北京典型樹種木質組織碳釋放速率溫度敏感性的時間變化規律和鉛錘分異特征
2018-03-08 07:43:34韓風森王曉琳
生態學報 2018年2期
關鍵詞:生長
韓風森 , 王曉琳 , 胡 聃
1 中國科學院生態環境研究中心,北京 100085 2 中國科學院大學,北京 100049 3 中國農業科學院煙草研究所, 青島 266101
木質組織碳排放(ECO2)是陸地生態系統碳平衡的重要組成部分[1- 3],分別占北方針葉林和溫帶闊葉林地上碳排放總量的25%—50%[4],因而對林業生產和群落碳收支起到至關重要的作用[5-8]。目前,關于ECO2的研究主要集中在自然生態系統中,對城市環境下人工群落中木質組織碳排放規律的報導相對欠缺。
在現有研究中,木質組織碳通量觀測多局限于樹干中的某一個點(通常為胸徑位置),測定其白天的CO2釋放速率[9-10],并基于這種非連續的位點測量,對個體或群落水平的ECO2進行估算[11- 13]。但越來越多的研究認為,ECO2本身的時間變化和鉛垂變異性對木質組織碳估算帶來很大誤差[14- 21]。季節性的生態因子變化和植物的生長代謝規律,是造成ECO2季節動態和晝夜變化的主要原因[18,22]。此外,溫度和活細胞含量在同一個體內的梯度變化,使ECO2在個體內存在明顯的鉛錘分異特征[14,23]。準確量化ECO2的時間變化規律和空間分異特征,成為準確估算木質組織碳排放的前提條件。
在所有生物和非生物因子中,溫度被認為是調控木質組織碳排放速率的關鍵因素[4,8,22,24- 27]。通常情況下,木質組織ECO2隨溫度呈指數增長,ECO2對溫度的響應程度用溫度敏感系數Q10(即溫度每升高10℃,ECO2變化的倍數)來表示[1,2,14,19,28-29],在絕大多數生態系統模型中被設為常數2.0[28,30]。但近年來,有研究發現Q10在不同樹種之間存在較大差異[31- 34],例如日本扁柏和北美短葉松的Q10值變化區間為1.2—3.2[22,35],而白楊和挪威云杉則集中分布在1.0—1.7和1.9—2.6之間[4,15]。據最新報道,絕大部分樹種的Q10范圍在1.0—6.0之間變化,平均數值約為2.0[36]。除種間差異外,Q10在同樹種內或某一個體內部也存在極大的變異性[16,37- 39]。無論Q10的種間差異還是種內變化,均隨測量季節或測量溫度的改變而發生變化[15,18]。
