kre1Δ釀酒酵母孢子固定肌酐酶體系的構(gòu)建及其酶學(xué)分析
2018-03-16 08:16:52潘華平中西秀樹(shù)孔軍高曉冬
食品與發(fā)酵工業(yè) 2018年2期
關(guān)鍵詞:殼聚糖
潘華平,中西秀樹(shù),孔軍,高曉冬
(江南大學(xué) 生物工程學(xué)院,糖化學(xué)與生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無(wú)錫,214122)
二倍體釀酒酵母(Saccharomycescerevisiae)在缺乏氮源及處于非發(fā)酵型碳源的條件下會(huì)進(jìn)入減數(shù)分裂和產(chǎn)孢過(guò)程并最終形成子囊孢子[1]。酵母孢子壁的組成從內(nèi)到外依次為甘露糖層、葡聚糖層、殼聚糖層和二酪氨酸層[2](圖1-A)。研究發(fā)現(xiàn),各層孢子壁的形成時(shí)間是有先后順序的,即只有當(dāng)內(nèi)層的孢子壁形成后,外層的孢子壁才能依次形成[3]。最先合成的甘露糖層由大量的甘露糖蛋白組成[4];葡聚糖層主要是以β-1,3-葡聚糖為主鏈及β-1,6-葡聚糖為支鏈組成[5];Chs3是殼聚糖層中幾丁質(zhì)的合成酶[6],敲除CHS3后會(huì)導(dǎo)致殼聚糖層無(wú)法形成,進(jìn)而導(dǎo)致外層的二酪氨酸層缺失;Dit1是二酪氨酸合成酶,敲除DIT1后無(wú)法形成二酪氨酸層[7]。此外,有一些功能未知的基因,如OSW2,也會(huì)影響孢子壁的形成,敲除OSW2后雖然不影響各層孢子壁的形成,但會(huì)改變孢子壁的通透性,并表現(xiàn)出乙醚敏感性[4]。成熟的酵母孢子是一種休眠細(xì)胞,孢子壁中的殼聚糖層和二酪氨酸層賦予孢子抵抗外界脅迫(如有機(jī)溶劑,高溫及酶降解等)壓力的能力。研究表明二酪氨酸層不僅具有抗蛋白酶及糖苷水解酶類(lèi)降解的能力,而且可以阻擋孢內(nèi)的可溶性分泌蛋白外泌及阻止孢外各類(lèi)有害的酶擴(kuò)散進(jìn)入到孢子內(nèi)部[8-9]。
基于酵母孢子壁的獨(dú)特性,實(shí)驗(yàn)室利用酵母孢子開(kāi)發(fā)了一種新型的酶生物固定法,即釀酒酵母孢子微膠囊酶固定法。該方法的原理是將目標(biāo)酶的基因與酵母分泌信號(hào)肽連接重組轉(zhuǎn)入釀酒酵母孢子中,表達(dá)的目標(biāo)酶在往外分泌的過(guò)程中會(huì)被孢子壁中的殼聚糖層吸附住,同時(shí)也會(huì)被二酪氨酸層攔截,并阻擋其分泌到孢外,使目標(biāo)酶被包埋固定在孢子壁中。反應(yīng)時(shí),直接用孢子微膠囊酶進(jìn)行催化反應(yīng)即可。由于酵母孢子壁具有抗脅迫能力,因此,被固定在孢子壁中的酶也具有抗逆性。此外還可通過(guò)基因工程手段改變孢子壁的通透性,保留孢子微膠囊酶抗逆性的同時(shí),提高催化酶活(圖1-B)。
實(shí)驗(yàn)室前期研究表明,利用有孢子壁缺陷的酵母孢子(如osw2Δ,dit1Δ孢子)包埋固定的酶,其酶活顯著高于野生型孢子,而且osw2Δ孢子微膠囊酶還表現(xiàn)出較強(qiáng)的抗逆性及較好的可重復(fù)利用性等優(yōu)勢(shì)[10-12]。KRE1是孢子壁中β-1,6-葡聚糖合成的基因。敲除KRE1后,雖然不影響孢子壁的形成,但可能影響孢子壁的通透性并表現(xiàn)出乙醚敏感性[13]。為了驗(yàn)證kre1Δ孢子是否具有與osw2Δ孢子一樣的酶膠囊載體性能,本課題利用kre1Δ孢子及其他有孢子壁缺陷的酵母孢子對(duì)肌酐酶(creatininase CA)進(jìn)行固定,并對(duì)其酶學(xué)性能進(jìn)行了研究。
1 材料與方法
1.1 菌株、質(zhì)粒和引物
本研究所用的菌株、質(zhì)粒及引物詳見(jiàn)表1。
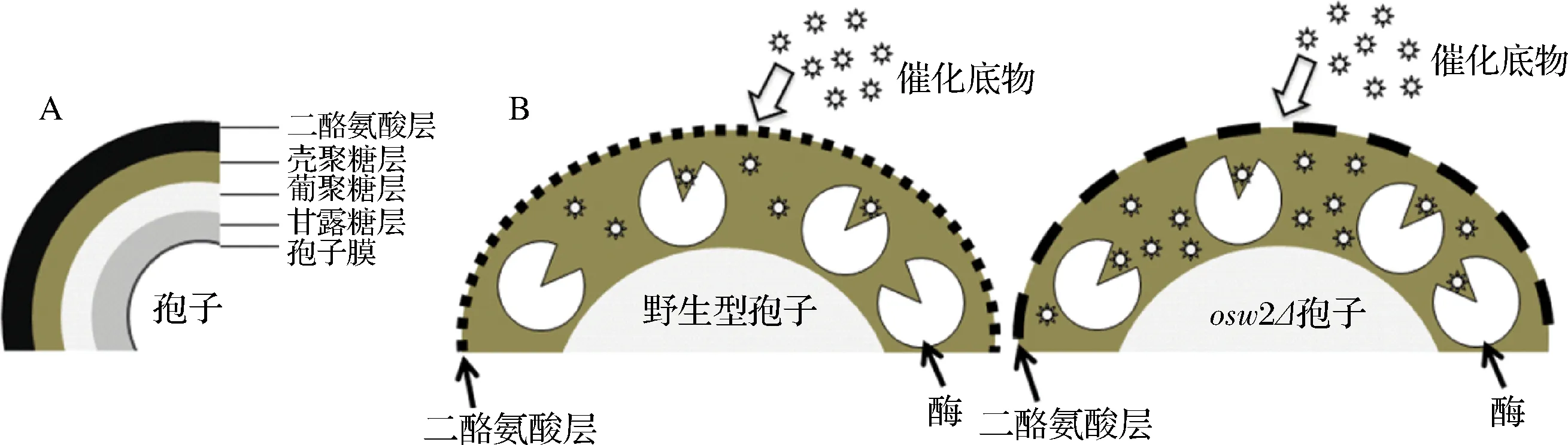
