紅松人工林和相鄰次生林林下紅松天然更新種群格局分析
2018-03-20 09:22:33王琴香沈海龍和春庭NguyenThanhTuan
森林工程 2018年2期
王琴香,沈海龍,*,和春庭,Nguyen Thanh Tuan
0 引言
紅松(Pinus koraiensis)是溫帶地區地帶性頂極群落闊葉紅松林的建群種,其天然更新種群狀況對群落的穩定發展具有重要意義。因此,紅松人工更新一直是研究的重點[1],利用天然更新來維持和恢復重建闊葉紅松混交林也一直是森林經營管理的關注重點[2-18]。從20世紀50年代中后期開始,東北林區營造了大量紅松人工林,這些紅松人工林通常與周圍不同類型的天然次生林呈鑲嵌分布。
自然條件下,植物實現成功更新必須具備三個要素:種源、傳播和萌發幼苗[19],以及適合幼苗生長發育的微生境,如林隙[20]。目前,很多紅松人工林已經達到結實盛期,可以提供豐富的種源。紅松種子傳播需要依靠動物的藏食習性,而對更新有利的動物主要是松鼠和星鴉。松鼠飽食后,會把松子埋入土壤中,通常每簇種子數2~4粒[6,21],最多達 18 粒[6]。星鴉頸部舌下囊可容納大量松子,胃和舌下囊松子數量可達30~50粒[7]。正常條件下,紅松人工林及其相鄰次生林內均不缺乏傳播紅松種子的動物,種子傳播有保障。但是,近年來人為球果采摘強度很高,導致紅松更新的種源日漸減少[2-3]。這不僅直接減少了天然更新的種子來源,而且間接增加了種子傳播動物對埋藏種子的消耗,從而減少了地下種子庫的種源。在高強度球果采摘的條件下,紅松人工林及其周圍次生林中的紅松天然更新是否會受影響?其受影響的程度如何?這些問題尚不清楚。
本研究以48 a紅松人工林及其相鄰蒙古櫟(Quercus mongolica)次生林為對象,研究兩種林分內天然更新紅松幼苗、幼樹種群的特點,分析相鄰的紅松人工林和次生林林下紅松天然更新種群格局的特點和差異及其與球果采摘強度的關系,旨在為紅松天然更新的調控提供科學依據,并為紅松球果采摘強度的控制決策提供參考。
