represents the number of quadrat in its sample point. Eg, “XL5” means “the fifth quadrat in Xinglong.”
個別植硅體形態(tài)所占百分含量的變化范圍較大(表 4),這與地上生長的植物種類直接相關。森林區(qū)的硅質突起和硅化氣孔波動較大是由“長松南”樣點的特殊性造成的,該樣點生長著干物質重量大且含硅量高的木賊,其主要植硅體類型為硅化氣孔(器),因此導致森林區(qū)硅化氣孔的最大值出現(xiàn)異常。草原區(qū)的短細胞植硅體百分含量的波動較大,這是群落內(nèi)植物種類的差異導致的。當?shù)厣现参锒酁樘J竹亞科(Arundinoideae)時,鞍型百分含量大;當早熟禾亞科(Pooideae)植物占優(yōu)勢時,帽型、齒型的百分含量就高。在東北地區(qū)禾本科植物中,各亞科的生境不盡相同,同一樣點內(nèi),各亞科的種類、數(shù)量不可能“勢均力敵”,因此各型植硅體的百分含量變幅很大。
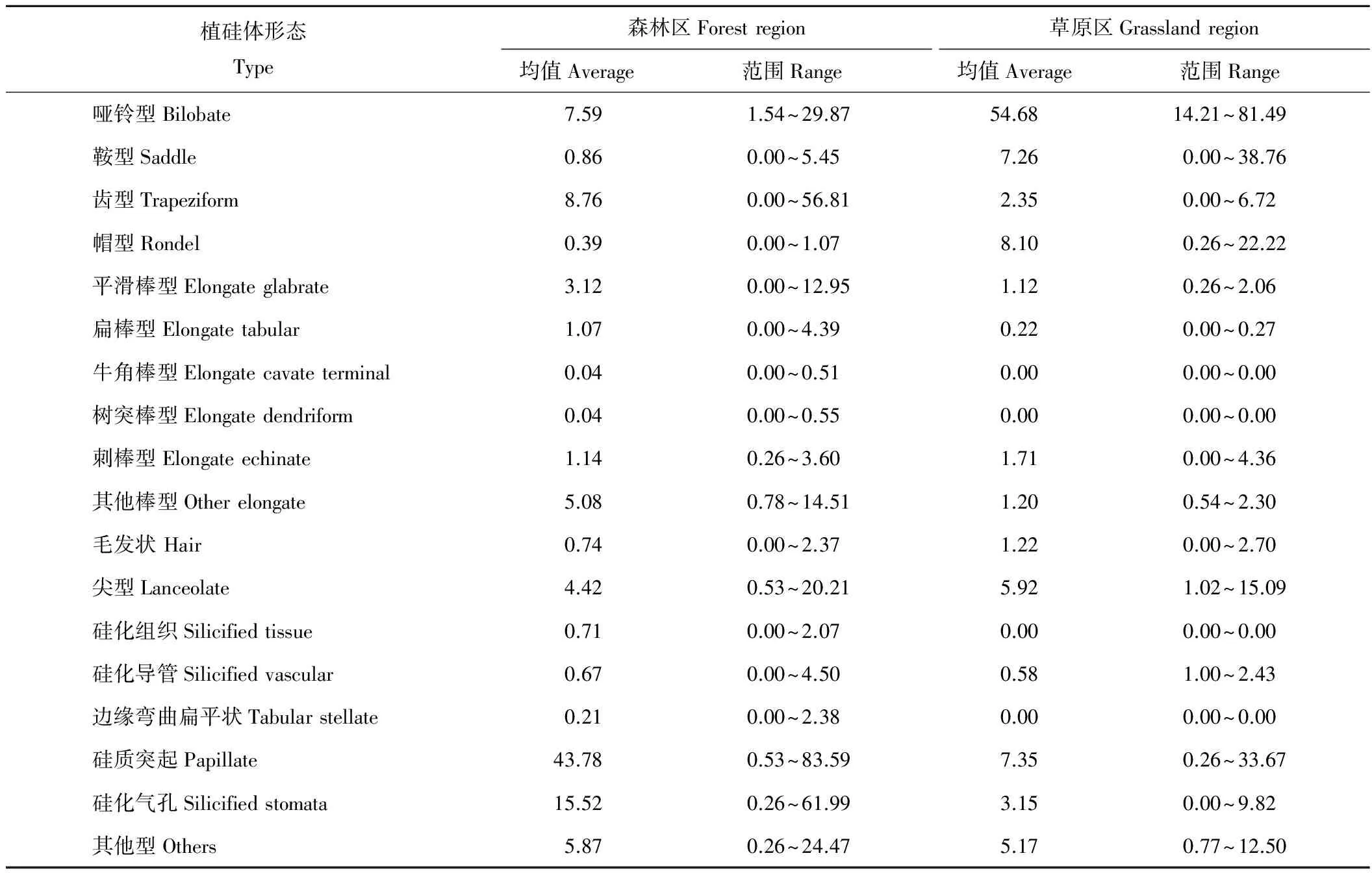
表4 森林區(qū)與草原區(qū)草本植物群落植硅體組合(相對百分含量) Table 4 The relative percentage content of herbaceous phytolith assemblages in forest region and grassland region (%)
2.2.1森林區(qū)草本植物群落植硅體組合 根據(jù)聚類分析結果(圖3和圖4),森林區(qū)的20個草本植物群落可劃分為4組。
第1組:興隆5、樺皮河1、興隆3、樺皮河3、栗子溝2、栗子溝4、興隆2。硅質突起的平均百分含量最大(74.52%),這是由于以上7個群落環(huán)境較濕潤,多生長莎草科薹草屬植物。短細胞中啞鈴型的百分含量差異不大,但在4組中的平均百分含量并不算高(5.44%)。
第2組:興隆1、栗子溝5、樺皮河4、樺皮河5、樺皮河2、栗子溝1。在這些群落中,短細胞的平均百分含量(39.18%)與其他類型植硅體相比居于首位,其中齒型植硅體的平均百分含量(27.50%)最高。另外,本組硅質突起的平均百分含量(30.58%)與其他類型植硅體相比排位靠前,但不是優(yōu)勢植硅體類型。長細胞植硅體的平均百分含量也比較高,為4組中的最高值(17.16%)。