為深入探究木質組織碳排放規律及其影響因素,本研究選取北京市4個典型樹種(國槐、旱柳、華北落葉松和側柏)作為研究對象,對每個樹種不同高度上的木質組織碳排放規律及其氣象因子和形態指標做了連續觀測。旨在:(1)比較不同樹種之間的碳排放差異及其時空動態;(2)探究木質組織碳排放對溫度的響應在不同時間尺度下的變化規律,以及特定時間內不同高度上的分異特征。
1 材料和方法
1.1 研究區概況
研究區位于北京市海淀區東升八家郊野公園(40.03°N,116.33°E),屬于典型的溫帶半濕潤季風型大陸性氣候,夏季炎熱多雨,冬季寒冷干燥,2014年年平均氣溫為13.4℃,年降水量為590—600 mm,降水時間多集中于夏季。該樣地占地面積101.45 hm2,森林覆蓋率達到70%以上,林分密度約為870株/hm2,平均樹高和胸徑分別為(8.5±0.8) m和(11.3±1.4) cm(2013年調查數據)。該樣地內闊葉樹種主要是國槐(Sophorajaponica)和旱柳(Salixmatsudana),針葉樹種以華北落葉松(Larixprincipis-rupprechtii)和側柏為主(Platycladusorientalis),4個樹種占該人工群落地上生物量的80%以上。其中,闊葉樹種在4月份出現葉片,9月份開始落葉;針葉樹種全年長有葉片,但木質組織的徑向生長主要集中在5—9月。
1.2 樣地設置和試驗方法
按群落內個體徑級分布,采用分層抽樣的方法選取樣地內生長良好的國槐、旱柳、華北落葉松和側柏各3株作為本次實驗的研究對象。于2014年1月至12月期間,逐月觀測其ECO2排放規律,以及相應的氣象因子(大氣溫度、濕度、光照和樹干內部溫度)和形態指標(徑向生長速率)。實驗所選取的12株樣樹高度和胸徑范圍分別為8.37—16.4 m和16.5—31.5 cm(表1)。
1.2.1樣點設置和ECO2測定
在每株樣樹上,我們選擇4個高度作為本次研究的觀測樣點,即10、130、240、330 cm,分別對應樹干的基部、胸高、頂部及一級分枝位置。木質組織碳通量測定設定在2014年1—12月的中上旬進行,晝夜連續觀測木質組織碳排放速率(以單位面積表示)。木質組織ECO2測定裝置采用LI- 8100土壤碳通量測量系統(LI-COR, Lincoln, USA)及自制呼吸環(Polyvinyl Chloride, 60 P)。