A-酵母孢子壁結(jié)構(gòu);B-孢子微膠囊酶催化反應(yīng)圖1 酵母孢子微膠囊酶固定化示意圖Fig.1 Yeast spore-encapsulated enzyme

菌株/質(zhì)粒/引物基因型/說(shuō)明/序列來(lái)源AN120MATα/MATaARG4/arg4-NspIhis3ΔSK/his3ΔSKho::LYS2/ho::LYS2leu2/leu2lys2/lys2RME1/rme1::LEU2trp1::hisG/trp1::hisGura3/ura3[14]chs3ΔMATα/MATaARG4/arg4-NspIhis3ΔSK/his3ΔSKho::LYS2/ho::LYS2leu2/leu2lys2/lys2RME1/rme1::LEU2trp1::hisG/trp1::hisGura3/ura3chs3Δ::his5+/chs3Δ::his5+[4]dit1ΔMATα/MATaARG4/arg4-NspIhis3ΔSK/his3ΔSKho::LYS2/ho::LYS2leu2/leu2lys2/lys2RME1/rme1::LEU2trp1::hisG/trp1::hisGura3/ura3dit1Δ::his5+/dit1Δ::his5+[15]osw2ΔMATα/MATaARG4/arg4-NspIhis3ΔSK/his3ΔSKho::LYS2/ho::LYS2leu2/leu2lys2/lys2RME1/rme1::LEU2trp1::hisG/trp1::hisGura3/ura3osw2Δ::his5+/osw2Δ::his5+[12]kre1ΔMATα/MATaARG4/arg4-NspIhis3ΔSK/his3ΔSKho::LYS2/ho::LYS2leu2/leu2lys2/lys2RME1/rme1::LEU2trp1::hisG/trp1::hisGura3/ura3kre1Δ::kanMX6/kre1Δ::kanMX6本研究質(zhì)粒pRS424?TEFpr?ss?CA?3HATRP1標(biāo)記多拷貝含信號(hào)肽的CA?3HA基因[11]引物HP27TATGCTACAGTAAAGACCTCTTCAACTTCTGCAAGACAATCAAAAAAAAACGGATCCCCGGGTTAATTAA本研究HP28ATTTTTACTCTTATATATGTATATGTTTGTGTTTATTTCTGAATAAAAATGAATTCGAGCTCGTTTAAAC本研究HP29TCATAAACTGAGAATGGGGC本研究HP30GCTGATGCATCAAGTTTGTC本研究
1.2 培養(yǎng)基、主要試劑和溶液
1.2.1 培養(yǎng)基
LB-氨芐培養(yǎng)基:蛋白胨10 g/L,酵母提取物5 g/L,NaCl 10 g/L,氨芐抗生素終質(zhì)量濃度 50 μg/mL,瓊脂粉20 g/L(固體培養(yǎng)基)。
YPAD培養(yǎng)基:酵母提取物10 g/L,Peptone-A 20 g/L,腺嘌呤0.03 g/L,葡萄糖 20 g/L,瓊脂粉20 g/L(固體培養(yǎng)基)。
YPAce培養(yǎng)基:酵母提取物10 g/L,Peptone-A 20 g/L,腺嘌呤0.03 g/L,醋酸鉀 20 g/L。
產(chǎn)孢培養(yǎng)基:醋酸鉀 20 g/L。
氨基酸缺陷型培養(yǎng)基(SD培養(yǎng)基):YNB 6.7 g/L,氨基酸缺陷型粉末 4 g/L,葡萄糖 20 g/L,瓊脂粉20 g/L(固體培養(yǎng)基)。
1.2.2 主要試劑
Lyticase,Creatinine 購(gòu)自Sigma-Aldrich公司(中國(guó),上海);酵母提取物、蛋白胨購(gòu)自O(shè)xoid公司;質(zhì)粒抽提試劑盒、Peptone-A、酵母基礎(chǔ)氮源培養(yǎng)基(YNB)、氨芐抗生素、腺嘌呤、脫脂奶粉購(gòu)自上海生工生物工程有限公司;SDS-PAGE 凝膠配制試劑盒、ClarityTMWestern ECL Substrate 顯色劑購(gòu)自碧云天生物技術(shù)公司;一抗Mouse anti-HA、二抗Goat anti-mouse IGg-HRP購(gòu)自北京全式金生物技術(shù)有限公司;PVDF膜購(gòu)自Bio-RAD公司;葡萄糖、瓊脂粉、醋酸鉀、α-萘酚、二乙酰、Na2CO3、NaHCO3、磷酸鹽、NaOH及無(wú)水甲醇等分析純?cè)噭┵?gòu)自國(guó)藥集團(tuán)化學(xué)試劑有限公司。