1 研究方法
1.1 研究地概況
調查樣地位于吉林省敦化市寒蔥嶺林場(43°00′~43°12′N,127°55′~128°05′E), 該 林 場屬于溫帶大陸性濕潤季風氣候,冬季干燥寒冷,夏季溫暖濕潤,年平均氣溫2.6℃,極端最高氣溫可達33.4℃,極端最低氣溫可達38.3℃,無霜期120 d左右,年平均降水量約621.4 mm。紅松人工林樣地設置在12林班46小班的紅松人工林,樣地坡向西,坡度13°,土壤為典型暗棕壤;林齡48 a,平均樹高15.5 m,胸徑20.4 cm,平均郁閉度0.7;草本層平均蓋度40%,平均高為20 cm。蒙古櫟次生林樣地設置在同一林班的51小班,樣地坡向北,坡度14°,郁閉度0.7,土壤為典型暗棕壤;平均年齡59 a,平均樹高16.2 m,平均胸徑14.6 cm;下木高度1.6 m,蓋度30%;草本層平均高度30 cm,蓋度40%。
1.2 調查方法
考慮到紅松幼苗矮小不易發現,為了保證數據的精確度,選擇了草木還沒完全覆蓋地面的六月份開展調查。在緊鄰紅松人工林的蒙古櫟次生林林分內設置15個400 m2的樣帶(長×寬=20 m×20 m)調查次生林內紅松天然更新,在紅松人工林內的三個坡位上分別設置700 m2的三個條形樣帶(長×寬=70 m×10 m)。測量樣地內所有紅松幼苗幼樹的年齡、輪枝數、株/簇數、高度、第一活枝高以及生長情況。幼苗幼樹高度使用卷尺測量,測量精度為0.1 cm,地徑使用電子游標卡尺測量。并記錄樣地內上層木的樹種、樹高、胸徑和草本層的種類、蓋度和平均高度。
2 結果與分析
2.1 紅松人工林和蒙古櫟次生林林內紅松天然更新種群的年齡結構
由圖1(a)可知,在蒙古櫟次生林中,紅松天然更新種群密度隨年齡變化出現兩個峰值,第一個峰值在1a時出現,更新密度達到515株/hm2,然后隨年齡增加天然更新種群密度一直在下降,到14a時下降的趨勢變為增加的趨勢,到22 a時出現第二個峰值,更新密度達到32株/hm2。