在測量模式下,非色散紅外氣體分析儀(IRGA)用于監測密閉氣室內的CO2濃度變化,然后通過數據記錄器擬合氣室內CO2濃度隨時間的線性變化,進而計算出木質組織表面的碳排放速率。每次測量前,都要檢查呼吸氣室的氣密性,確保呼吸環不漏氣。測量時,控制空氣流速為0.5 L/min,測量結束后及時取走呼吸氣室,以防影響樹木的正常生長和代謝。為避免太陽輻射,所有樣點選擇在樹干的背光面(即北面)。

表1 樣樹基本特征
a:測量高度, heights for measurement (cm);D:測點直徑,diameters at measurement point (cm);SVS:單位面積內的邊材體積,sapwood volume per unit of stem surface (cm3/cm2)
1.2.2 氣象因子和徑向生長測定
木質組織ECO2測定期間,同步測定大氣溫度、濕度及樹干溫度。空氣溫度和相對濕度的測定采用美國Onset公司生產的HOBO Pro V2 U23-002溫濕度探頭(Onset Computer Corporation, Bourne, MA, USA),該探頭的溫度精度及分辨率分別為±0.21℃和0.02℃(適用范圍0和50℃之間),濕度精度及分辨率分別為±2.5%和0.1%(適用范圍10%和90%之間)。枝干溫度測定采用同公司生產的HOBO Pro V2 U23-004溫度探頭,其精度和分辨率與U23-002相同。大氣溫濕度及枝干溫度的測量時間間隔均為2 min。枝干徑向生長速率采用測樹器進行測量(CR10X, Campbell Scientific, Logan, UT),月末與月初的直徑差值代表當月的徑向生長量。
1.3 溫度敏感性系數Q10的計算
木質組織碳排放對溫度的敏感性系數,通過ECO2與木質組織溫度(TW)的指數擬合方程來計算[24]:
ECO2=E0×eKTW
(1)
其中,ECO2和E0分別為溫度TW和0℃下木質組織的碳通量速率(μmol m-2s-1),K為ECO2和TW的擬合參數(表征ECO2對溫度的敏感程度)。相應地,溫度敏感系數Q10計算為:
Q10=e10K
(2)
在比較Q10的季節變化時,將每個樹種在當前月份內的ECO2與TW進行單獨擬合,并計算出每個樹種在當前月份內的Q10值;在對Q10的日不對稱性分析中,根據樹干溫度與大氣溫度的相關關系將一天之內劃分4個時段,并將每個時段內的ECO2與TW進行單獨擬合,計算出該時段內的Q10值。