1.2.3 實(shí)驗(yàn)溶液
原生質(zhì)體溶液:K3PO3溶液濃度50 mmol/L,pH 7.5,山梨醇濃度1.4 mol/L,β-巰基乙醇濃度40 mmol/L。
TBST溶液:5 mol/L的NaCl 30 mL,1 mol/L的Tris·HCl(pH 8.0)10 mL,吐溫-20取0.5 mL,再加去離子水定容至1 L。
尿素溶液:尿素48.04 g,NaCl 0.585 g,1 mol/L的Tris·HCl(pH 8.0)取2.5 mL,加水定容至100 mL。
轉(zhuǎn)膜緩沖液:甘氨酸14.4 g,Tris 3.03 g,無(wú)水甲醇200 mL,加水定容至1 L。
5 g/L 脫脂牛奶:脫脂奶粉5 g,加TBST溶液定容至100 mL。
pH反應(yīng)緩沖液:pH 2~5,50 mmol/L醋酸鈉溶液;pH 6~9,50 mmol/L KH2PO4/K2HPO4溶液;pH 10~12,50 mmol/L Na2HPO4/Na3PO4。
1.3 實(shí)驗(yàn)方法
1.3.1 肌酐酶質(zhì)粒在釀酒酵母菌株中的重組轉(zhuǎn)化
肌酐酶質(zhì)粒pRS426-TEFpr-ss-CA-3HA由孔軍提供[11]。將該質(zhì)粒分別轉(zhuǎn)入野生型(wt)、dit1Δ、chs3Δ、osw2Δ及kre1Δ菌株中,然后將轉(zhuǎn)化細(xì)胞液涂到尿嘧啶缺陷型培養(yǎng)基平板上進(jìn)行篩選,長(zhǎng)出來(lái)的單菌落就是陽(yáng)性重組轉(zhuǎn)化子。同時(shí)也將空載pRS426-TEF轉(zhuǎn)到野生型中作為對(duì)照組。
1.3.2 重組酵母菌株的生長(zhǎng)、產(chǎn)孢及孢子純化
將陽(yáng)性重組轉(zhuǎn)化子接種到5 mL的尿嘧啶缺陷型培養(yǎng)基中,30 ℃搖床過(guò)夜培養(yǎng),取1 mL培養(yǎng)液轉(zhuǎn)接到80 mL的YPAce培養(yǎng)基中,30 ℃搖床培養(yǎng)24 h,4 000g離心1 min收集細(xì)胞,用無(wú)菌水洗1遍后將細(xì)胞重懸于80 mL的產(chǎn)孢培養(yǎng)基中,30 ℃搖床培養(yǎng)24 h后鏡檢產(chǎn)孢率。當(dāng)產(chǎn)孢率大于90%時(shí),4 000g離心1 min收集孢子。由于成熟的孢子被母細(xì)胞子囊殼包裹,因此必需先用細(xì)胞壁裂解酶(lyticase)對(duì)子囊殼進(jìn)行裂解,再超聲破碎,使孢子從子囊殼中釋放出來(lái),然后用Percoll密度梯度離心方法分離純化孢子[12-16],將分離后的孢子進(jìn)行真空冷凍干燥備用。
1.3.3 蛋白質(zhì)免疫印跡分析肌酐酶在酵母孢子中的表達(dá)
將含有肌酐酶(CA-3HA)的菌株及對(duì)照用產(chǎn)孢培養(yǎng)基產(chǎn)孢后,收集孢子。然后將孢子重懸于500 μL的含有蛋白酶抑制劑的8 mol/L尿素溶液中,加入適量玻璃珠。按振蕩1 min、冰上1 min的循環(huán)破碎孢子,振蕩破碎40 min后,9 000g離心10 min,取上清液,用Nano Drop 2000分光光度計(jì)測(cè)定蛋白質(zhì)濃度。然后統(tǒng)一上樣50 μg蛋白進(jìn)行SDS-PAG電泳(5%的濃縮膠,10%的分離膠),電泳結(jié)束后,從下到上按濾紙、PVDF膜、凝膠、濾紙的疊放順序放入電轉(zhuǎn)儀 (25 V,1.0 A,30 min) 內(nèi)進(jìn)行轉(zhuǎn)膜,轉(zhuǎn)膜完畢后用50 g/L的脫脂牛奶室溫封閉PVDF膜2 h,一抗(mouse anti-HA,1∶5 000稀釋)室溫孵育2 h,TBST溶液洗3次(10 min/次),二抗(goat anti-mouse IgG HRP,1∶6 000稀釋)室溫孵育2 h,TBST溶液洗3次后,用ECL顯色試劑盒對(duì)PVDF膜顯色,再用Image Quant LAS4000 進(jìn)行拍照。