第3組:長松南3、栗子溝3。這兩個群落除了硅質突起和硅化氣孔以外,其余類型植硅體的平均百分含量遠低于森林區(qū)其他3組。另外,長松南3的硅化氣孔含量遠遠高于栗子溝3,栗子溝3的硅質突起含量遠遠高于長松南3。這是由于長松南3處木賊較多,栗子溝3處莎草科薹草屬植物較多。
第4組:長松南4、長松南5、長松南1、興隆4、長松南2。這組硅化氣孔的平均百分含量最高(43.83%),這是由于群落內(nèi)生長著干重大且硅含量高的木賊,其主要植硅體類型為硅化氣孔(器)。
2.2.2草原區(qū)草本植物群落植硅體組合 根據(jù)聚類分析結果(圖3和圖4),草原區(qū)的6個草本植物群落可劃分為2組。
第1組:代欽塔拉1、罕山2、罕山1。這3個群落的短細胞植硅體的平均百分含量相差不大(77.28%),但在兩組中居于首位。其中,啞鈴型植硅體的平均百分含量最高(73.72%),遠遠高于其他群落,這是由于本組群落以狗尾草為優(yōu)勢種。
第2組:代欽塔拉2、新佳木1、新佳木2。在所有群落中,這3個群落的短細胞植硅體平均百分含量低于上一組(67.50%),其中,啞鈴型的平均百分含量最高(35.64%),帽型次之(15.59%)。這是由于該組草本植物群落中有狗尾草和羊草存在。此外,新佳木長有屬于黍亞科的牛鞭草,能夠產(chǎn)生十字型植硅體。代欽塔拉2處硅質突起的百分含量達到了草原區(qū)的最大值(33.67%),這是由于該群落中生長著莎草科植物,從而導致硅質突起的百分含量較高。新佳木2處鞍型植硅體的百分含量是草原區(qū)最高的,其形態(tài)主要為普通鞍型,這是由于此處生長著蘆葦。
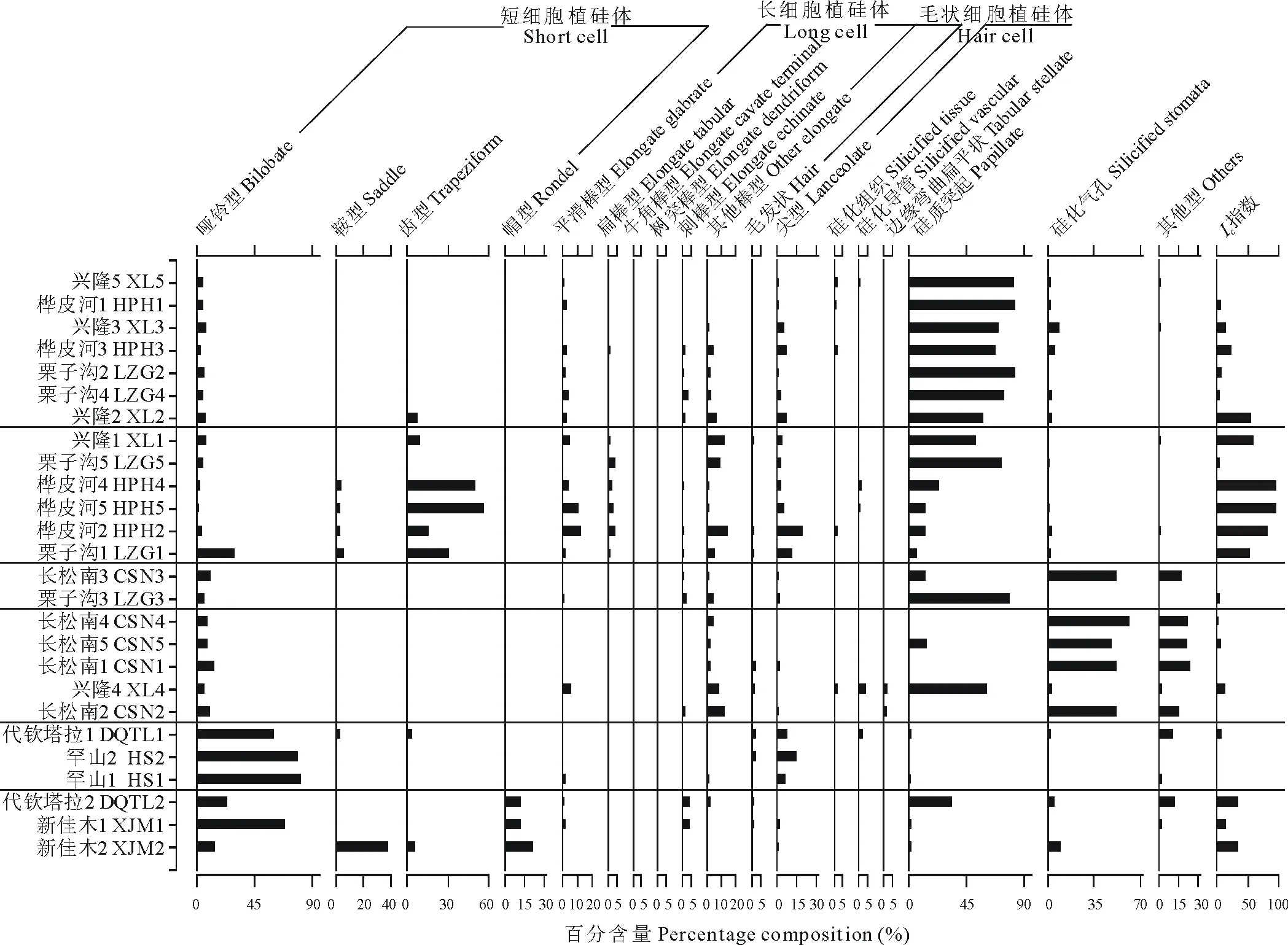
圖4 森林區(qū)與草原區(qū)草本植物群落植硅體組合及聚類結果Fig.4 The phytolith assemblages and clustering results of the 26 herbaceous plants in forest region and grassland region
2.3 東北森林區(qū)與草原區(qū)草本植物群落植硅體指數(shù)對比
利用Ic指數(shù) [Ic=早熟禾亞科植硅體/(早熟禾亞科植硅體+畫眉草亞科植硅體+黍亞科植硅體)×100%][17],對森林區(qū)與草原區(qū)草本植物群落植硅體指數(shù)進行對比。其中,早熟禾亞科植硅體的代表型為齒型和帽型植硅體,屬于C3型植物植硅體,指示寒冷環(huán)境;畫眉草亞科植硅體的代表型為短鞍型植硅體,黍亞科植硅體的代表型為十字型和啞鈴型植硅體,兩類代表型植硅體都屬于C4型植物植硅體,指示溫暖環(huán)境[6,17-20]。
