高溫脅迫后小白菜的補償生長能力及防御機制*1
2018-03-19 05:18:56孫晨曦趙和麗鄭箐舟宋星林
中國農業氣象 2018年2期
關鍵詞:生長
韓 瑋,孫晨曦,趙和麗,胡 琪,鄭箐舟,宋星林
(1.南京信息工程大學應用氣象學院/江蘇省農業氣象重點實驗室,南京 210044;2.山東省環境規劃研究院,濟南 250100)
小白菜(Brassica chinensis L.)又名不結球白菜、青菜、油菜。原產于中國,在中國栽培十分廣泛,南北各地均有分布,其生長快,產量高,消費量居各類蔬菜之首[1]。由于其喜冷涼氣候條件,夏季高溫障礙導致生長緩慢,且在自然條件下,夏季氣溫波動較大,研究極端高溫天氣過后,小白菜的生長恢復情況具有重要意義。
高溫對植物生長一般有負面影響,但是在自然選擇的作用下,生物存在補償能力,能夠減少或消除脅迫所帶來的不利影響,這是對環境變化的一種適應性[2]。補償生長表現為低補償、等補償和超補償,補償類型因物種、生長期和脅迫程度而異[3-5],輕度脅迫條件下植物會發生超補償或等量補償,但隨著脅迫強度的增大,可出現低補償反應[6-7]。李留勇[8]發現,早期高溫處理結束后,水稻干物質增加量表現出超補償效應,這種補償效應可能在減輕水稻高溫傷害方面起一定的作用。孫憲芝等[9]發現高溫脅迫解除后,遭受低強度脅迫的菊花植株可較快恢復生長,高強度脅迫導致幼苗出現不可逆傷害。李婷等[10]發現經過 6d的高溫脅迫后,銅綠微囊藻生長速率高于對照,出現超補償生長。補償生長的出現可能歸因于某些植物應答機制,諸如光合作用活性增強、葉綠素含量提高、抗氧化酶活力增強,生長率增大等[11-13]。
前人對高溫后植物補償生長方面有過不少研究,但是對于小白菜的研究多偏重于高溫脅迫對白菜生理生化指標的影響[14-16],如高溫脅迫降低小白菜生長量和葉片凈光合速率[14],提高細胞膜傷害率和丙二醛含量等[15],對高溫脅迫后小白菜的補償生長卻鮮有研究。隨著全球氣候變暖,高溫成為作物生長面臨的主要逆境[17-20]。但是,自然條件下,高溫天氣往往是不連續的,對高溫過后小白菜在補償性生長方面的生理生化機制進行研究,有助于了解小白菜補償生長中的特點、規律和生理機制,對準確估測小白菜高溫脅迫后的產量補償能力具有重要意義,可為自然條件下小白菜合理規避高溫風險提供指導。本研究針對小白菜的高溫傷害癥狀、產量補償能力、生理生化作用對不同時間高溫脅迫的響應展開研究,以期查明小白菜的補償生長能力并從生物內源性因素方面探討小白菜的補償機制,為夏季小白菜產量品質預測及環境調控提供理論依據和技術支持。
1 材料與方法
1.1 實驗設計
實驗于2017年3-4月在南京信息工程大學江蘇省農業氣象重點實驗室的人工氣候箱內進行。以青梗類型“上海青”Brassica chinensis L.為實驗材料,選取籽粒飽滿,整齊一致的種子 6~8粒直播于高30cm,直徑20cm的塑料盆中?長至三葉一心時,每盆定植3株。栽培條件為:晝/夜溫度為25℃/18℃,光照條件2000lx,光暗周期12h/12h,空氣相對濕度75%,每2d澆1次水,用稱重法將土壤含水量控制在田間持水量的80%左右?養分條件維持在適宜水平?待小白菜長至 5片真葉時,選取長勢基本一致的植株進行高溫脅迫處理。根據長江中下游地區夏季常見的高溫天氣,實驗設高溫組(HT),其晝夜溫度設為 37℃/30℃,對照組(CK),晝/夜溫度為 25℃/18℃,每組3個重復。實驗分為2個階段:脅迫階段和恢復階段。高溫培養時間分別為3d、6d和9d,處理后,均置于晝/夜溫度為 25℃/18℃的光照培養箱內恢復生長9d。分別在高溫脅迫3、6、9d后以及恢復生長的3、6、9d后進行葉片光合參數測定,并取樣測定葉綠素含量、可溶性蛋白含量、丙二醛(MDA)和保護酶活性。由于高溫脅迫9d后的小白菜均在恢復過程中死亡,因此該處理無恢復期數據。
1.2 觀測項目
(1)小白菜干重測定:在高溫脅迫的 0d、3d、6d、9d以及恢復期的3d、6d、9d將植株地上部分和地下部分在通風干燥箱內 105℃殺青,然后 80℃烘至恒重,稱重?小白菜干物質相對增量指當次測量的植株干重與前一次測量的干重之差。
(2)光合參數的測定:選取植株第3或第4片展開葉,用LI-6400便攜式光合儀測定葉片的凈光合速率、胞間CO2濃度、氣孔導度、蒸騰速率,每個處理重復測定3次。葉室(2cm×3cm)內設定溫度20℃,輻射強度為 600μmolm-2·s-1。
(3)葉綠素含量的測定:選取每棵植株上第 3或第 4片展開葉,洗凈擦干,剪碎,稱取 0.2g,置于95%乙醇中48h直至葉片中的葉綠素完全被提取出?取提取液在紫外分光光度計UV-1800(日本島津)中采用比色法測定光密度值[21]?