由圖1(b)可知,與蒙古櫟次生林相鄰,并為其林分內紅松更新提供種源的紅松人工林內紅松天然更新種群年齡結構不完整,最大只存在4 a更新苗,1 a更新苗的密度高達6 228株/hm2,以后隨苗齡增加,更新密度下降劇烈,2 a更新苗的密度為132株/hm2,3 a更新苗的密度為456株/hm2,4 a更新苗的密度已低至7株/hm2。
2.2 紅松人工林和蒙古櫟次生林1-3a紅松天然更新幼苗分布情況
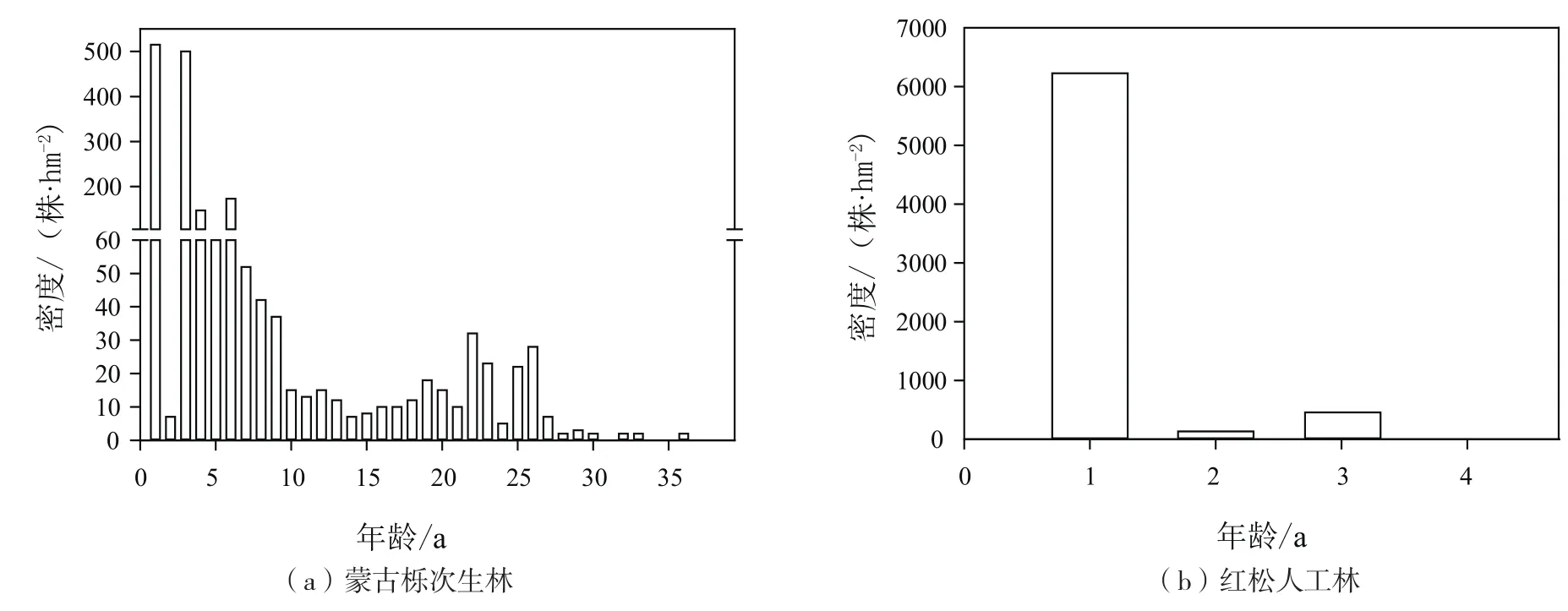
圖1 蒙古櫟次生林和紅松人工林林內天然更新紅松種群年齡結構Fig.1 Age structures of Korean pine natural regenerated seedlings in Mongolian oak secondary forest and Korean pine plantation
由圖2可知,紅松人工林和蒙古櫟次生林兩種林分中,紅松天然更新1~3 a幼苗的分布不同于以往的“山”狀分布—以簇生為主,而是以單株分布為主,隨株/簇的增加,簇數呈指數下降,分布圖表現為倒“J”形。在紅松人工林林內更新苗簇數與株數的相關系數介于0.935 4~0.998 2,其中2 a苗的相關系數最大,達到0.998 2。蒙古櫟次生林內更新苗簇數與株數的相關系數介于0.926 6~1,其中2 a苗的相關系數高達1。由圖2(a)和圖2(d)可知,1 a更新苗在紅松人工林和蒙古櫟次生林兩種林分中的總簇數分別是1 746簇和157簇,人工林內1 a苗簇數約是次生林的11倍;由圖2(b)和圖2(e)可知,2 a更新苗在兩種林分中的總簇數分別是63簇和7簇,人工林內2 a苗簇數是次生林的9倍;由圖2(c)和圖2(f)可知,3 a更新苗在兩種林分中的總簇數分別是187簇和203簇,二者數量接近,比值約為1。