1.4 數據處理
不同數據組之間的差異性比較,采用SPSS 19.0中的單因素方差(one-way ANOVA)和最小顯著差異法分析(LSD)。分析溫度和徑向生長對木質組織碳通量的季節變化影響時,選擇溫度和徑向生長為自變量,木質組織碳通量速率為因變量做多重線性回歸分析,利用各自變量與因變量之間的偏相關系數的絕對值來判斷溫度和徑向生長對木質組織碳通量的影響程度。
2 結果和分析
2.1 ECO2的季節動態和影響因子分析
實驗期間,4個樹種在單位面積內的木質組織表面CO2通量速率(ECO2)均表現為單峰型季節變化節律,從1月份開始,ECO2(μmol m-2s-1)隨枝干溫度(TW)的增加而增加,到7月(旱柳和側柏)、8月份(國槐和華北落葉松)達到最高,之后緩慢下降,12月降為最低,與TW季節變化格局基本一致(P<0.05)。但在不同樹種間,枝干ECO2存在明顯差異,闊葉樹種(國槐、旱柳)的年均ECO2水平(國槐和旱柳分別為(1.89±0.57) μmol m-2s-1和(1.72±0.44) μmol m-2s-1)顯著高于針葉樹種(華北落葉松和側柏分別為(1.50±0.45) μmol m-2s-1和(1.41±0.46) μmol m-2s-1)(P<0.05),且夏季尤為明顯(圖1)。
為排除溫度對ECO2的影響,對所有月份內ECO2進行溫度歸一化處理(用E0表示)后發現,E0仍存在明顯的季節變化,并且與木質組織徑向生長模式基本一致。為進一步分析枝干ECO2季節變化的影響因素,本研究選取組織溫度、徑向生長速率及其組合為自變量,ECO2為因變量做線性回歸分析,發現加入溫度、徑向生長及其組合變量后,其擬合程度明顯高于單一變量。這一結果表明,溫度和木質組織的徑向生長共同影響ECO2的季節變化。通過對兩因子的偏相關分析發現,在溫度和徑向生長中,ECO2對溫度的響應更加敏感(圖1,表2)。
2.2 ECO2的溫度敏感性(Q10)
2.2.1 季節變化和鉛錘梯度分布
對于本研究所選取的4個樹種,溫度是影響ECO2季節變化的主導因素。但對于不同的樹種而言,ECO2對溫度的敏感程度存在顯著差異(圖2)。本研究中,國槐、旱柳、華北落葉松和側柏的年均Q10值分別為2.32,2.09,2.57和2.49,針葉樹種的溫度敏感性系數(Q10)顯著高于闊葉樹種。