根據(jù)表4結果,森林區(qū)內(nèi)產(chǎn)生的齒型和帽型植硅體平均百分含量之和達9.15%,是啞鈴型和鞍型植硅體含量的1.08倍。草原區(qū)內(nèi)產(chǎn)生的啞鈴型和鞍型植硅體平均百分含量之和達61.93%,是齒型和帽型植硅體含量的5.93倍。經(jīng)計算,總體看來,森林區(qū)草本植物群落Ic指數(shù)(Ic=27%)遠高于草原區(qū)草本植物群落(Ic=16%),這說明森林生境內(nèi)有更多早熟禾亞科植物生長,森林區(qū)草本植物群落的生長環(huán)境較草原區(qū)草本植物群落而言更加寒冷。
但在森林區(qū)與草原區(qū)中也存在著與Ic指數(shù)的總體趨勢不相同的草本植物群落。森林區(qū)草本植物群落中的長松南4、長松南5、長松南1、興隆4、長松南2這5個樣方的Ic指數(shù)均值較低(Ic=6%)是因為這5個群落中生長著木賊,其主要植硅體類型為硅化氣孔(器),使得早熟禾亞科代表型植硅體百分含量偏低,導致Ic指數(shù)較低。草原區(qū)草本植物群落中的代欽塔拉2、新佳木1、新佳木2這3個樣方的Ic指數(shù)均值較高(Ic=29%)是因為這3個群落中生長著羊草,產(chǎn)生了較多的帽型,使得早熟禾亞科代表型植硅體百分含量偏高,導致Ic指數(shù)較高。
3 討論
3.1 東北森林區(qū)與草原區(qū)草本植物植硅體組合類型
本研究共觀察到了15種規(guī)則的植硅體類型,包括鞍型、齒型、帽型、啞鈴型、棒型、毛發(fā)狀、尖型、硅質突起、塊狀、扇型、邊緣彎曲扁平狀、硅化氣孔、硅化導管、“表面硅質突起的球型”、“絮狀長條型”。其中,疣狀棒型、三棱柱型、硅化氣孔(木賊)以及邊緣彎曲扁平狀植硅體僅見于森林區(qū);十字型植硅體僅見于草原區(qū)。可以認為上述兩類形態(tài)的植硅體,分別為森林區(qū)草本植物植硅體和草原區(qū)草本植物植硅體的特征型。
啞鈴型在森林區(qū)和草原區(qū)均有分布,但森林區(qū)多為A型鈴,而相對干燥的草原區(qū)多為C型鈴。Lü等[21]曾對美國東南部和中國禾草產(chǎn)生的啞鈴型進行了研究,他們認為典型濕生草本植物產(chǎn)生的啞鈴型植硅體的柄較短,而旱生草本植物啞鈴型植硅體的鈴柄卻較長,本研究觀察到的結果與其是一致的。另外,本研究涉及的木賊科植物產(chǎn)生的硅化氣孔器特征鮮明,地層中木賊科氣孔的出現(xiàn)可能指示濕潤環(huán)境。