(4)酶活性參數的測定:超氧化物歧化酶(SOD)活性用氮藍四唑(NBT)法測定,以抑制NBT光化學還原的50%為1個酶活性單位;過氧化物酶(POD)活性用愈創木酚法測定,以每克鮮重每分鐘在470nm處光吸收值變化0.1為1個酶活性單位;過氧化氫酶(CAT)活性測定用紫外吸收法,以每克鮮重每分鐘在240nm處光吸收值變化0.1為1個酶活性單位[22]。
(5)可溶性蛋白含量的測定:采用考馬斯亮藍G-250 法測定[22]。
(6)丙二醛(MDA)的測定:采用硫代巴比妥酸(TBA)法測定[22]。
1.3 數據分析
補償指數(G/C)為相同時段內實驗組與對照組生長量的比值,用以表示生物的補償生長模式。G為實驗組補償量,C為對照組生長量,G/C>1為超補償,G/C=1為等補償,G/C<1為低補償[6]。
應用SPSS19.0軟件,采用Duncan法進行差異顯著性分析。利用灰色關聯法對各指標與補償指數的關系進行分析。
2 結果與分析
2.1 高溫脅迫不同天數后小白菜的補償生長量比較
由圖 1a可以看出,與脅迫前相比,高溫脅迫3d、6d和9d后,小白菜生物量相對增加量逐漸減小,3d處理為0.309g,6d為0.072g,高溫處理9d后植株死亡,增加量為 0,而對照的生物量相對增加量穩定在 0.45g左右。可見,在此高溫條件下,小白菜植株生長受到明顯抑制,其中高溫處理 3d組受到的抑制最小,處理6d組居中,處理9d組的傷害最大,直接造成植株死亡。圖 1b顯示,處理3d組和處理6d組脅迫結束后,植株在適宜溫度下恢復,3d、6d和9d后其生長表現不同,脅迫3d組干物質增加量迅速上升,恢復生長3d后其增加量尚顯著低于對照,但恢復生長6天后,其干物質增加量等于或高于對照,而脅迫6d組生長逐步恢復,但其生長速率一直顯著低于對照;脅迫9d后小白菜多數死亡,即停止生長。說明經3d高溫脅迫后,小白菜的生長仍可達到正常水平,6d和9d的高溫則引起小白菜的不可逆損傷。
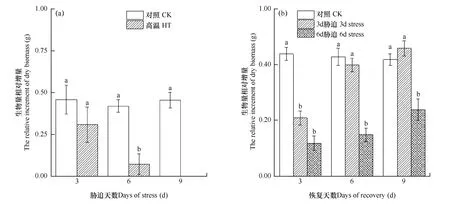
圖1 高溫脅迫后(a)及恢復后(b)小白菜生物量相對增量的比較Fig. 1 The relative increment of dry biomass of Chinese cabbage during the stress phase (a) and the recovery phase (b)
進一步計算脅迫結束恢復生長后小白菜的生長補償指數,結果見圖 2。由圖可見,3d脅迫組小白菜在恢復的6d以內補償指數均小于1,為低補償階段,但是在恢復的 6~9d則呈現超補償,補償指數達到1.10。6d脅迫組則在整個恢復期均呈現低補償。可見,經過不同時間的高溫脅迫小白菜出現了不同程度的補償效應,補償能力與脅迫時間有關。推測經過不同時間的高溫脅迫,小白菜的各項生理生化指標,如光合能力、保護酶活性等可能產生了一系列的應激調整,從而影響植株的補償能力。
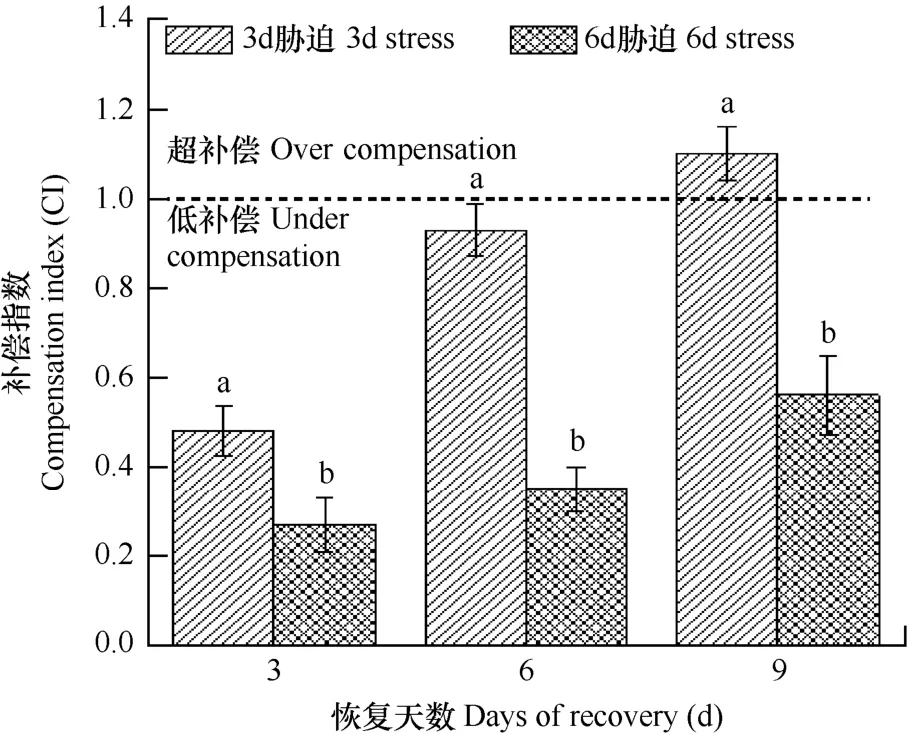
圖2 高溫脅迫后恢復期的補償指數Fig. 2 Compensation indices (CI) of the relative increment ofdry biomass of Chinese cabbage during the recovery phase
2.2 高溫脅迫不同天數后小白菜葉片葉綠素含量及光合特性的比較