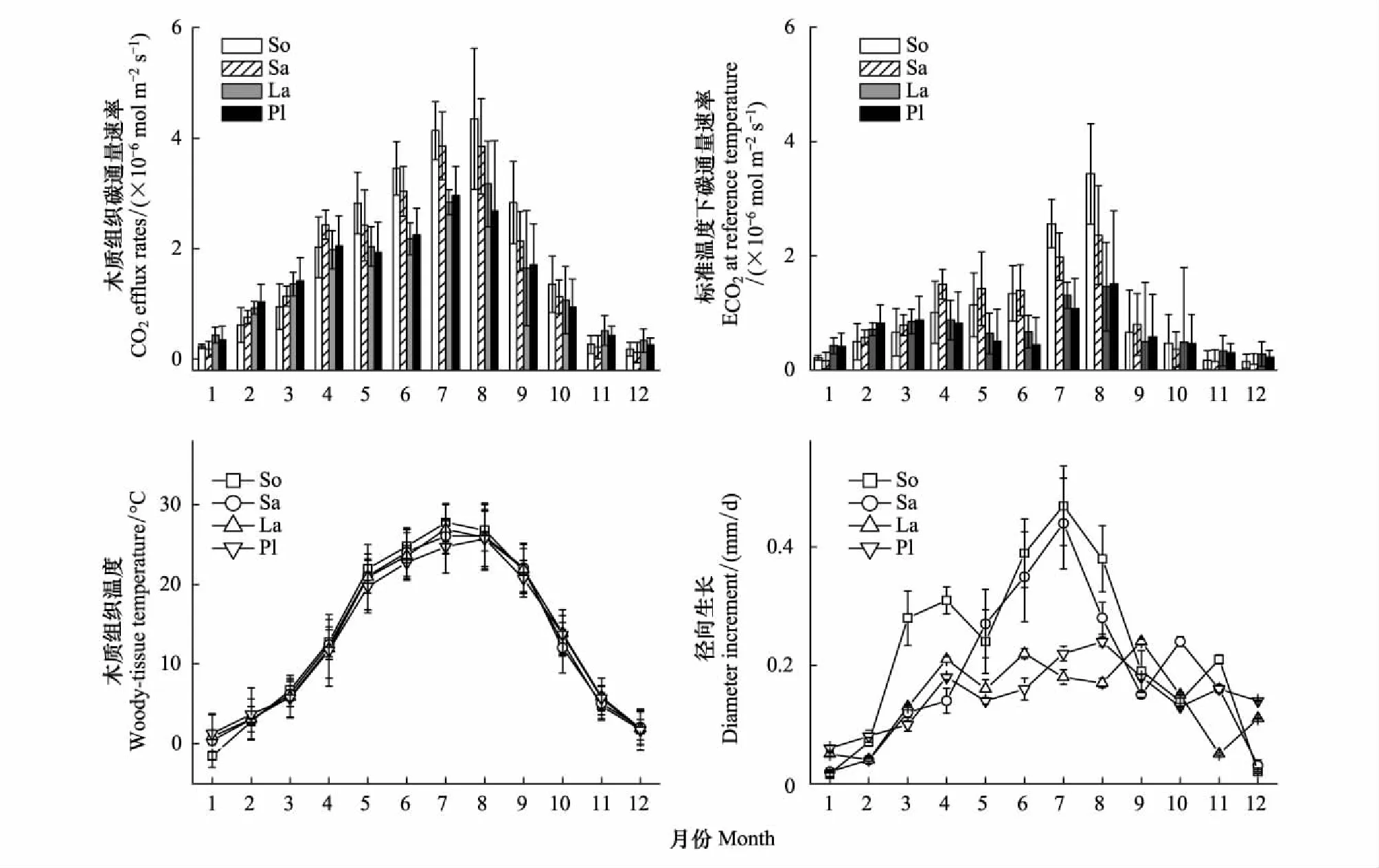
圖1 枝干呼吸速率ECO2、溫度歸一化后的(0℃)的ECO2、枝干溫度(TW)和徑向增長速率的季節變異Fig.1 Seasonal variations of CO2 efflux rates, CO2 efflux rates at the reference temperature 0℃, woody-tissue temperature and monthly diameter increment for 4 tree speciesSo: 國槐, Sophora japonica; Sa: 旱柳, Salix matsudana; La: 華北落葉松, Larix principis-rupprechtii; Pl: 側柏, Platycladus orientalis圖中數據為平均值±標準差,平均值為樹種內3個樣本在4個高度上的月平均值,誤差棒代表各指標在不同個體間的變異程度,用均值的一倍方差表示(n=12)
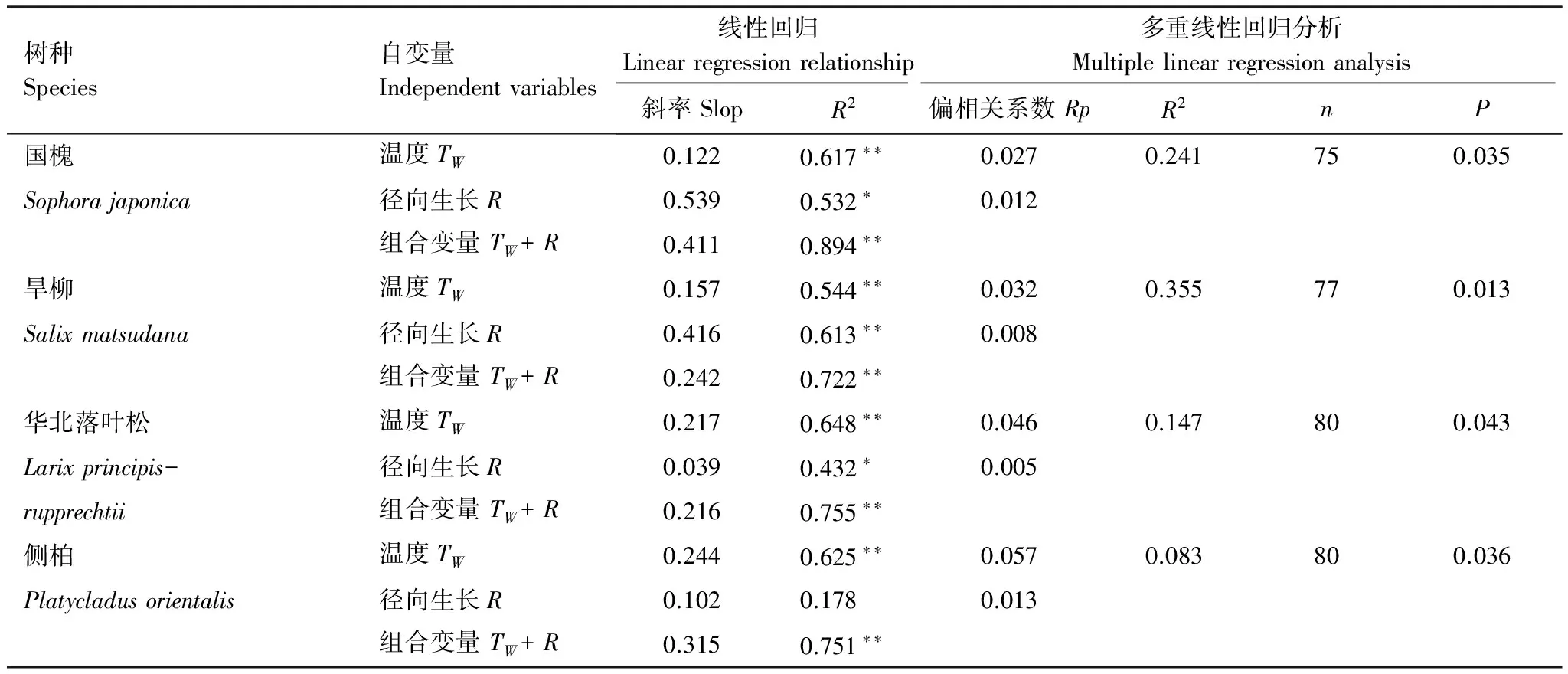
樹種Species自變量Independentvariables線性回歸Linearregressionrelationship多重線性回歸分析Multiplelinearregressionanalysis斜率SlopR2偏相關系數RpR2nP國槐溫度TW0.1220.617??0.0270.241750.035Sophorajaponica徑向生長R0.5390.532?0.012組合變量TW+R0.4110.894??旱柳溫度TW0.1570.544??0.0320.355770.013Salixmatsudana徑向生長R0.4160.613??0.008組合變量TW+R0.2420.722??華北落葉松溫度TW0.2170.648??0.0460.147800.043Larixprincipis-徑向生長R0.0390.432?0.005rupprechtii組合變量TW+R0.2160.755??側柏溫度TW0.2440.625??0.0570.083800.036Platycladusorientalis徑向生長R0.1020.1780.013組合變量TW+R0.3150.751??