1.3.4 肌酐酶酶活檢測(cè)
肌酐酶酶活的測(cè)定方法參考KIKKOMAN (Kikkoman, Noda Japan) C1-E肌酐酶活力測(cè)定方法。原理是通過(guò)測(cè)定肌酸產(chǎn)量來(lái)檢測(cè),即肌酐被肌酐酶降解后,產(chǎn)生肌酸,在堿性條件和α-萘酚的催化下,肌酸的胍基與二乙酰生成紫紅色物質(zhì),該物質(zhì)在OD525處有最大吸收峰。然后再根據(jù)OD525值來(lái)計(jì)算酶活。單位肌酐酶活定義為:在37 ℃和pH 6.8的反應(yīng)液中,水解生成1 μmol肌酸所需的肌酐酶的量即為1 U。所有的酶活測(cè)試都是用5 mg冷凍干燥孢子或50 μL (1.4 U) 的游離酶加入到反應(yīng)液中,37 ℃,反應(yīng)10 min后加入終止液并置于冰上,加入顯色液后測(cè)量OD525值。
為了測(cè)定溫度對(duì)肌酐酶活的影響,本研究測(cè)定了游離酶及孢子微膠囊酶反應(yīng)液(pH 6.5)在不同溫度下(30~90 ℃)的酶活。為了測(cè)定pH對(duì)肌酐酶活的影響,本研究測(cè)定了不同pH反應(yīng)緩沖液在60 ℃下反應(yīng)的酶活。為了檢測(cè)孢子微膠囊肌酐酶的可重復(fù)利用性,統(tǒng)一稱(chēng)取5 mg冷凍干燥的孢子進(jìn)行反應(yīng),首次反應(yīng)結(jié)束后離心收集孢子,測(cè)定上清中的酶活,用含有0.5% Triton X-100的0.6 mol/L NaCl溶液洗滌孢子1遍,然后再次測(cè)量孢子的肌酐酶活,如此重復(fù)操作4次。
1.3.5 孢子微膠囊肌酐酶的抗逆性研究
為了檢測(cè)肌酐酶抗蛋白酶K的性能,統(tǒng)一稱(chēng)取5 mg冷凍干燥孢子或20 μL游離酶,加入100 μL蛋白酶K(終濃度為30 U/mL)緩沖液(50 mmol/L Tris-HCl pH 7.5,10 mmol/L CaCl2),37 ℃孵育12 h。然后用含有0.5% Triton X-100的0.6 mol/L NaCl溶液洗滌孢子2遍,再測(cè)定酶活。為了檢測(cè)肌酐酶抗SDS的性能,統(tǒng)一稱(chēng)取5 mg冷凍干燥孢子或20 μL游離酶,加入100 μL 5% SDS,30 ℃孵育10 min。然后用含有0.5% Triton X-100的0.6 mol/L NaCl溶液洗滌孢子2遍,再測(cè)定酶活。
1.3.6 數(shù)據(jù)統(tǒng)計(jì)分析
統(tǒng)計(jì)數(shù)據(jù)為3次獨(dú)立重復(fù)實(shí)驗(yàn)的平均值,數(shù)據(jù)分析用Microsoft Excel軟件處理。
2 結(jié)果與討論
2.1 肌酐酶在酵母孢子中的表達(dá)
為了檢測(cè)肌酐酶在各突變株孢子內(nèi)的表達(dá)水平差異,需要進(jìn)行蛋白免疫印記(Western blot)分析且上樣量一致。從圖2中可知,除對(duì)照組沒(méi)有條帶外,其他孢子中都有條帶,但不同孢子中肌酐酶的表達(dá)水平有明顯差異,也即不同突變株孢子包埋固定肌酐酶的能力有差異。由于野生型、osw2Δ、kre1Δ孢子都有二酪氨酸層,所以三者固定肌酐酶的量基本一致。dit1Δ孢子雖無(wú)二酪氨酸層,但有殼聚糖層能吸附部分肌酐酶,因此其固定肌酐酶的量明顯少于野生型、osw2Δ、kre1Δ孢子,而chs3Δ孢子缺少殼聚糖層和二酪氨酸層,不能吸附、阻止肌酐酶外泄,所以chs3Δ孢子基本不能固定肌酐酶。