值得注意的是,在過去的研究中,粗糙球型和刺球型植硅體一直被認為是棕櫚科植物的特征型[16],而光滑球型被認為可能來源于草本植物。但在研究過程中,在森林區(qū)長松南樣點的草本植物群落中卻發(fā)現(xiàn)了表面不甚光滑、帶有硅質突起的球型,其形態(tài)與在棕櫚科(Palmae)中發(fā)現(xiàn)的刺球型和褶球型[16]不盡相同。從硅化程度上看,本研究發(fā)現(xiàn)的球型植硅體硅化較淺;從大小上看,棕櫚科刺球型植硅體的球體大小在2.5~20.0 μm,本研究觀察到的表面硅質突起球型植硅體大小至少在10 μm 以上。此外,僅在森林區(qū)樺皮河樣點和栗子溝樣點出現(xiàn)的“絮狀長條型”植硅體也應當被注意,但其細胞來源尚未明確。
3.2 東北森林區(qū)與草原區(qū)草本植物群落相同類型植硅體的數(shù)量
森林區(qū)草本植物群落植硅體的平均百分含量為硅質突起(43.78%)>硅化氣孔(15.52%)>齒型(8.76%)>啞鈴型(7.59%)>其他棒型(5.08%)>尖型(4.42%)>平滑棒型(3.12%)>刺棒型(1.14%)>扁棒型(1.07%),這幾種形態(tài)的累積平均百分含量為90.46%。硅質突起為森林區(qū)草本植物群落的優(yōu)勢植硅體類型,可以指示濕潤環(huán)境。此外,植硅體組合中硅化氣孔的平均百分含量也較高。有研究表明[22-24],CO2濃度、溫度、水分等環(huán)境因子可以影響植物的蒸騰作用,從而引起植物氣孔的變化。本研究森林區(qū)草本植物群落植硅體組合中的硅化氣孔較多,可能是因為森林區(qū)降水較多,氣候濕潤,且樹冠高大,阻礙了草本植物接受太陽輻射,溫度較低,因此產(chǎn)生了較多的氣孔[23-24]。
草原區(qū)草本植物群落植硅體的平均百分含量為啞鈴型(54.68%)>帽型(8.10%)>硅質突起(7.35%)>鞍型(7.26%)>尖型(5.92%)>硅化氣孔(3.15%)>齒型(2.35%)>刺棒型(1.71%),這幾種形態(tài)的累積平均百分含量為90.51%。草原區(qū)植物群落多生長禾本科植物,其植硅體組合中短細胞的平均百分含量較高。黃翡等[25]曾對內(nèi)蒙古典型草原禾本科植硅體形態(tài)進行過研究,認為帽型、啞鈴型等短細胞植硅體在草原區(qū)禾本科植物中占絕對優(yōu)勢。本研究結果與前人一致。
3.3 東北森林區(qū)與草原區(qū)草本植物群落植硅體指數(shù)的指示意義