TW:日均溫,mean daily temperature;R:月徑向生長速率,Monthly radial growth rate;Rp:偏相關系數,Partial correlation coefficients;**:P<0.01;*:P<0.05
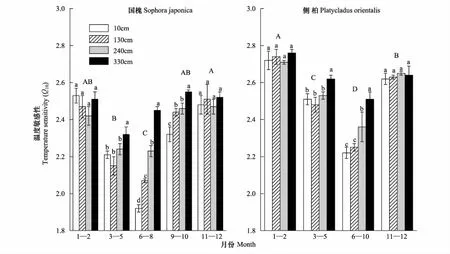
圖2 溫度敏感性系數(Q10)的垂直梯度分布Fig.2 Vertical variations in temperature-sensitivity coefficient (Q10)闊葉和針葉分別以國槐(a)和側柏(b)為例;不同大寫字母代表不同月份之間存在差異性差異,不同小寫字母代表不同高度之間存在差異(P<0.05),圖中數據為平均值±標準差(n=3)
根據Q10在不同月份之間有無顯著性差異,將全年分為4—5個時間區間。圖2中,分別以國槐和側柏為例,ECO2對溫度的敏感性在夏季最低,冬季最高。在樹木的生長階段(3—10月份),木質組織碳排放速率對溫度的敏感性(Q10)存在明顯的垂直梯度,且不同月份其分布模式不同。總體上,Q10隨木質組織高度的增加而增加。其中,闊葉樹種(以國槐為例)的鉛錘變異程度大于針葉樹種(以側柏為例)。