考慮到自然條件下紅松種子要經過兩個冬天才能打破休眠而萌發,結合紅松種子的豐欠年,1 a更新苗(產生種子時間:2014年,平年)、2 a更新苗(產生種子時間:2013年,欠年)、3 a更新苗(產生種子時間:2012年,豐年),結合圖2可知,在豐年,球果采摘對不同林分類型的影響差別不大,次生林內也有相當多的種子,而在平年和欠年,球果采摘嚴重影響了種源附近林分的紅松天然更新,差別能高達9~11倍。
2.3 紅松人工林和蒙古櫟次生林林內1~3a紅松天然更新幼生長情況
由圖3可知,在紅松人工林中紅松天然更新1,2,3 a幼苗高度分別為6.02、7.56、9.67 cm,地徑分別為1.35、1.18、1.55 mm,高度表現為增長趨勢,地徑為先下降后增長。在蒙古櫟次生林中紅松天然更新1,2,3 a幼苗高度分別為7、8.2、12.24 cm,地徑分別為:1.86、1.94、2.72 mm,高度和地徑都表現出增加趨勢。總的來說,蒙古櫟次生林中紅松天然更新幼苗生長情況優于紅松人工林,蒙古櫟次生林中1 a和2 a紅松幼苗的高度大于紅松人工林中相同年齡的更新苗,但差異不顯著(p>0.05),而3 a更新苗的高度卻極顯著大于紅松人工林中的更新苗(p<0.001)。在自然環境下,兩種林分內1 a、2 a到3 a紅松天然更新幼苗的地徑之間差異均極顯著(p<0.001),并且兩種林分內紅松天然更新苗地徑差異隨著苗齡增加而變得越來越大(1 a更新苗差值為0.51 mm,2 a更新苗差值為0.76 mm,3 a更新苗差值為1.17 mm)。
3 結論和討論
種群年齡結構分析是揭示種群現狀和更新的重要途徑之一[22],一個健康發展的種群,其年齡結構往往呈現倒J形,幼苗數量最多,隨年齡增長數量不斷減少[23]。而在本次研究調查中,蒙古櫟次生林和紅松人工林內紅松天然更新種群年齡結構皆不是健康發展種群的倒J形。蒙古櫟次生林中兩個更新峰值的出現代表了兩種球果采摘強度下的紅松天然更新情況,峰值分別出現在1 a和22 a,種群年齡結構表現為先下降后上升的趨勢;在14 a的時候由下降轉為上升,原因可能是以14a是兩個不同采摘強度(強度一:豐年采摘,欠年不采;強度二:豐年、欠年全采)的分界點。