圖2 蛋白免疫印跡分析肌酐酶在酵母孢子中的表達(dá)Fig.2 Western blot analysis of creatininase expression level in yeast spores
為了驗(yàn)證該推論是否正確,分別對(duì)野生型、chs3Δ孢子及對(duì)照組中的培養(yǎng)液進(jìn)行酶活測(cè)定(見(jiàn)圖3),發(fā)現(xiàn)chs3Δ孢子培養(yǎng)液中的肌酐酶活性顯著高于野生型孢子的培養(yǎng)液,而野生型孢子培養(yǎng)液的酶活明顯高于對(duì)照組,是因?yàn)殒咦颖谥袣ぞ厶菍雍投野彼釋拥男纬尚枰欢螘r(shí)間,在孢子壁沒(méi)有完全組裝形成之前,是無(wú)法阻止肌酐酶外泄的,所以野生型孢子的培養(yǎng)液也有酶活。該結(jié)果說(shuō)明,孢子壁中的殼聚糖層和二酪氨酸層是影響孢子固定肌酐酶能力的主要因子,特別是二酪氨酸層對(duì)孢子固定肌酐酶的能力有重要影響。同時(shí),該結(jié)果也說(shuō)明kre1Δ孢子固定肌酐酶的能力與野生型、osw2Δ孢子相同。
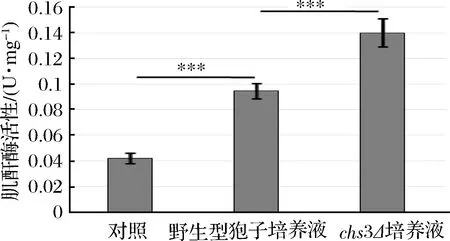
圖3 不同突變體孢子培養(yǎng)液中肌酐酶活性Fig.3 Activity of creatininase in the cultureof different mutant spores (***p<0.001)
2.2 孢子中肌酐酶的酶活比較
為了檢測(cè)肌酐酶在孢子內(nèi)是否有活性,需要進(jìn)行酶活檢測(cè)(圖4)。圖4-A的結(jié)果表明,kre1Δ孢子與osw2Δ孢子的肌酐酶活最高,dit1Δ孢子的酶活略低于kre1Δ、osw2Δ,這三者間的酶活無(wú)明顯差異,但都顯著高于野生型孢子與chs3Δ孢子的肌酐酶活,而不含肌酐酶的對(duì)照組酶活最低,顯著低于野生型孢子。圖4-A與圖2的結(jié)果略有不同,圖2中野生型孢子包埋固定肌酐酶的量明顯多于dit1Δ孢子,但酶活卻顯著低于dit1Δ孢子,原因是野生型孢子中的二酪氨酸層完整嚴(yán)密,所以能包埋固定較多的肌酐酶,但同時(shí)也阻礙了孢外底物與酶充分接觸,因此酶活較低。

A-不同突變體孢子酶活;B-野生型完整孢子與野生型破碎孢子酶活圖4 孢子微膠囊固定肌酐酶活性Fig.4 Activity of spore-encapsulated creatininase (*p<0.05;***p<0.001)
為了驗(yàn)證該假設(shè)是否正確,取等量的野生型孢子兩份,1份進(jìn)行破碎處理,另1份不破碎,然后對(duì)這兩份樣品中的肌酐酶活進(jìn)行測(cè)定。從圖4-B中可看出,野生型破碎孢子中的酶活明顯高于野生型完整孢子中的酶活,說(shuō)明野生型孢子中的二酪氨酸層確實(shí)會(huì)阻礙底物與酶的接觸反應(yīng),從而影響其酶活。而dit1Δ孢子雖無(wú)二酪氨酸層,但還有殼聚糖層能吸附固定部分肌酐酶,所以其固定酶的能力有所減弱,但由于沒(méi)有二酪氨酸層的阻擋,孢外底物可充分與殼聚糖層上的肌酐酶接觸反應(yīng),所以dit1Δ孢子的酶活反而比野生型孢子的酶活高(圖4-A)。而kre1Δ與osw2Δ雖有二酪氨酸層,但二者的孢子壁可能出現(xiàn)一定程度的松散,增加了孢子壁的通透性。這樣既能包埋固定較多的酶,同時(shí)孢外底物也能通過(guò)二酪氨酸層與酶接觸反應(yīng),所以酶活較高。而chs3Δ孢子沒(méi)有二酪氨酸層和殼聚糖層,基本不能包埋固定肌酐酶,所以酶活較低。該結(jié)果表明kre1Δ孢子與osw2Δ孢子一樣,不僅能固定較多的酶,而且酶活也顯著高于野生型孢子。
2.3 孢子中肌酐酶的可重復(fù)利用性分析