前人研究表明,植物體內(nèi)的植硅體組合特征可以記錄局地的植被面貌特征,植被面貌的變化主要受控于氣候和環(huán)境的變化[18]。為了更好地揭示植硅體組合特征對氣候和環(huán)境的指示性,Ic[17]、干旱指數(shù)Iph[26]等植硅體指數(shù)被用來反映植硅體組合對氣候或環(huán)境變化的響應[25-26]。
本研究采用Ic指數(shù)來指示森林區(qū)與草原區(qū)不同的環(huán)境特征。在本研究中,森林生境內(nèi)產(chǎn)生了較多指示寒冷環(huán)境的齒型和帽型植硅體,而草原生境內(nèi)則產(chǎn)生了較多指示溫暖環(huán)境的啞鈴型植硅體,這與前人研究結果一致[6,17-18,20,27]。經(jīng)計算,森林區(qū)草本植物群落Ic指數(shù)(Ic=27%)遠高于草原區(qū)草本植物群落(Ic=16%),這說明森林生境內(nèi)有更多早熟禾亞科植物生長,森林區(qū)草本植物群落的生長環(huán)境較草原區(qū)草本植物群落而言更加寒冷。
4 結論
從植硅體組合類型來看,東北森林區(qū)與草原區(qū)的草本植物可產(chǎn)生部分相同類型植硅體,但有一些類型的植硅體只產(chǎn)生于森林區(qū)或草原區(qū),如研究中發(fā)現(xiàn)的疣狀棒型、三棱柱型、硅化氣孔(木賊)以及邊緣彎曲扁平狀植硅體可作為東北森林區(qū)草本植物植硅體的特征型;十字型可作為東北草原區(qū)草本植物植硅體的特征型。另外,啞鈴型在森林區(qū)和草原區(qū)均有分布,但森林區(qū)多為A型鈴,鈴柄較短;而草原區(qū)多為C型鈴,鈴柄較長。在研究過程中,還在森林區(qū)發(fā)現(xiàn)了一些特殊形態(tài)植硅體——表面不甚光滑、帶有硅質突起、大小至少在 10 μm 以上的球型植硅體,以及一種細胞來源尚未明確的“絮狀長條型”植硅體。
從相同類型植硅體的數(shù)量來看,本研究區(qū)內(nèi)的森林區(qū)草本植物群落植硅體組合主要為硅質突起、硅化氣孔、齒型、啞鈴型、其他棒型、尖型、平滑棒型、刺棒型以及扁棒型,其中硅質突起為森林區(qū)草本植物群落的優(yōu)勢植硅體類型,可以指示濕潤環(huán)境。本研究區(qū)內(nèi)的草原區(qū)草本植物群落植硅體組合主要為啞鈴型、帽型、硅質突起、鞍型、尖型、硅化氣孔、齒型以及刺棒型,其中啞鈴型植硅體在草原區(qū)草本植物群落植硅體組合中占絕對優(yōu)勢。