結合寒蔥嶺當地的球果承包采摘歷史,應該是近21年來采摘嚴重,采摘方式是豐欠年全樹全采,種源缺少了導致苗齡<22 a的紅松幼苗數量的不足;而22年前(1994年)松子價格低廉,球果采摘只集中在豐年,所以1994年以前紅松種群是健康發展的,現在種群結構的不完整的主要原因是近21年來不斷增強的球果采摘造成的。人工林有大量的萌發幼苗,但這些幼苗在第三年后死亡率幾乎達到100%,具體原因尚待研究。
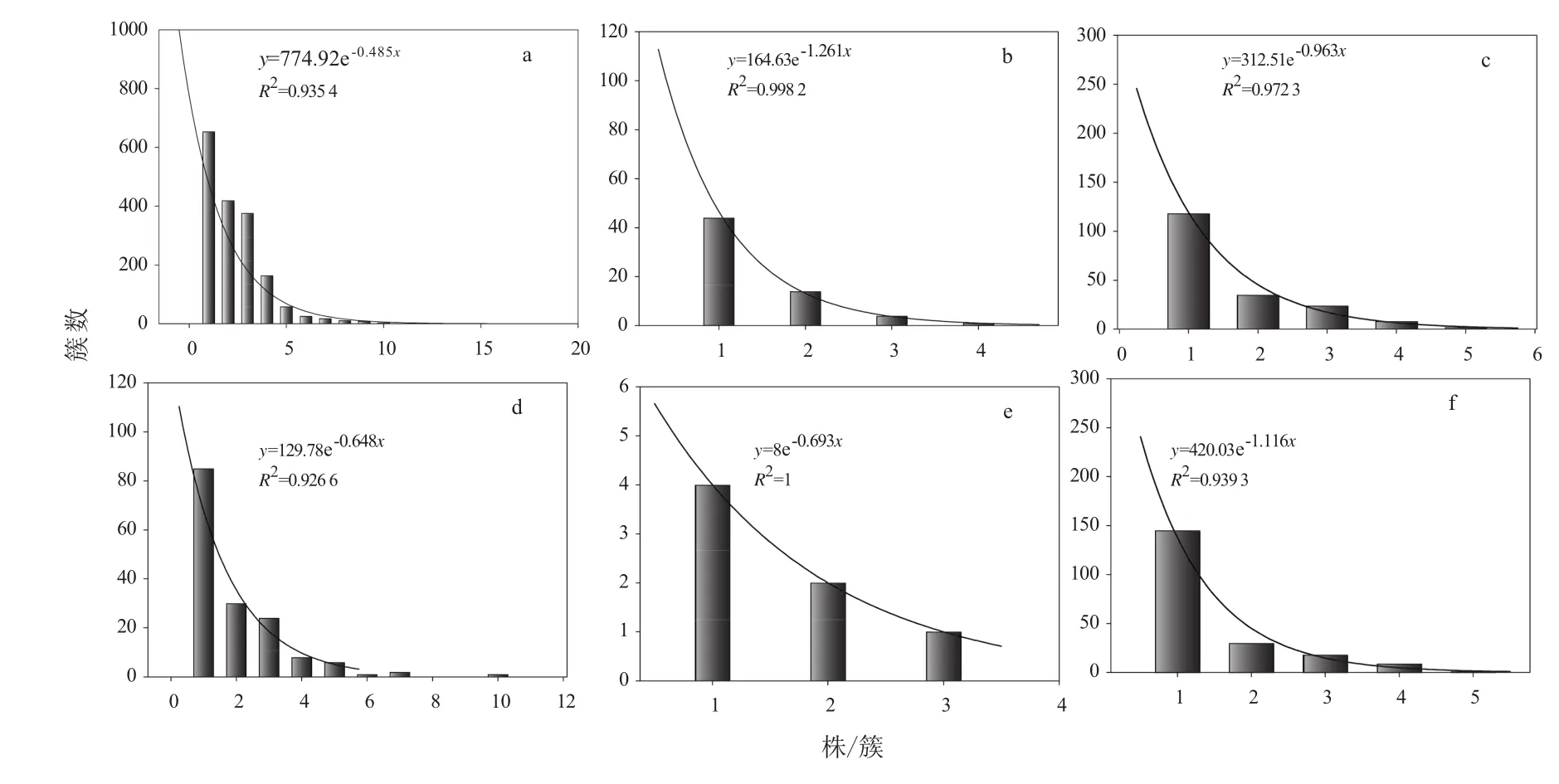
圖2 紅松人工林和蒙古櫟次生林林內1~3 a幼苗的株/簇分布Fig.2 The distribution of Korean pine 1 to 3 year old seedlings regenerated in Korean pine plantation and Mongolian oak secondary forest
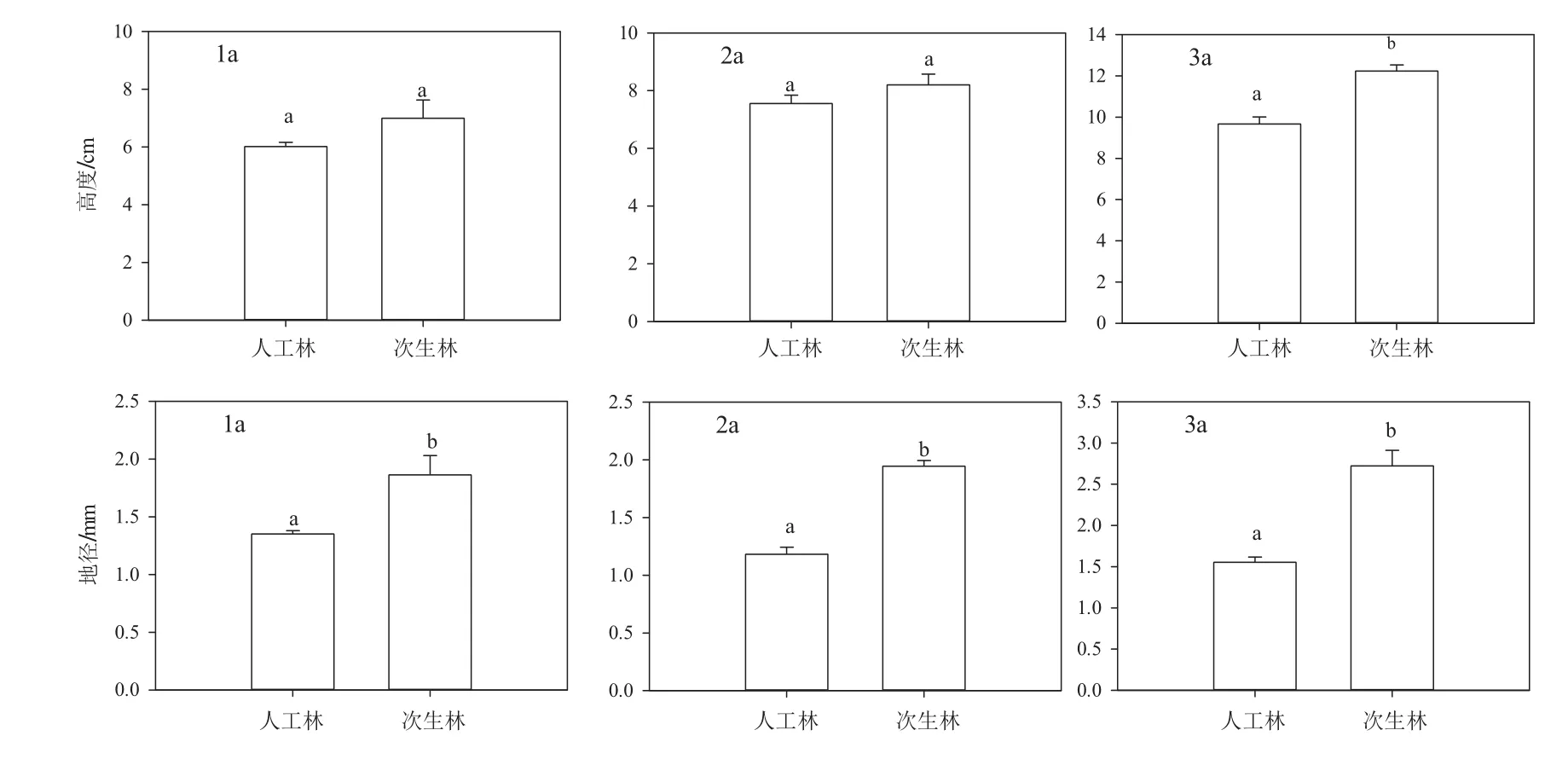
圖3 蒙古櫟次生林和紅松人工林內天然更新紅松幼苗苗高和地徑生長狀態Fig.3 The height and diameter growth of Korean pine seedlings natural regenerated in Mongolian oak secondary forest and Korean pine plantation
根據紅松更新與傳播動物之間的聯系,松子以多粒被埋藏于土壤中,據上世紀80年代松子埋藏情況調查,單粒占全部埋藏種子數的9.7%,2~4粒 /簇占 60.3%,5~7粒 /簇占 16.6%,8~10粒 /簇占 5.7%,10~20粒 /簇占 7.6%[24]。由前人研究可知,埋藏種子/簇的分布情況呈山狀,峰值出現在2~4粒/簇。而一年生幼苗直接來自于種子庫,所以1 a萌發幼苗分布與種子庫種子分布相似,只是萌發幼苗的數量少于種子的數量[25]。