高溫脅迫及恢復生長期小白菜葉片葉綠素總量隨時間的變化如圖 3所示。由圖可見,在高溫脅迫階段,脅迫組的葉綠素a含量在前3d呈上升趨勢,高于對照處理水平,之后則呈下降趨勢,9d低于對照水平,比對照下降了16.96%。脅迫3d、6d時葉綠素總量分別為3.03、2.82mg·g-1,比對照升高了28.26%和18.49%。在恢復階段,脅迫3d組葉綠素含量逐漸下降,處理組與對照組的差異逐漸減小,直至降至對照水平,而脅迫6d組,葉綠素含量則逐漸下降至低于對照水平。
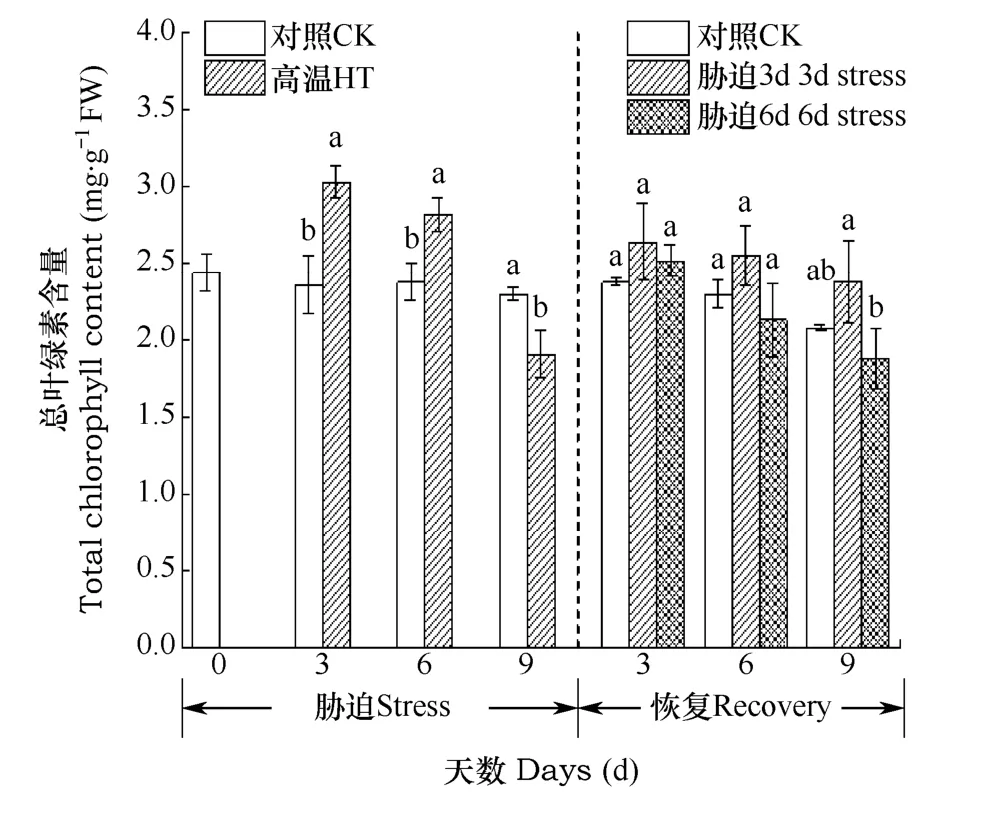
圖3 高溫脅迫(3、6、9d)后及恢復(3、6、9d)后小白菜葉綠素總量的比較Fig. 3 Total chlorophyll content of Chinese cabbage during the stress phase (3, 6, 9d) and the recovery phase (3, 6, 9d)
光合速率的高低是衡量葉片活力的重要生理指標,凈光合速率的高低基本反映了葉片光合生理功能的強弱。如圖4所示,在脅迫階段,葉片Pn呈持續下降趨勢,并在脅迫9d時出現負值。Gs和Tr也隨著高溫脅迫時間的增加而持續下降。可見高溫有可能影響了葉片上氣孔的開關程度,由于氣孔限制和高溫傷害共同造成光合系統效率降低。而 Ci隨著高溫脅迫時間的增加而持續上升,胞間 CO2濃度與凈光合速率始終呈現負相關,說明氣孔的開放程度與胞間 CO2濃度的相互作用影響了葉片的光合速率。在恢復階段,3d處理組葉片Pn、Gs、Tr快速上升并高于對照水平,6d處理組Pn、Gs、Tr雖也呈現上升趨勢,但始終低于對照。可見,3d的短期高溫脅迫可能觸發了了小白菜的某些防御應答機制,使光合速率出現超補償,有利于植株的快速恢復,說明 3d短期的高溫脅迫不足以對光合系統造成不可恢復性的影響。而6d長期高溫脅迫則會對小白菜造成不可逆的損害,即使在恢復期,光合能力也不能達到對照水平。
2.3 高溫脅迫不同天數后小白菜葉片 MDA、可溶性蛋白和保護酶活性的比較
蛋白質是生物體的重要組成部分,可調節植物生長發育和抗性形成等過程,大多數酶也是蛋白質,細胞在適應環境脅迫的過程中,需調節自身蛋白質的合成與分解。由圖5a可見,高溫脅迫期,小白菜可溶性蛋白含量顯著上升,顯著高于對照(圖5a),說明小白菜葉片在高溫脅迫下能迅速積累可溶性蛋白以抵御高溫脅迫的傷害。在恢復階段,3d處理組與6d處理組可溶性蛋白在恢復的前6d均繼續呈現上升趨勢,之后迅速下降至對照水平。
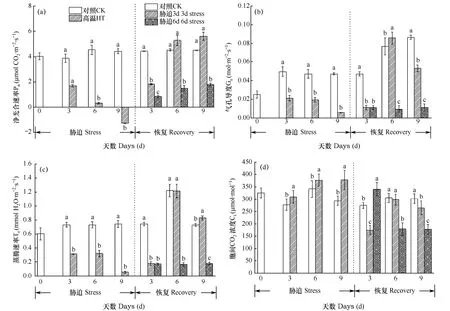
圖4 脅迫期及恢復期小白菜凈光合速率(a)、氣孔導度(b)、蒸騰速率(c)和胞間CO2濃度(d)的變化Fig. 4 Photosynthetic parameters Pn (a),Gs (b),Tr (c) 和 Ci (d) of Chinese cabbage during the stress phase and the recovery phase
MDA含量可反應細胞在高溫脅迫中受損傷的嚴重程度和抗逆性的強弱。由圖5b可以看出,高溫脅迫階段,隨著高溫脅迫時間的推移,MDA含量持續增加,含量顯著高于對照,高溫處理3d、6d和9d組的含量比對照分別提高 36.13%、57.66%和63.24%,脅迫時間越長,MDA含量越高,細胞受損越嚴重。在恢復階段,3d脅迫組和6d脅迫組MDA含量均呈下降趨勢,但是脅迫3d處理的MDA含量一直低于6d處理。
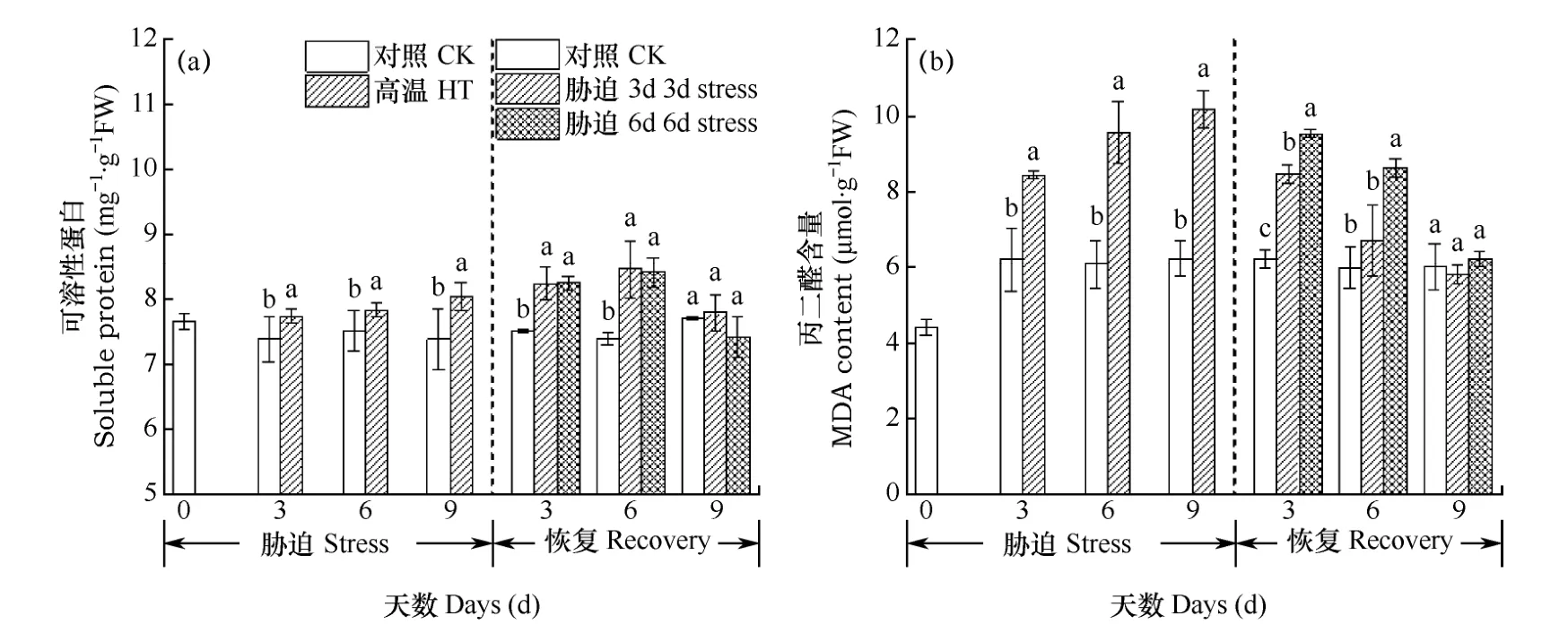
圖5 脅迫期及恢復期小白菜可溶性蛋白(a)和丙二醛(b)含量Fig. 5 Soluble protein (a) and MDA (b) contents of Chinese cabbage during the stress phase and the recovery phase