對于本研究所選取的4個樹種,Q10在鉛垂梯度上的分布模式為:夏季(6—8月),闊葉樹種的Q10值在4個高度間均存在顯著差異(P<0.05),針葉樹種的樹干頂部(270 cm)和一級分枝(330 cm)Q10明顯高于另外兩個高度(P<0.05),樹干基部(10 cm)和胸高位置(130 cm)之間差異不明顯(P> 0.05);春(3—5月)、秋(9—10)季節闊葉樹種的垂直分區模式發生變化,分別由夏季的4區變為2區或3區,而針葉樹種的仍保持夏季原有的3區模式;在冬季月份(1—2和11—12),木質組織ECO2溫度敏感性在4個高度之間差異不顯著(P> 0.05),垂直分布模式消失(圖2)。
2.2.2日間分區和Q10日不對稱性分析
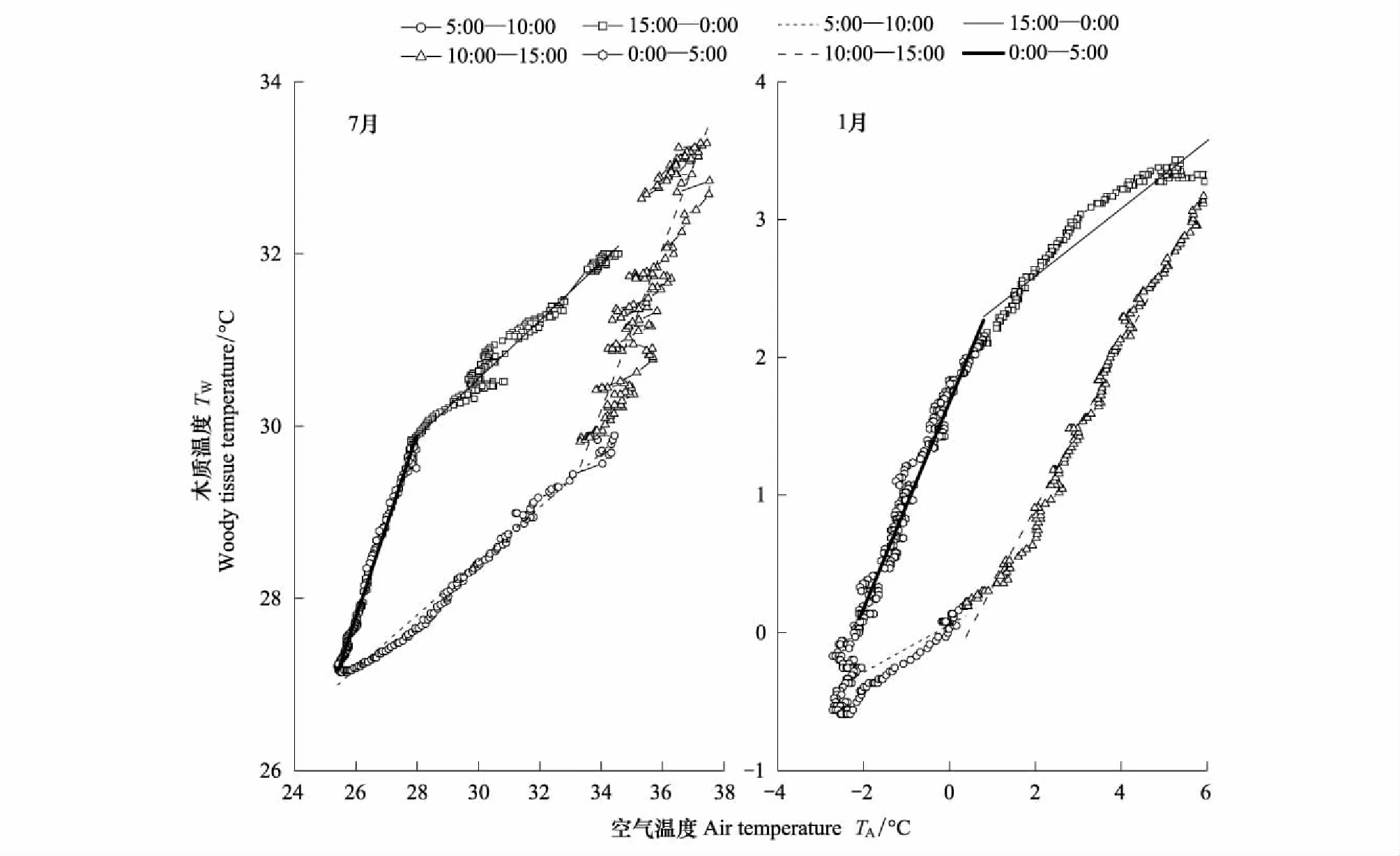
圖3 不同時段內枝干-大氣溫度相關分析(以130 cm高度為例)Fig.3 Correlation analysis of branches-air temperature in different periods of time
為比較Q10在日間尺度上的變異特征,本研究選取樹木的典型生長月份(7月)和非生長月份(1月)做了對比研究。根據氣溫(TA)與枝干溫度(TW)之間的相對高低和相關性強弱可以將一天分為4個時段,分別為00:00—05:00,05:00—10:00,10:00—15:00和15:00—24:00(圖3)。在每個時段內,TW與TA之間存在極強的相關性(R2> 0.70,P<0.01),且TW明顯滯后于TA(圖3中逆時針)。