由于野生型孢子與kre1Δ、osw2Δ孢子都能固定較多的酶,而dit1Δ孢子雖然固定酶的能力有所減弱,但酶活較高。因此以可重復(fù)利用性為標(biāo)準(zhǔn)來(lái)進(jìn)一步確認(rèn)孢子微膠囊肌酐酶的穩(wěn)定性。從圖5中可看出,在重復(fù)利用5次后,野生型孢子的肌酐酶相對(duì)酶活下降最慢,其殘留肌酐酶的相對(duì)酶活還有87.36%,其次為kre1Δ,其相對(duì)酶活還有85.13%,而后是osw2Δ,其相對(duì)酶活還有78.86%,而dit1Δ孢子肌酐酶的相對(duì)酶活下降最快,其殘留肌酐酶的相對(duì)酶活只有56.97%。該結(jié)果表明kre1Δ與osw2Δ孢子一樣都有較好的可重復(fù)利用性,同時(shí)酶活也高。
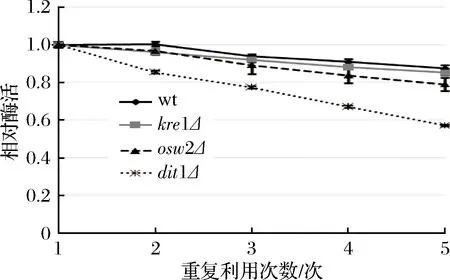
圖5 孢子微膠囊肌酐酶的可重復(fù)利用性分析Fig.5 The reusability of encapsulated creatininase
2.4 孢子肌酐酶的抗逆性分析
為了研究孢子微膠囊肌酐酶是否具有抗逆性,分別利用蛋白酶K和5%的SDS對(duì)孢子微膠囊酶及游離酶進(jìn)行處理,然后檢測(cè)肌酐酶活,并與未經(jīng)蛋白酶K及SDS處理的酶活進(jìn)行比較。從圖6中可看出,經(jīng)蛋白酶K處理后,野生型、kre1Δ、osw2Δ及dit1Δ孢子的相對(duì)酶活分別為96.47%,86.81%,83.02%,73.26%;經(jīng)5%的SDS處理后(圖7),野生型、kre1Δ、osw2Δ及dit1Δ孢子的相對(duì)酶活分別為92.63%,84.63%,82.25%,66.36%。而游離肌酐酶的抗逆性較弱,經(jīng)蛋白酶K及SDS處理后,其相對(duì)酶活分別為40.65%,41.02%。該結(jié)果表明孢子固定的肌酐酶都表現(xiàn)出較強(qiáng)的抗蛋白酶K及抗SDS能力,且孢子微膠囊肌酐酶的抗逆性顯著高于游離酶。同時(shí),該結(jié)果也說(shuō)明不同孢子壁缺陷的孢子,其抗逆性也有差異,因dit1Δ孢子沒(méi)有二酪氨酸層的保護(hù),所以抗逆性不如有二酪氨酸層的孢子,而kre1Δ孢子與野生型、osw2Δ孢子一樣都有較強(qiáng)的抗逆性。
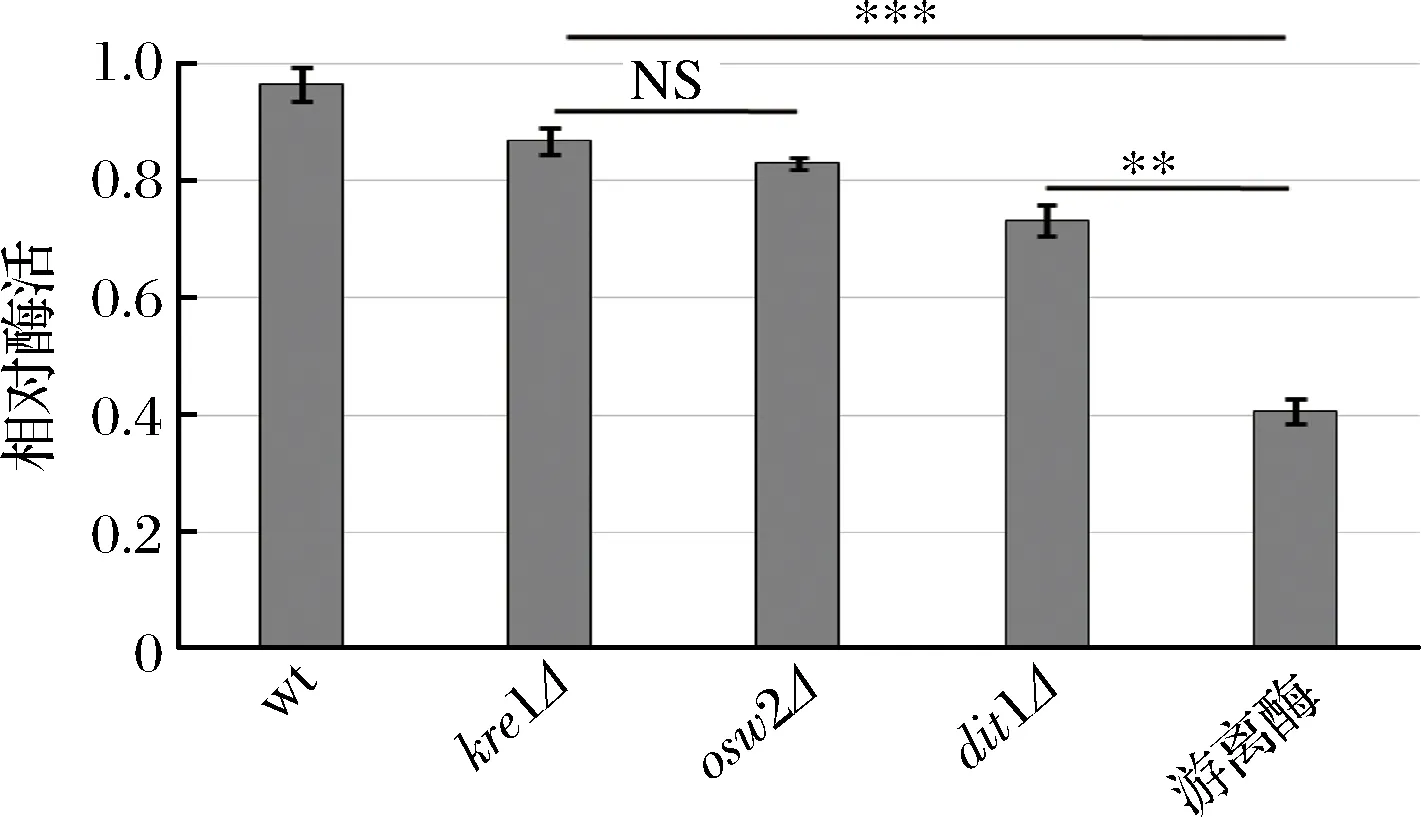
圖6 孢子微膠囊肌酐酶抗蛋白酶K能力分析Fig.6 Stability of encapsulated creatininase against proteinase K treatment (NS,不顯著**p<0.01;***p<0.001)
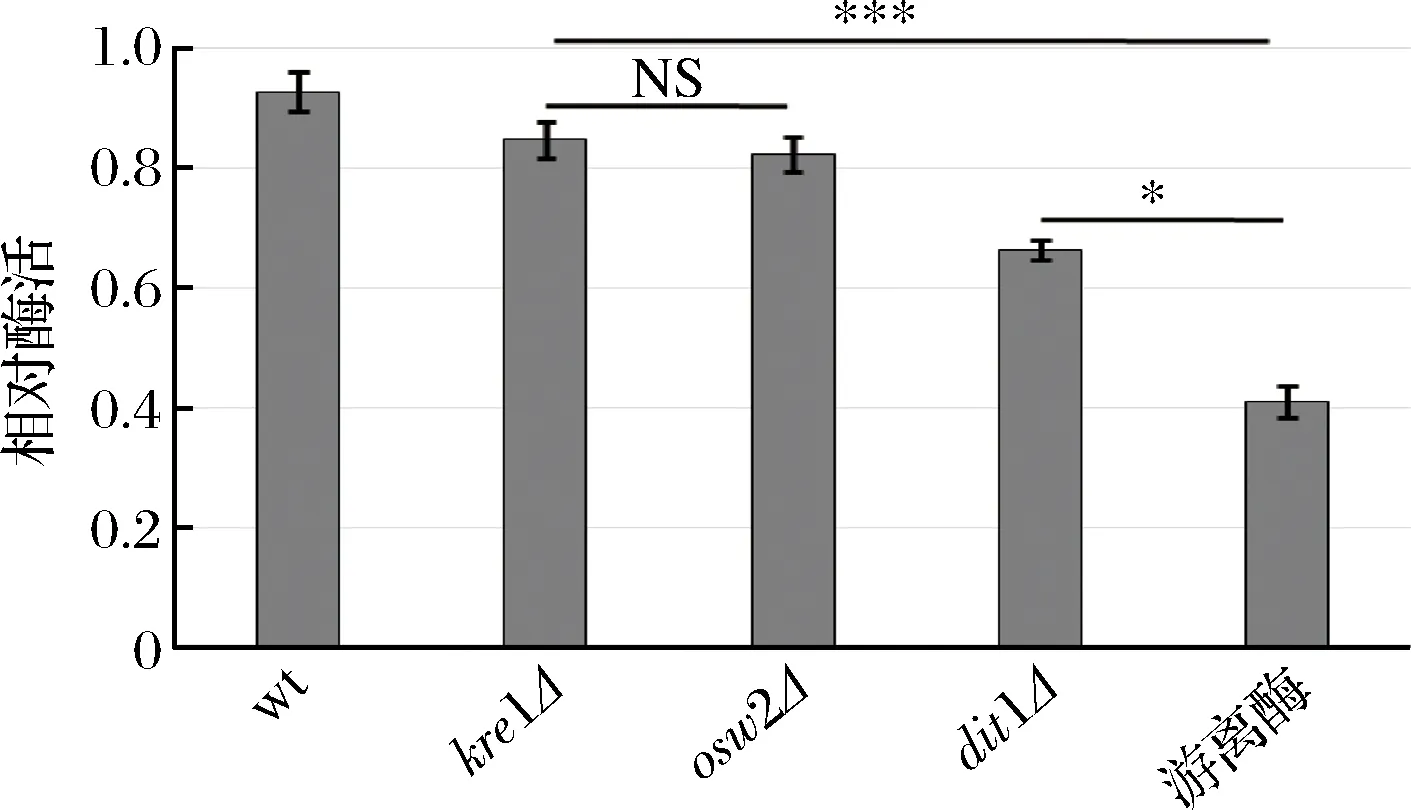
圖7 孢子微膠囊肌酐酶抗SDS能力分析Fig.7 Stability of encapsulated creatininase against SDS treatment (NS,不顯著*p<0.05;***p<0.001)
2.5 溫度和pH對(duì)肌酐酶的影響
綜合上述結(jié)果可知,dit1Δ孢子在可重復(fù)利用性及抗逆性等方面都遜色于其他類(lèi)型孢子,因此為了研究不同溫度和pH對(duì)孢子微膠囊肌酐酶的影響,只對(duì)野生型、kre1Δ、osw2Δ及對(duì)照組游離酶進(jìn)行分析。從圖8中可看出,孢子微膠囊肌酐酶和游離肌酐酶的酶活最適溫度都為60 ℃,當(dāng)溫度高于60 ℃以后,游離肌酐酶的酶活逐漸下降,在90 ℃高溫時(shí),游離酶的相對(duì)酶活只有60%左右。與游離肌酐酶相比,經(jīng)孢子固定的肌酐酶在不同溫度下都表現(xiàn)出較好的穩(wěn)定性,即使在90 ℃高溫條件下,孢子固定的肌酐酶相對(duì)酶活依然維持在80%以上,說(shuō)明孢子固定的肌酐酶具有較好的耐高溫性。在不同pH條件下反應(yīng)時(shí),孢子固定的肌酐酶和游離肌酐酶的相對(duì)酶活最適pH均為6.0 (圖9)。當(dāng)大于pH 7.0,游離肌酐酶的相對(duì)酶活隨著pH值升高而迅速下降,當(dāng)pH 12.0時(shí),游離酶的相對(duì)酶活降到40%以下,而孢子固定的肌酐酶相對(duì)酶活雖然也有所下降,但依然維持在80%以上,說(shuō)明孢子固定的肌酐酶表現(xiàn)出較廣范圍的pH耐受性。同時(shí),該結(jié)果也表明kre1Δ孢子與野生型、osw2Δ孢子一樣,都有較好的耐高溫性及較廣范圍的pH耐受性。