SOD、CAT、POD是抗氧化酶系的重要成員,負責清除有氧呼吸相伴產生的活性氧及自由基,以避免細胞質膜等受到過氧化損傷。從圖6可以看出,在高溫脅迫階段,SOD、CAT酶活性均呈上升趨勢,顯著高于對照,高溫處理3、6和9d組的SOD活性分別比對照提高21.66%、27.45%和24.88%,CAT活性分別比對照提高10.17%、22.13%和16.00%。因為脅迫會引起活性氧增加,細胞則通過提高自身抗氧化系統的酶活性和抗氧化劑的含量來清除過量的氧自由基。而恢復階段,保護酶的變化趨勢較復雜,3d處理組SOD、CAT、POD保護酶活性逐漸恢復至對照水平,與對照的差異逐漸減小。而6d處理組在恢復階段SOD和POD活性持續上升,高于對照水平,CAT持續下降,低于對照水平,出現一定的紊亂。
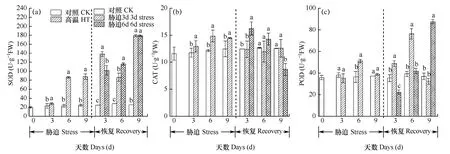
圖6 脅迫期及恢復期小白菜超氧化物歧化酶(a)、過氧化氫酶(b)和過氧化物酶(c)活性Fig. 6 SOD (a), CAT (b), and POD (c) activities of Chinese cabbage during the stress phase and the recovery phase
2.4 高溫脅迫不同天數后小白菜補償生長的機制分析
利用每個生理生化指標的歸一化數值繪制雷達圖,以確定小白菜生長限制指標(圖 7)。每個獨立的數軸對應一個生理生化指標,數軸呈輻射狀分布在中心點周圍,把不同數軸上的指標歸一化數值用折線連接起來所形成的曲線代表不同的處理。越靠近數軸的外側,代表對應指標有利于小白菜生長,越靠近數軸的原點,代表指標不利于小白菜生長。由圖可見,在3d處理組,各項指標歸一化指標評分均較高,各指標的評價值除CAT和MDA外均為最大值,評分結果所構成的多邊形面積在 3個處理中最大,可見,3d處理各項生長指標較好,在恢復過程中達到了較好狀態,因此,其恢復生長后期干重增長速率高于對照,可出現超補償現象;而對于 6d處理和 CK,評分結果構成的多邊形向圓心明顯收縮,說明各項指標綜合狀況較差。雷達圖不僅綜合反映了評價對象的好壞,更直接表現了其在單個指標上的優劣。在6d處理組,Pn、Gs、葉綠素和MDA含量的評價值靠近圓心,說明它在這方面具有很明顯的劣勢,Pn、Gs、葉綠素和MDA含量是6d處理的生長限制因素;在CK處理中,保護酶活性和可溶性蛋白含量的評價值靠近圓心,說明它在保護酶活性和可溶性蛋白方面較劣勢。
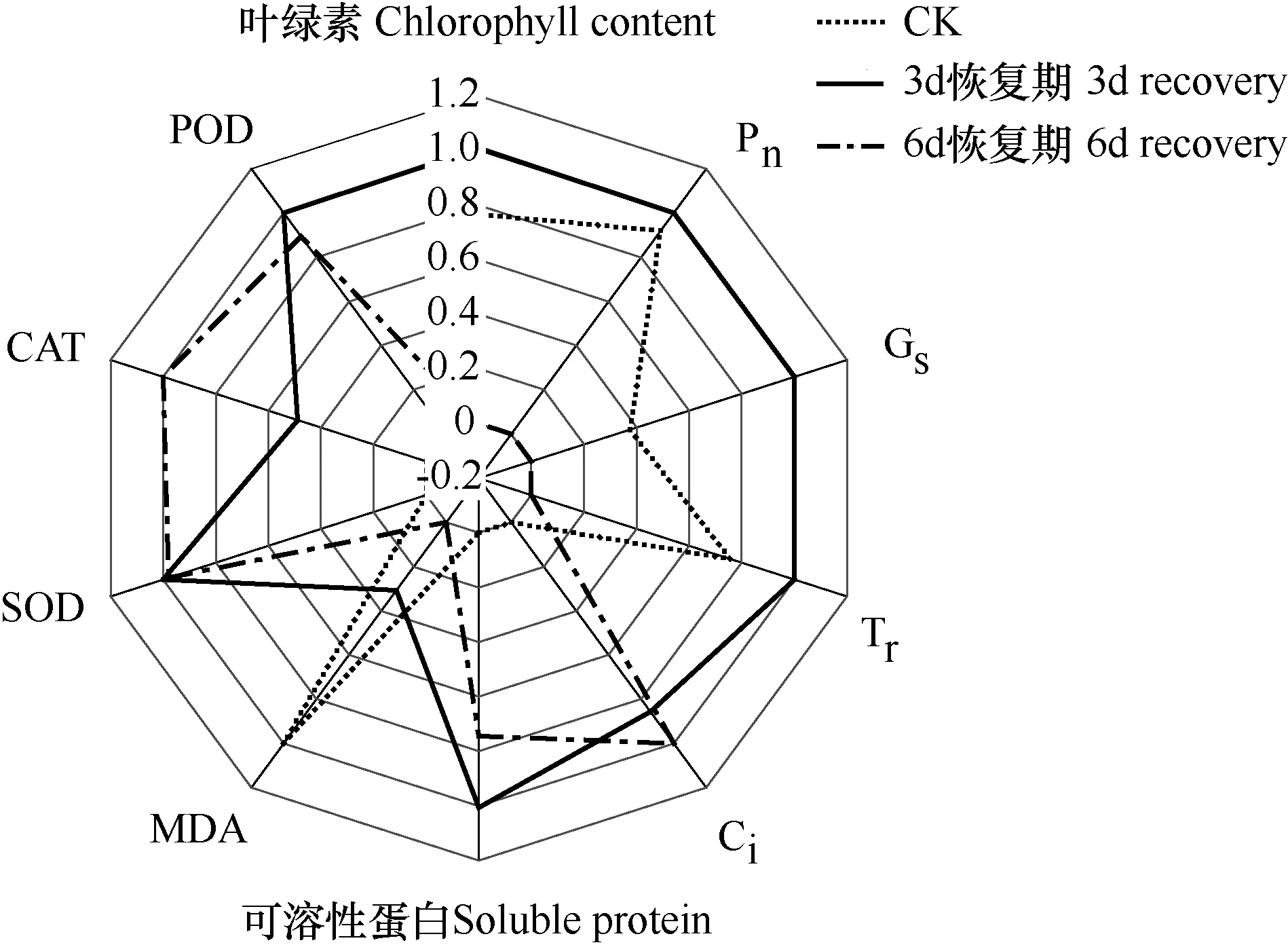
圖7 基于10項指標的小白菜恢復期生長分析Fig. 7 Description of Chinese cabbage growth based on the 10 factors of different treatments during the recovery phase
為了研究小白菜補償生長與生理生化指標的具體關系,利用灰色關聯法分析了10項具體的生理生化指標(葉綠素、Pn、Gs、Tr、Ci、可溶性蛋白、SOD、CAT、POD、MDA)與小白菜干物質增加量的關系。影響因素對目標因素的重要性越大,關聯系數越高。由表1可見,與小白菜干物質增加量關聯密切的因素主要有Pn(0.6287),其次是Gs(0.5518)、可溶性蛋白(0.4559)、MDA(0.4446)和葉綠素含量

表1 恢復期小白菜干重相對增量與生理生化指標之間的灰色關聯度Table 1 Grey correlation degree between Chinese cabbage absolute increment weight and biochemical indices during the recovery phase