本研究區(qū)內(nèi)的森林區(qū)草本植物群落產(chǎn)生了較多指示寒冷環(huán)境的齒型和帽型植硅體,而草原區(qū)草本植物群落則產(chǎn)生了較多指示溫暖環(huán)境的啞鈴型植硅體。根據(jù)Ic指數(shù),本研究區(qū)內(nèi)的森林區(qū)草本植物群落Ic指數(shù)(Ic=27%)遠高于草原區(qū)(Ic=16%),這說明森林區(qū)草本植物群落產(chǎn)生的植硅體組合較草原區(qū)草本植物群落植硅體組合具有更強的寒冷環(huán)境的指示意義。
References:
[1] Li S Y, Li X B, Ying G,etal. Vegetation indexes-biomass models for typical semi-arid steppe—A case study for Xilinhot in Northern China. Journal of Plant Ecology, 2007, 31(1): 23-31.
李素英, 李曉兵, 鶯歌, 等. 基于植被指數(shù)的典型草原區(qū)生物量模型——以內(nèi)蒙古錫林浩特市為例. 植物生態(tài)學報, 2007, 31(1): 23-31.
[2] Huang F, Lisa K, Xiong S F,etal. Holocene grassland vegetation, environment and human activities in the Eastern part of Nei Mongol. Science in China, 2004, 34(11): 1029-1040.
黃翡, Lisa K, 熊尚發(fā), 等. 內(nèi)蒙古中東部全新世草原植被、環(huán)境及人類活動. 中國科學, 2004, 34(11): 1029-1040.
[3] Zhang J P, Lü H Y, Wu N Q,etal. Phytolith evidence of millet agriculture during about 6000-2100cal. aB. P. in the Guanzhong Basin, China. Quaternary Sciences, 2010, 30(2): 287-297.
張健平, 呂厚遠, 吳乃琴, 等. 關中盆地6000-2100cal. aB. P. 期間黍、粟農(nóng)業(yè)的植硅體證據(jù). 第四紀研究, 2010, 30(2): 287-297.