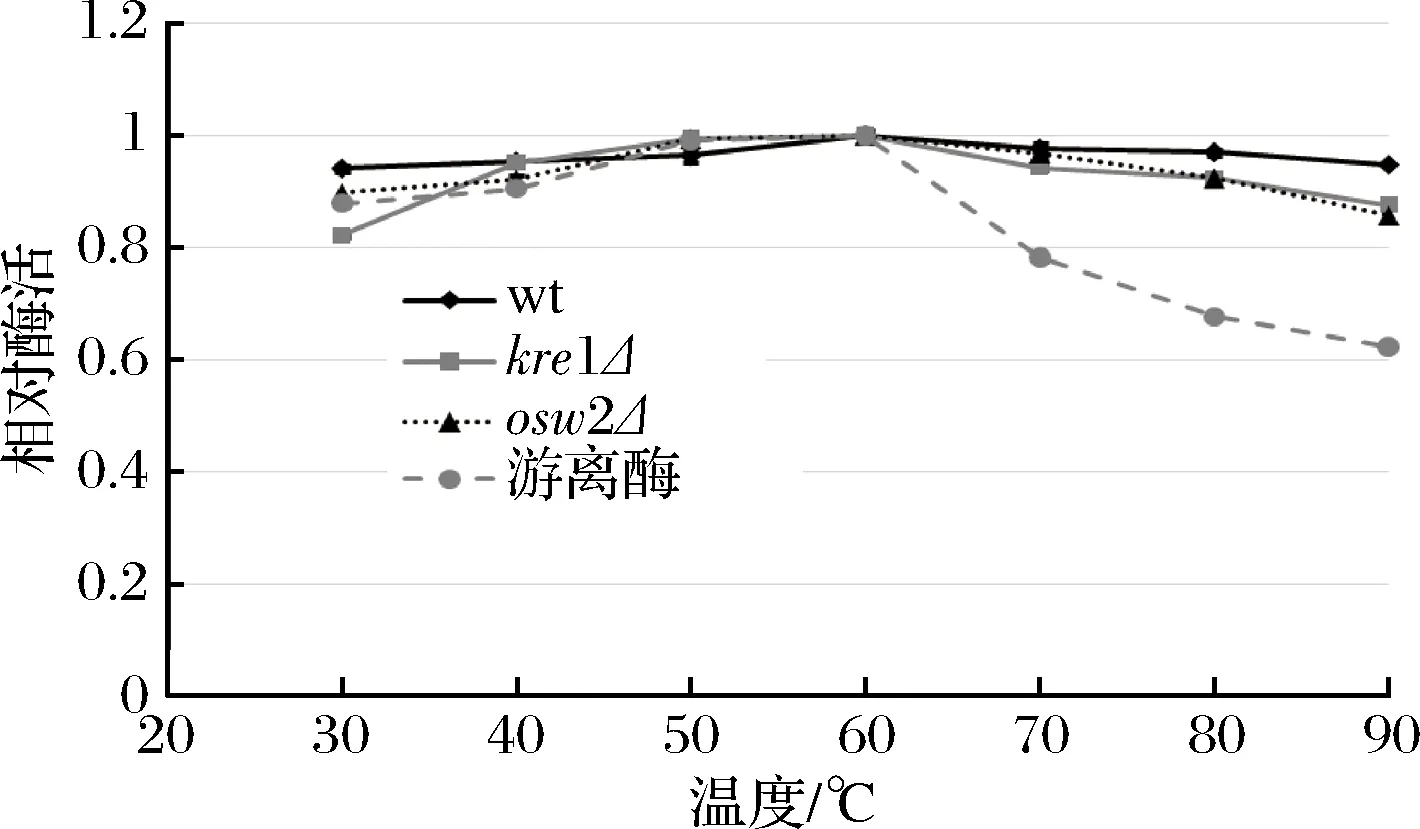
圖8 溫度對(duì)孢子微膠囊肌酐酶活的影響Fig.8 The effect of temperature on activity of the spore- encapsulated creatininase
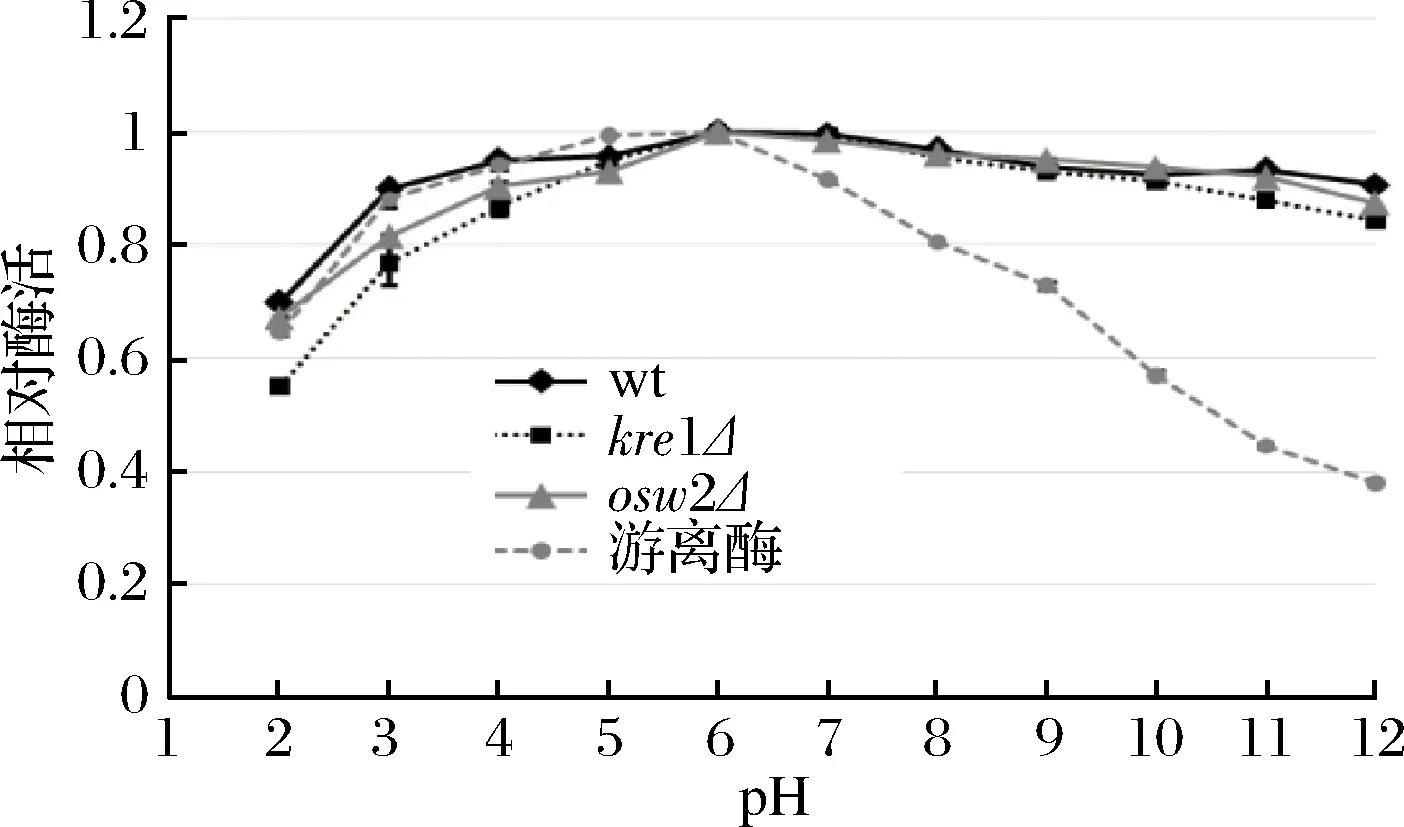
圖9 pH對(duì)孢子微膠囊肌酐酶活的影響Fig.9 The effect of pH on activity of the spore- encapsulated creatininase
3 結(jié)論
利用不同孢子壁缺陷的酵母孢子對(duì)肌酐酶進(jìn)行固定,并對(duì)孢子微膠囊肌酐酶的酶學(xué)性能進(jìn)行了分析。雖然KRE1與OSW2功能不同,但kre1Δ孢子固定的肌酐酶酶學(xué)性能與osw2Δ孢子固定的肌酐酶學(xué)性能相近,二者酶活都顯著高于野生型孢子。因游離肌酐酶沒(méi)有任何保護(hù),直接暴露在環(huán)境中,所以不能重復(fù)利用且抗逆性較弱,其酶活受外界的溫度、pH等因素的影響較大,而孢子固定的肌酐酶由于有孢子壁,特別是二酪氨酸層的保護(hù),表現(xiàn)出較好的可重復(fù)利用性,不僅抗逆性強(qiáng)而且具有較強(qiáng)的耐高溫性及較廣范圍的pH耐受性。因此,kre1Δ孢子也可作為優(yōu)良的酶固定化載體在以后的研究中加以應(yīng)用。
肌酐酶也稱(chēng)肌酐酰胺水解酶,可將肌酐催化生成肌酸,肌酸在堿性條件和α-萘酚的催化下可生成紫紅色物質(zhì),該物質(zhì)在OD525處有最大吸收峰。因肌酐在很多腎臟類(lèi)疾病中都有重要的診斷指示作用[17],但游離的肌酐酶在強(qiáng)酸、強(qiáng)堿和高溫等條件下不穩(wěn)定,同時(shí)也存在不易長(zhǎng)期貯存、無(wú)法重復(fù)利用及不易保持活力等缺點(diǎn),其應(yīng)用受到一定的限制。而固定化后的酶,有較高的穩(wěn)定性且能重復(fù)利用,可在一定程度上解決這些問(wèn)題。目前,固定化酶的方法主要有化學(xué)法和物理法兩大類(lèi),化學(xué)法包括交聯(lián)法和結(jié)合法;物理法包括吸附法與包埋法。但這些方法各有缺點(diǎn)[18],如交聯(lián)法反應(yīng)條件比較激烈,交聯(lián)過(guò)程需要酶分子參與化學(xué)反應(yīng),固定化后酶容易失活且不易回收;結(jié)合法反應(yīng)條件較苛刻,操作工藝復(fù)雜且固定化后酶活力有所下降;吸附法固定的酶與載體的結(jié)合力較弱,容易受外界因素影響而脫離;包埋法則易受包埋孔徑的限制,大分子底物難與被包埋的酶分子接觸,只適合小分子催化底物的反應(yīng)。酵母孢子微膠囊酶固定法是一種新型的生物固定方法。該方法的優(yōu)勢(shì)是直接在酵母孢子內(nèi)表達(dá)酶,無(wú)需提純酶,即可直接應(yīng)用;生產(chǎn)工藝簡(jiǎn)單,不改變酶分子結(jié)構(gòu),固定化后酶活較高;可常溫長(zhǎng)期儲(chǔ)存,而且具有較好的抗逆性及可重復(fù)利用性;同時(shí)還可利用分子生物學(xué)手段改變孢子壁通透性,提高酶活。鑒于游離肌酐酶存在諸多應(yīng)用限制,而用酵母孢子微膠囊肌酐酶固定法可有效克服這些困難。因此,有望用酵母孢子建立一種快速方便的肌酐酶臨床診斷方法,本研究為該方法的建立提供了一定的理論依據(jù)。
[1] NEIMAN A M. Ascospore formation in the yeastSaccharomycescerevisiae[J]. Microbiology & Molecular Biology Reviews Mmbr, 2005, 69(4): 565-584.