3 結論與討論
灰色關聯分析表明,與小白菜恢復生長速率關系最密切的指標為 Pn。周瑞蓮等[12]研究也發現單葉蔓荊補償生長與Pn呈顯著正相關。有關研究表明,高溫直接傷害植物的光合系統[23]。在脅迫期,植物Pn下降,在恢復期,3d處理迅速恢復甚至超過對照水平,6d處理也有緩慢恢復,但是一直顯著低于對照水平,說明長期的高溫脅迫會對小白菜的光合作用造成嚴重影響。諸多其它作物上的實驗也證明了這點,孫憲芝等[9]研究表明,5d內33/28℃短期處理后菊花的光合作用可以在較短時間內得到恢復,而脅迫時間大于5d的Pn在短期正常條件下不能完全恢復。張潔等[24]研究表明,即使35℃亞高溫處理番茄2h,番茄葉片 Pn在一日內也很難完全恢復,處理時間越長,光合作用恢復的時間也將越長。然而王晨陽等[25]發現小麥經過36h的38℃高溫處理后,在恢復期小麥旗葉的Pn略呈上升趨勢。這與本研究中3d處理迅速恢復甚至超過對照水平相似。因此,小白菜對高溫脅迫較敏感,3d以上的高溫傷害將對產量造成嚴重影響。
根據灰色關聯分析,與小白菜恢復生長速率關系密切的指標為Gs。一般認為氣孔對CO2和水汽進出葉片的共同控制作用是葉片光合-蒸騰耦合關系形成的原因,它是決定植物光合強度和水分蒸騰強度的重要因素,氣孔調節在植物適應環境變化和環境壓力中起著關鍵作用[26]。
灰色關聯分析還表明,與小白菜生長速率關系密切的指標為可溶性蛋白。趙偉潔等[13]研究也發現,糜子的補償生長與可溶性蛋白含量呈顯著正相關。在脅迫期,植物可溶性蛋白一直呈上升趨勢,在恢復期的前3d植物可溶性蛋白繼續上升,之后下降至(0.4414),說明這些因子與小白菜干重增加有較密切關系,是影響小白菜補償生長速率的主要因素。正常水平。可溶性蛋白的積累可能是植物細胞適應高溫的一種保護策略,增加的蛋白質含量可能表明增強生理代謝[27]。一方面可能與合成更能適應溫度脅迫環境的蛋白即熱激蛋白有關[27];另一方面可能是高溫誘導或增加了某些抗逆性酶的含量和活性從而引起可溶性蛋白的積累[27]。高溫脅迫下小白菜葉片內的可溶性蛋白含量呈上升趨勢,表明小白菜體內的可溶性蛋白在抵御高溫脅迫過程中起到積極的作用。
MDA是膜脂過氧化過程中的分解產物,表示膜脂過氧化程度和對逆境反應的強弱。MDA含量越高,則損傷越大[21]。MDA具有細胞毒性,能引起細胞膜功能紊亂[28]。在高溫脅迫期,MDA含量累積,隨著脅迫時間的增加而增加。這表明雖然抗氧化酶隨脅迫時間的推移而升高,但增高的抗氧化酶活性仍然不足以應付越來越多的自由基,導致細胞膜脂過氧化程度加重,MDA增加。然而,在恢復期,MDA含量顯著下降,表明當受損細胞處于修復階段時,膜功能逐漸恢復,小白菜的生長逐漸恢復正常。
有研究發現,植物補償生長與葉綠素含量呈顯著正相關[13]。葉綠素是植物重要的光合色素,在光子捕獲、光能轉換和電子傳遞等過程中起重要作用,直接影響光合效率和代謝水平[21],這在植物對溫度的反應中起關鍵作用。在本研究中,高溫脅迫階段,葉綠素含量在前3~6d呈上升趨勢,高于對照水平,之后則呈下降趨勢,9d低于對照水平。在恢復階段,3d處理葉綠素含量下降直到接近正常水平,6d處理則降低直至低于對照水平。雖然多種作物葉綠素含量會隨高溫脅迫而下降,但是本研究中葉綠素含量在脅迫階段增加,與董瑜的研究一致[29-30],這可能與植物種類有關,可能是被響應和抗氧化相關基因在轉錄水平控制引起的,植物以此對抗高溫脅迫。