因而,1 a幼苗株/簇與簇數之間理論上應表現出“山”狀分布(由1 a轉化的2 a苗和3 a的分布情況也同理),幼苗多以2~4株/簇分布,單株和4株以上分布逐漸減少。然而在本研究中,不論是人工林、次生林,還是1、2、3 a苗,幼苗都以單株存在為主,隨每簇株樹的增加,簇數呈指數下降,分布圖表現為倒“J”形。高的單株率的形成有以下兩方面原因,一是高的動物遇見率,球果采摘后,捕食者在冬天食物不足時,就會多次重復取食,使得種子多呈單粒存在[3];二是自然環境的影響,幼苗生命力脆弱存活不易,死亡率很高。紅松幼年耐蔭且適當遮陰也有利于幼苗的生長[9-12],即使在郁閉度達到0.8~0.9的柞樹紅松林中天然更新也很好[13]。而次生林不同光照條件下紅松3a幼苗栽植實驗證明,4 a幼苗可在樹冠開度7%以上生長[14]。這說明,自然環境影響應該不是導致這種分布狀態的主要原因。高強度球果采摘導致的種子庫種源不足、動物取食種子和幼苗頻度大大增加,可能是主要的原因。
有研究用紅松人工林下土壤和枯落葉凋落物浸提液培育紅松種子,結果表明,人工林下浸提液抑制了紅松幼苗的生長[26]。在本次研究中,蒙古櫟次生林中紅松天然更新幼苗生長情況優于紅松人工林,比較二者可知,蒙古櫟次生林的林內環境有利于紅松天然更新幼苗生長,紅松人工林內的環境不利于紅松天然更新幼苗的生長,更新幼苗生長緩慢,長勢越來越弱直至死亡,這是紅松人工林內沒有更大年齡更新苗的一個原因。
[1]馮志忠, 劉云洲, 宋海玉, 等. 闊葉紅松林擇伐強度及更新技術的研究[J]. 森林工程, 2001, 17(2): 1-2, 16.
[2]劉足根, 姬蘭柱, 郝占慶, 等. 松果采摘對長白山自然保護區紅松天然更新的影響[J]. 應用生態學報, 2004, 15(6): 958-962.
[3]金光澤, 楊桂燕, 馬建章, 等. 松果采摘對小興安嶺主要林型紅松土壤種子庫和幼苗庫的影響[J]. 自然資源學報, 2010,25(11): 1845-1854.