[4] Gu Y S, Zhao Z J, Deborah M P. Phytolith morphology research on wild and domesticated rice species in East Asia. Quaternary International, 2013, 287: 141-148.
[5] Liu H Y, Jie D M, Liu L D,etal. The shape factors of phytolith in selected plants from Changbai Mountains and their implications. Quaternary Sciences, 2013, 33(6): 1234-1244.
劉洪妍, 介冬梅, 劉利丹, 等. 長白山區(qū)典型禾本科植物植硅體形狀系數(shù). 第四紀研究, 2013, 33(6): 1234-1244.
[6] Wang Y J, Lü H Y. The study of phytolith and its application. Beijing: China Ocean Press, 1992.
王永吉, 呂厚遠. 植物硅酸體研究及應用. 北京: 海洋出版社, 1992.
[7] Liu L D, Jie D M, Liu H Y,etal. Change characters ofPhragmitesaustralisphytolith in Northeast China. Chinese Journal of Plant Ecology, 2013, 37(9): 861-871.
劉利丹, 介冬梅, 劉洪妍, 等. 東北地區(qū)蘆葦植硅體的變化特征. 植物生態(tài)學報, 2013, 37(9): 861-871.
[8] Liu H Y, Jie D M, Liu L D,etal. The spatial distribution ofPhragmitescommunis phytoliths from Northeast China. Acta Micropalaeontologica Sinica, 2013, 30(2): 191-198.
劉洪妍, 介冬梅, 劉利丹, 等. 東北地區(qū)蘆葦植硅體形態(tài)的空間差異. 微體古生物學報, 2013, 30(2): 191-198.
[9] Jie D M, Liu Z Y, Shi L X,etal. Characteristics of phytoliths inLeymuschinensisfrom different habitats on the Songnen Plain in Northeast China and their environmental implications. Science in China, 2010, 40(4): 493-502.
介冬梅, 劉朝陽, 石連旋, 等. 松嫩平原不同生境羊草植硅體形態(tài)特征及環(huán)境意義. 中國科學, 2010, 40(4): 493-502.
[10] Qin L, Li J, Wang L,etal. The morphology and assemblages of phytolith in Pooideae from the Qinghai-Tibetan Plateau. Acta Palaeontologica Sinica, 2008, 47(2): 176-184.
秦利, 李杰, 旺羅, 等. 青藏高原常見早熟禾亞科植硅體形態(tài)特征初步研究. 古生物學報, 2008, 47(2): 176-184.
[11] Li Q, Xu D K, Lü H Y. Morphology of phytolith in Bambusoideae (Gramineae) and its ecological significance. Quaternary Sciences, 2005, 25(6): 777-784.
李泉, 徐德克, 呂厚遠. 竹亞科植硅體形態(tài)學研究及其生態(tài)學意義. 第四紀研究, 2005, 25(6): 777-784.
[12] Deng D S. The studies on phytolith system of Cyperaceae. Guihaia, 1998, 18(3): 204-208.
鄧德山. 莎草科植物硅酸體的研究. 廣西植物, 1998, 18(3): 204-208.
[13] Li Y Y, Leng X T. The comparative study of pollen-spora analysis and formation environment of peat mire between Northeast of China and Belarus. Journal of Northeast Normal University (Natural Science Edition), 1997, (3): 97-103.
李宜垠, 冷雪天. 中國東北地區(qū)與白俄羅斯泥炭沼澤的孢粉分析及形成環(huán)境對比. 東北師范大學學報(自然科學版), 1997, (3): 97-103.
[14] Zhao G S, Wang J B, Fan W Y,etal. Vegetation net primary productivity in Northeast China in 2000-2008: simulation and sea sonal change. Chinese Journal of Applied Ecology, 2011, 22(3): 621-630.
趙國帥, 王軍邦, 范文義, 等. 2000-2008年中國東北地區(qū)植被凈初級生產(chǎn)力的模擬及季節(jié)變化. 應用生態(tài)學報, 2011, 22(3): 621-630.