SOD、CAT和POD是細胞抵御活性氧傷害的重要保護性酶,它們在清除O2-、H2O2和過氧化物,阻止或減少羥基自由基形成,保持膜系統免受損傷方面起著重要作用,與植物的抗逆性密切相關[31-32]。正常條件下,植物細胞內自由基的產生與清除之間保持著動態平衡狀態,但在逆境條件下,植物細胞內的活性氧產生與清除的平衡關系受到破壞,積累的自由基就會對細胞造成傷害[33]。本實驗發現,SOD、CAT、POD含量在脅迫期大多呈現上升趨勢,恢復期則緩慢下降。這可能是植物的自身保護機制,熱應激可能誘發表達氧化代謝的變化,激活基因表達,增加抗氧化酶活性[34]。Wang等[35-36]也發現高溫脅迫下小麥SOD活性增加,查倩等[30]發現高溫導致抗氧化酶活性增高。當然過度的高溫傷害也會引起保護酶活性的降低[37]。
高溫是抑制夏季小白菜生長的環境因素之一。從本研究可以看出,小白菜在37/30℃高溫處理下生長受阻,高溫傷害對小白菜傷害主要是降低葉片光合速率,提高細胞膜脂過氧化程度,小白菜在不良條件下難以完成正常生理活動,從而導致細胞生長遲緩、產物合成能力下降[38]。超補償是生物與環境的長期博弈中形成的抗逆機理的重要組成部分,適度脅迫下,作物自身調控潛力可實現超補償生長[7]。本實驗中,不同脅迫時間處理補償能力不同,這與王利彬等[3]研究結論相近,即補償效應與脅迫程度有關,重度脅迫尤其是長時間重度脅迫會降低補償效應甚至產生傷害效應,而適度脅迫可產生超補償效應。3d脅迫組由于植株機體應激調整,啟動防御應答機制,保護酶活性上升,可溶性蛋白積累,適度防御高溫傷害,另外,3d脅迫使細胞內產生更多葉綠素,對葉片光合作用和恢復生長最為有利,Pn快速恢復,因此可以出現超補償生長。灰色相關分析表明,Pn、Gs、可溶性蛋白、MDA和葉綠素含量在恢復期補償生長中發揮了重要作用,是產生超補償生長的主要驅動因素。但在持續(高溫處理 6d)高溫下,Pn迅速下降,MDA含量迅速上升,即使保護酶活性上升,但仍然不足以有效清除嚴重高溫傷害引起的過量活性氧積累,而且積累過量的活性氧及其引發的膜脂過氧化產物反過來對防御體系起到破壞作用,從而加劇膜脂過氧化。因此,6d高溫脅迫后Pn恢復緩慢,小白菜表現為低補償生長。9d高溫脅迫后小白菜Pn下降,葉綠素含量降低,在恢復過程中死亡。整體來看,3d及其以上的高溫脅迫均會給小白菜生長帶來不可逆損傷,生產中要盡量避免高溫脅迫的影響,即使有高溫條件出現也應盡可能將高溫脅迫時間控制在3d以內,否則將對產量產生嚴重影響。
綜合看來,小白菜在37/30℃高溫處理下生長受阻,但是在脅迫解除后,3d組在恢復前6d呈現低補償,恢復的6~9d呈現超補償;6d組在整個恢復期均呈現低補償;9d組則在恢復過程中死亡。因此,超過3d的高溫脅迫將引起小白菜的不可逆損傷。小白菜在高溫脅迫下,啟動防御應答機制,Pn、Gs、可溶性蛋白、MDA和葉綠素含量是產生補償生長的主要驅動因素。
References
[1]譚文,楊再強,李軍.基于溫光效應的小白菜營養品質模擬模型研究[J].中國農業氣象,2016,37(1):59-67.Tan W,Yang Z Q,Li J.Simulation of nutrient quality of pakchoi based on temperature-light function[J].Chinese Journal of Agrometeorology,2016,37(1):59-67.(in Chinese)
[2]Ali M,Nicieza A,Wootton R J.Compensatory growth in fishes:a response to growth depression[J].Fish and Fisheries,2003,4(2):147-190.