[4]魯長虎. 動物與紅松天然更新關系的研究綜述[J]. 生態學雜志, 2003, 22(1): 49-53.
[5]陶大立, 趙大昌. 紅松天然更新對動物的依賴性—一個排除動物影響的球果發芽實驗[J]. 生物多樣性, 1995,3(3): 131-133.
[6]魯長虎, 劉伯文, 吳建平. 闊葉紅松林中星鴉和松鼠對紅松種子的取食和傳播[J]. 東北林業大學學報, 2001, 29(5): 96-98.
[7]趙錫如. 星鴉與紅松更新關系[J]. 林業實用技術, 1987 (9): 32.
[8]王樹力, 葛劍平, 徐繼成, 等. 小興安嶺楊樺林下紅松種群天然更新的格局與過程[J]. 東北林業大學學報, 1993,21(5):7-13.
[9]王樹力, 武敬輝, 史永純. 紅松種群天然更新及幼年生長與林分結構關系的研究[J]. 吉林林學院學報, 1998, 14 (1): 6-10.
[10]教士奇, 劉大興, 聞殿墀. 紅松闊葉樹混交林的采伐方式與更新方法[J]. 林業科學, 1966, 11(2): 106-113.
[11]朱濟凡, 劉慎諤, 王戰, 等. 小興安嶺紅松針闊葉混交林[J].林業科學, 1958, 4(4): 355-369.
[12]金鑫, 胡萬良, 丁磊, 等. 遮陰對紅松幼苗生長及光合特性的影響[J]. 東北林業大學學報, 2009, 37(9): 12-13.
[13]徐振邦, 代力民, 陳吉泉, 等.長白山紅松闊葉混交林森林天然更新條件的研究[J].生態學報, 2001, 21(9): 1413-1420.
[14]Sun Y R, Zhu J J, Sun O J, et al. Photosynthetic and growth responses of Pinus koraiensis, seedlings to canopy openness:Implications for the restoration of mixed-broadleaved Korean pine forests[J]. Environmental & Experimental Botany, 2016,129: 118-126.
[15]劉宏偉,王國義.紅松果林營建與發展的幾點建議[J].林業科技, 2016, 41(4) : 52-54.
[16]孫浩然,孟慶彬,張巍.小興安嶺林區紅松不同家系子代生活力與出苗率測定[J].林業科技, 2016, 41(3) : 7-9.
[17]郭樹平. 小興安嶺闊葉紅松林林分結構與修復[J]. 森林工程, 2014, 30(6) : 8-12.
[18]鄭穎.不同林齡紅松優樹結實的經濟效益分析[J].林業科技, 2016, 41(2) : 44-45.
[19]肖治術, 張知彬, 王玉山. 以種子為繁殖體的植物更新模型研究[J]. 生態學雜志, 2003, 22(4): 70-75.
[20]于金瑩, 亢新剛. 林隙影響天然林更新研究進展[J]. 森林工程, 2005, 21(5): 1-4.
[21]Hayashida M. Seed dispersal by red squirrels and subsequent establishment of Korean pine[J]. Forest Ecology &Management, 1989, 28(2):115-129.
[22]Brotherson J D, Rushforth S R, Evenson W E, et al. Population dynamics and age relationships of 8 tree species in Navajo National Monument, Arizona[J]. Journal of Range Management, 1983,36(2):173-196.
[23]梁士楚, 李久林, 程仕澤. 貴州青巖油杉種群年齡結構和動態的研究[J]. 應用生態學報, 2002, 13(1): 21-26.
[24]劉慶洪. 紅松闊葉林中紅松種子的分布及更新[J].植物生態學與地植物學學報.1988, 12(2):134-142.
[25]劉慶洪. 小興安嶺紅松種群天然更新的特點[J].林業科學,1987, 23(3): 265-276.
[26]陳立新, 李少博, 喬璐, 等. 凋落物葉和土壤浸提液對紅松種子萌發及幼苗生長的影響[J].南京林業大學學報(自然科學版).2016, 40(2): 81-87.