[2] RIJ K V. Electron microscopy of germinating ascospores ofSaccharomycescerevisiae[J]. Archives of Microbiology, 1978, 117(1): 73-77.
[3] TACHIKAWA H, BLOECHER A, TATCHELL K, et al. A Gip1p-Glc7p phosphatase complex regulates septin organization and spore wall formation [J]. Journal of Cell Biology, 2001, 155(5): 797-808.
[4] COLUCCIO A, BOGENGRUBER E, CONRAD M N, et al. Morphogenetic pathway of spore wall assembly inSaccharomycescerevisiae[J]. Eukaryotic Cell, 2004, 3(6): 1 464-1 475.
[5] LESAGE G, BUSSEY H. Cell wall assembly inSaccharomycescerevisiae[J]. Microbiol Mol Biol Rev, 2006, 70(2): 317-343.
[6] PAMMER M, BRIZA P, ELLINGER A, et al.DIT101 (CSD2,CAL1), a cell cycle-regulated yeast gene required for synthesis of chitin in cell walls and chitosan in spore walls [J]. Yeast, 1992, 8(12): 1 089-1 099.
[7] BRIZA P, ECKERSTORFER M, BREITENBACH M. The sporulation-specific enzymes encoded by theDIT1 andDIT2 genes catalyze a two-step reaction leading to a solubleLL-dityrosine-containing precursor of the yeast spore wall [J]. Proceedings of the National Academy of Sciences, 1994, 91(10): 4 524-4 528.
[8] BRIZA P, ELLINGER A, WINKLER G, et al. Characterization of aDL-dityrosine-containing macromolecule from yeast ascospore walls [J]. Journal of Biological Chemistry, 1990, 265(25): 15 118-15 123.
[9] SUDA Y, RODRIGUEZ R K, COLUCCIO A E, et al. A screen for spore wall permeability mutants identifies a secreted protease required for proper spore wall assembly [J]. Plos One, 2009, 4(9): 1-10.
[10] KONG Jan, LI Zi-jie, ZHANG Hui-jie, et al. Production of encapsulated creatinase using yeast spores [J]. Bioengineered, 2016,8(4): 1-9.
[11] KONG Jun, LI Zi-jie, ZHANG Hui-jie, et al. Consecutive hydrolysis of creatinine using creatininase and creatinase encapsulated inSaccharomycescerevisiaespores [J]. Biotechnology Letters, 2017,39(2): 261-267.
[12] SHI L, LI Z, TACHIKAWA H, et al. Use of yeast spores for microencapsulation of enzymes [J]. Applied & Environmental Microbiology, 2014, 80(15): 4 502-4 510.
[13] PAN H P, WANG N, TACHIKAWA H, et al. ?-1,6-glucan synthesis-associated genes are required for proper spore wall formation inSaccharomycescerevisiae[J]. Yeast, 2017,34(1):431-446.
[14] NEIMAN A M, KATZ L, BRENNWALD P J. Identification of domains required for developmentally regulated SNARE function inSaccharomycescerevisiae[J]. Genetics(Print), 2000, 155(4): 1 643-1 655.
[15] ZHANG H, TACHIKAWA H, GAO X D, et al. Applied usage of yeast spores as chitosan beads [J]. Applied & Environmental Microbiology, 2014, 80(16): 5 098-5 105.
[16] 李毅, 李子杰, 中西秀樹(shù), 等. 木糖異構(gòu)酶釀酒酵母孢子“微膠囊”的構(gòu)建及酶學(xué)性質(zhì)分析 [J]. 食品與發(fā)酵工業(yè), 2015, 41(4): 6-11.
[17] 李萍, 趙瑩, 余霆. 肌酐在腎小球?yàn)V過(guò)功能損傷診斷中的價(jià)值的系統(tǒng)評(píng)價(jià) [J]. 中國(guó)循證醫(yī)學(xué)雜志, 2004, 4(11): 752-758.
[18] 隋穎. 生物酶固定化方法的研究新進(jìn)展 [J]. 山東化工, 2013, 42(8): 71-72.
猜你喜歡
河北科技師范學(xué)院學(xué)報(bào)(2022年2期)2022-08-26 08:55:40
河北科技師范學(xué)院學(xué)報(bào)(2021年1期)2021-05-10 03:34:20
中成藥(2017年12期)2018-01-19 02:06:57
電源技術(shù)(2017年1期)2017-03-20 13:37:59
廣西科技大學(xué)學(xué)報(bào)(2016年1期)2016-06-22 13:10:38
天然產(chǎn)物研究與開(kāi)發(fā)(2016年1期)2016-06-05 10:29:25
食品界(2016年4期)2016-02-27 07:36:46
中國(guó)果菜(2015年2期)2015-03-11 20:01:01
應(yīng)用化工(2014年7期)2014-08-09 09:20:21
應(yīng)用技術(shù)學(xué)報(bào)(2014年4期)2014-02-28 14:52:40