[3]王利彬,祖偉,董守坤,等.干旱程度及時期對復水后大豆生長和代謝補償效應的影響[J].農業工程學報,2015,31(11):150-156.Wang L B,Zu W,Dong S K,et al.Effects of drought stresses and times on compensation effect after re-watering in soybean[J].Transactions of the CSAE,2015,31(11):150-156. (in Chinese)
[4]孫永江,杜遠鵬,翟衡.高溫脅迫下不同光強對‘赤霞珠’葡萄PSII活性及恢復的影響[J].植物生理學報,2014,50(8):1209-1215.Sun Y J,Du Y P,Zhai H.Effects of different light intensity on psii activity and recovery of Vitis vinifera cv. cabernet sauvignon leaves under high temperature stress[J].Plant Physiology Journal,2014,50(8):1209-1215.(in Chinese)
[5]韓冬,楊菲,楊再強,等.高溫對茶樹葉片光合及抗逆特性的影響和恢復[J].中國農業氣象,2016,37(3):297-306.Han D,Yang F,Yang Z Q,et al.Effects of high temperature stress and recovery on photosynthesis and stress tolerance of tea leaves[J].Chinese Journal of Agrometeorology,2016,37(3):297-306.(in Chinese)
[6]Belsky A J.Does herbivory benefit plants:a review of the evidence[J].The American Naturalist,1986,127(6):870-892.
[7]銀敏華,李援農,周昌明,等.調虧灌水和分蘗干擾對冬小麥生長的補償效應[J].應用生態學報,2015,26(10):3011-3019.Yih M H,Li Y N,Zhou C M,et al.Compensation effects of regulated deficit irrigation and tillering interference to winter wheat[J].Chinese Journal of Applied Ecology,2015,26(10):3011-3019.(in Chinese)
[8]李留勇.不同生育期高溫處理對水稻生長及產量形成的影響[D].武漢:華中農業大學,2015.Li L Y.Effects of high temperature treatments at different growth stages on the growth and yield formation of rice[D].Wuhan:Huazhong Agriculture University,2015.(in Chinese)
[9]孫憲芝,郭俊娥,鄭成淑.菊花的高溫傷害及生長恢復研究[J].山東農業大學學報:自然科學版,2013,(1):6-11.Sun X Z,Guo J E,Zheng C S.Study on heat damage and vegetative recovery of chrysanthemum[J].Journal of Shandong Agricultural University (Natural Science),2013,(1): 6-11.(in Chinese)
[10]李婷,景元書,韓瑋.亞高溫脅迫解除后銅綠微囊藻的生長恢復[J].應用生態學報,2014,25(11):3337-3343.Li T,Jing Y S,Han W.Recovery growth of Microcystis aeruginosa after the sub-high temperature stress[J].Chinese Journal of Applied Ecology,2014,25(11):3337-3343.(in Chinese)
[11]Han W,Jing Y S, Li T.Compensatory growth in Microcystis aeruginosa after moderate high-temperature exposure[J].Journal of Limnology,2015,74(3):549-558.
[12]周瑞蓮,賈有余,侯月利,等.不同厚度沙埋下植物光合特性變化與補償性生長的關系[J].生態學報,2016,36(24): 8111-8119.Zhou R L,Jia Y Y,Hou Y L,et al.Relationship between changes in photosynthetic characteristics and plant compensatory growth under different sandburial depths[J]. Acta Ecologica Sinica,2016,36(24):8111-8119.(in Chinese)
[13]趙偉潔,李翠,晁桂梅,等.復水對水分脅迫條件下糜子生長的補償效應[J].中國農業大學學報,2014,19(5):55-65.Zhao W J,LI C,Chao G M,et al.Compensation effect of rewatering on the growth of broomcorn millet under water stress[J].Journal of China Agricultural University,2014,19(5):55-65.(in Chinese)
[14]徐芬芬,柯維忠,王愛斌,等.NaCl 預處理緩解小白菜高溫脅迫的光合生理機制[J].生態科學,2016,35(3):161-164.Xu F F,Ke W Z,Wang A B,et al.Photosynthetic mechanism of NaCl pretreatment on Brassica campestris ssp.chinensis L.under high temperature stress[J].Ecological Science,2016,35(3):161-164.(in Chinese)
[15]劉燕燕,沈火林,劉以前.高溫脅迫對不結球白菜幼苗生長及生理指標的影響[J].華北農學報,2005,20(5):25-29.Liu Y Y,Shen H L,Liu Y Q.A study on the relationship between the growth and physiology of pakchoi and heat tolerance[J].Acta Agriculture Boreali-sinica,2005,20(5):25-29.(in Chinese)
[16]劉靜靜.氣候因子和肥料結構對小白菜生長和品質的影響[D].上海:上海交通大學,2013.Liu J J.Influence analysis of different cultivated evironments and fertilizers on pakchoi[D].Shanghai:Shanghai Jiao Tong University,2013.(in Chinese)
[17]Sánchez B,Rasmussen A,Porter J R.Temperatures and the growth and development of maize and rice:a review[J].Global Change Biology,2014,20(2):408-417.