[15] Wang Y J, Lü H Y.Methods of phytolith analysis. Acta Botanica Sinica, 1994, 36(10): 797-804.
王永吉, 呂厚遠. 植物硅酸體的分析方法. 植物學報, 1994, 36(10): 797-804.
[16] Xu D K, Li Q, Lü H Y. Morphological analysis of phytoliths in Palmae and its environmental significance. Quaternary Sciences, 2005, 25(6): 785-982.
徐德克, 李泉, 呂厚遠. 棕櫚科植硅體形態(tài)分析及其環(huán)境意義. 第四紀研究, 2005, 25(6): 785-982.
[17] Twiss P C. Predicted world distribution of C3and C4grass phytoliths. Jr. Mulholland S C: Phytolith Systematics Emerging Issues, 1992.
[18] Liu H Y, Gu Y S, Tang Q Q,etal. Phytolith records of modern plant communities and surface soils on the Qingbang Island, Zhejiang Province, east China and its environmental significances. Acta Micropalaeontologica Sinica, 2017, 34(1): 77-83.
劉紅葉, 顧延生, 唐倩倩, 等. 浙江青浜島現(xiàn)代植物群落和表土植硅體研究及環(huán)境意義. 微體古生物學報, 2017, 34(1): 77-83.
[19] Guo M E, Jie D M, Ge Y,etal. Phytolith characteristics and their significance of environment in surface soils from wetlands of Changbai Shan area. Journal of Palaeogeography, 2012, 14(5): 639-650.
郭梅娥, 介冬梅, 葛勇, 等. 長白山區(qū)濕地表土植硅體特征及其環(huán)境意義. 古地理學報, 2012, 14(5): 639-650.
[20] Piperno D R, Pearsall D M. The silica bodies of tropical American grasses: morphology, taxonomy, and implications for grass systematics and fossil phytolith identification. Smithsonian Contributions to Botany, 1998, 85: 1-40.
[21] Lü H Y, Liu K B. Morphological variations of lobate phytoliths from grasses in China and the South-Eastern United States. Diversity and Distributions, 2003, 9: 73-87.
[22] Zheng S X, Shangguan Z P. Relationship between stomata parameters of plants and atmospheric CO2concentration change. Ecologic Science, 2005, 24(3): 264-267.
鄭淑霞, 上官周平. 陸生植物氣孔參數(shù)與大氣CO2濃度變化. 生態(tài)科學, 2005, 24(3): 264-267.
[23] Zhang L R, Niu H S, Wang S P,etal. Effects of temperature increase and grazing on stomatal density and length of four alpine kobresia meadow species, Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 2010, 30(24): 6961-6969.
張立榮, 牛海山, 汪詩平, 等. 增溫與放牧對矮嵩草草甸4種植物氣孔密度和氣孔長度的影響. 生態(tài)學報, 2010, 30(24): 6961-6969.
[24] Guo Y L, Wang J B, Ding B,etal. Regulation of stomatal movement under low atmospheric humidity. Plant Physiology Journal, 2014, 50(8): 1144-1150.
郭瑤琳, 王俊斌, 丁博, 等. 低空氣濕度下氣孔運動的調控. 植物生理學報, 2014, 50(8): 1144-1150.
[25] Huang F, Kealhofer L, Huang F B. Diagnostic phytoliths from Nei Mongol grassland. Acta Palaeontologica Sinica, 2004, 43(2): 246-253.
黃翡, Kealhofer L, 黃鳳寶. 內(nèi)蒙古典型草原禾本科植硅體形態(tài). 古生物學報, 2004, 43(2): 246-253.
[26] Diester-Haass L, Schrader H J, Thiede J. Sedimentological and paleoclimatological investigations of two pelagie ooze cores off Cape Barbas, North-West Africa. Meteor Forschung-Ergebnisse, 1973, 16: 19-66.
[27] Gu Y S, Pearsall D M, Xie S C,etal. Vegetation and fire history of a Chinese site in southern tropical Xishuangbanna derived from phytolith and charcoal records from Holocene sediments. Journal of Biogeography, 2008, 35: 325-341.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