[18]馮敏玉,祝必琴,雷俊,等.南昌高溫逼熟發生規律及其對早稻產量的影響[J].中國農業氣象,2014,35(3):287-292.Feng M Y,Zhu B Q,Lei J,et al.Characteristics of hightemperature forced maturity disaster and it’s impacts on early rice in Nanchang area[J].Chinese Journal of Agrometeorology,2014,35(3):287-292.(in Chinese)
[19]郭建茂,吳越,楊沈斌,等.典型高溫年不同播期一季稻產量差異及其原因分析[J].中國農業氣象,2017,38(2):121-130.Guo J M,Wu Y,Yang S B,et al.Yield differences and its causes for one season rice under different sowing dates in typical high temperature year[J].Chinese Journal of Agrometeorology,2017,38(2):121-130.(in Chinese)
[20]喻莎,陸魁東,謝佰承,等.高溫時數和熱積溫對超級早稻結實率的影響[J].中國農業氣象,2016,37(4):454-463.Yu S,Lu K D,Xie B C,et al.Effects of high temperature hours and thermal accumulated temperature on seed setting rate of super hybrid rice[J].Chinese Journal of Agrometeorology,2016,37(4):454-463.(in Chinese)
[21]李合生,孫群,趙世杰,等.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.Li H S,Sun Q,Zhao S J,et al.The experiment principle and technique on plant physiology and biochemistry[M].Beijing:Higher Education Press,2000.(in Chinese)
[22]陳建勛,王曉峰.植物生理學實驗指導[M].廣州:華南理工大學出版社,2002.Chen J X,Wang X F.Plant physiology experimental guidance[M].Guangzhou:South China University of Technology Press,2002.(in Chinese)
[23]Mathur S,Agrawal D,Jajoo A.Photosynthesis:response to high temperature stress[J].Journal of Photochemistry and Photobiology B:Biology,2014,137:116-126.
[24]張潔,李天來.短時間亞高溫處理及其恢復對番茄光合特性的影響[J].農業工程學報,2007,23(1):162-167.Zhang J,Li T L.Effects of short time sub-high temperature treatment and its recovery on photosynthetic characteristics of tomato(Lycopersicon.esculentum Mill.)[J].Transactions of the CSAE,2007,23(1):162-167.(in Chinese)
[25]王晨陽,郭天財,閻耀禮,等.花后短期高溫脅迫對小麥葉片光合性能的影響[J].作物學報,2004,30(1):88-91.Wang C Y,Guo T C,Yan Y L,et al.Effects of short postanthesis high temperature stress on leaf photosynthetic potential in winter wheat(Triticum aestivum L.)[J].Acta Agronomica Sinica,2004,30(1):88-91.(in Chinese)
[26]范嘉智,王丹,胡亞林,等.最優氣孔行為理論和氣孔導度模擬[J].植物生態學報,2016,40(6):631-642.Fan J Z,Wang D,Hu Y L,et al.Optimal stomatal behavior theory for simulating stomatal conductance[J].Chinese Journal of Plant Ecology,2016,40(6):631-642.(in Chinese)
[27]曹云英,段驊,王志琴,等.高溫對水稻葉片蛋白質表達的影響[J].生態學報,2010,(22):6009-6018.Cao Y Y,Duan Y,Wang Z Q,et al.Influence of protein expression in rice leaves under heat stress[J].Acta Ecologica Sinica,2010,(22):6009-6018.(in Chinese)
[28]徐敦明,馬志卿,馮俊濤,等.毒死蜱和鬼臼毒素脅迫對蔬菜上海青抗氧化酶系及丙二醛的影響[J].農業環境科學學報,2004,23(6):1089-1092.Xu D M,Ma Z Q,Feng J T,et al.Effects of chlorpyrifos and podophyllotoxin on antioxidative enzymes and the mda of vegetable(Brassica raps L.)[J].Journal of Agro-Environment Science,2004,23(6):1089-1092.(in Chinese)
[29]董瑜,田昆,郭緒虎,等.不同區域氣候條件影響下的納帕海濕地植物葉綠素熒光特性[J].生態環境學報,2013,22(4):588-594.Dong Y,Tian K,Guo X H,et al.Impacts of regional climate change on chlorophyll fluorescence characteristics of plateau wetland plants in Napahai,Yunnan China [J].Ecology and Environmental Sciences,2013,22(4):588-594.(in Chinese)
[30]查倩,奚曉軍,蔣愛麗,等.高溫脅迫對葡萄幼樹葉綠素熒光特性和抗氧化酶活性的影響[J].植物生理學報,2016,52(4):525-532.Zha Q,Xi X J,Jiang A L,et al.Effects of heat stress on chlorophyll fl uorescence characteristics and antioxidant activity in grapevines(Vitis vinifera L.cv.Xiahei)[J].Plant Physiology Journal,2016,52(4):525-532.(in Chinese)
[31]Xu S,Li J,Zhang X,et al.Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation,antioxidant metabolites,and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress[J].Environmental and Experimental Botany,2006,56(3):274-285.
[32]Laxman R H,Rao N K S,Biradar G,et al.Antioxidant enzymes activity and physiological response of tomato (Lycopersicon esculentum M.) genotypes under mild temperature stress[J].Indian Journal of Plant Physiology,2014,19(2):161-164.
[33]Valentine J,Foote C S,Greenberg A,et al.Active oxygen in biochemistry[M].Berlin:Springer Science & Business Media,2012.
[34]Mittler R,Vanderauwera S,Gollery M,et al.Reactive oxygen gene network of plants[J].Trends in Plant Science,2004,9(10):490-498.
[35]Wang C,Wen D,Sun A,et al.Differential activity and expression of antioxidant enzymes and alteration in osmolyte accumulation under high temperature stress in wheat seedlings[J].Journal of Cereal Science,2014,60(3): 653-659.
[36]Wang X,Cai J,Liu F,et al.Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings[J].Plant Physiology and Biochemistry,2014,74:185-192.
[37]Rai N,Rai K K,Tiwari G,et al.Changes in free radical generation,metabolites and antioxidant defense machinery in hyacinth bean(Lablab purpureus.L) in response to high temperature stress[J].Acta Physiologiae Plantarum,2015,37(3):1-12.
[38]Wahid A,Gelani S,Ashraf M,et al.Heat tolerance in plants:an overview[J].Environmental and Experimental Botany,2007,61(3):199-223.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