在不同時段內,其相關性明顯不同(生長季節尤為明顯):以國槐樹干的胸高位置(130 cm)為例(其他高度或樹種特點與之相似,表3),在生長月份(7月為代表),10:00—15:00和00:00—05:00為枝干快速傳熱階段,TW與TA的相關系數分別為0.33(R2=0.93,P<0.001)和0.36(R2=0.99,P<0.001)。其中,時段10:00—15:00內,TA高于TW,后者隨前者的升高而迅速升高;而時段00:00—05:00內,TW高于TA,此段內大氣溫度變化緩慢,而枝干內部溫度下降較快。05:00—10:00(TW隨TA緩慢上升)和15:00—00:00(TW隨TA緩慢下降)為慢速傳熱階段,此時氣溫高于枝干溫度,枝干溫度與氣溫的相關系數相對較低,分別為0.22(R2=0.98,P<0.01)和0.17(R2=0.88,P<0.01)。在非生長月份(以1月為例),TW和TA的相關性在各時段內差異不大(圖3)。
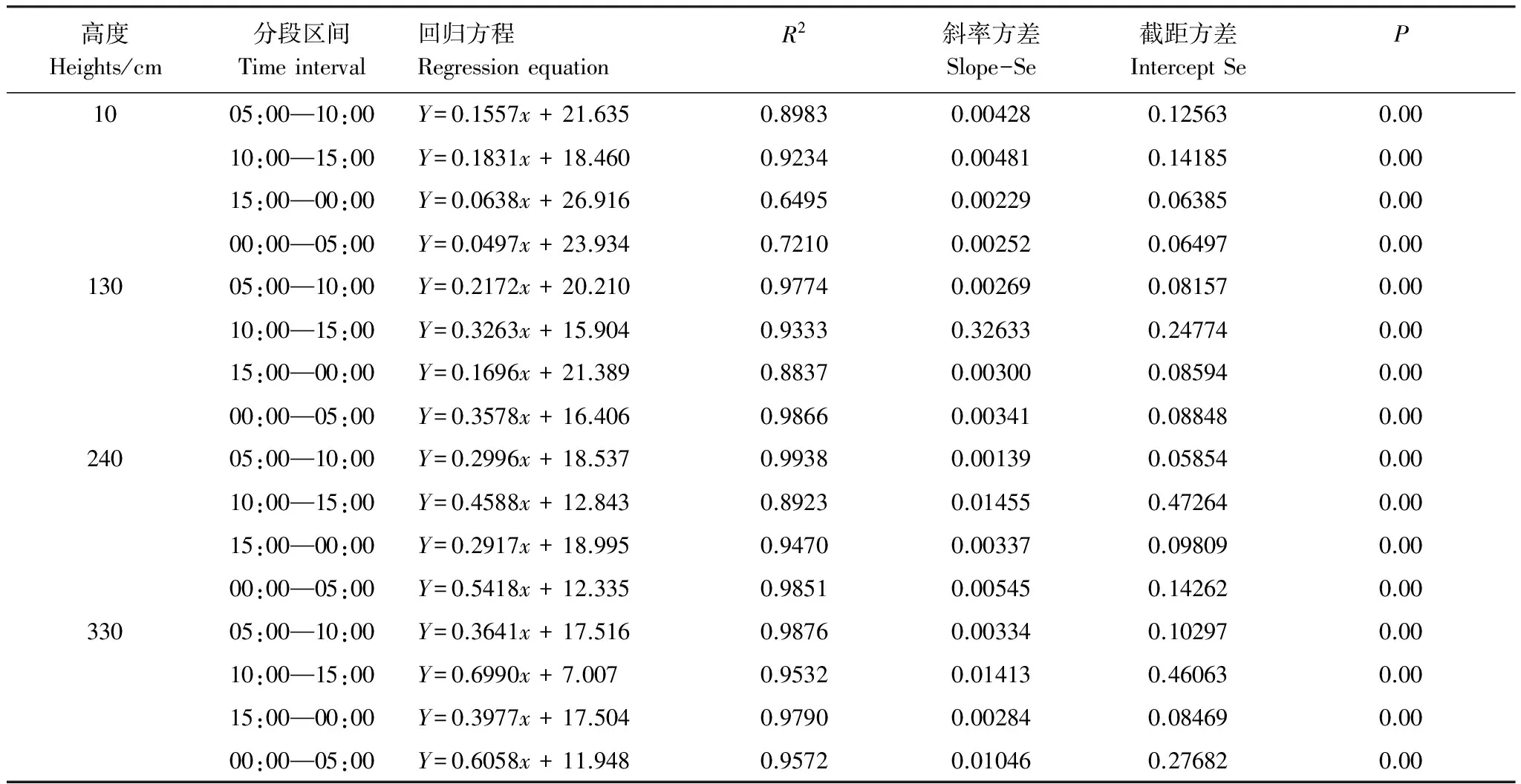
表3 生長季節枝干溫度和大氣溫度相關分析
為探究Q10的晝夜變化特征,本研究選擇了生長月份(6—8月)和非生長月份(1—3月),對4個時段內的Q10作了比較。研究結果顯示:在樹木的生長階段(6—8月份),闊葉樹種ECO2對溫度的敏感性存在晝夜不對稱性,即晚上Q10值顯著高于白天;在非生長月份(1—3月),這種不對稱性消失。全年內,針葉樹種在不存在日不對稱性(圖4)。
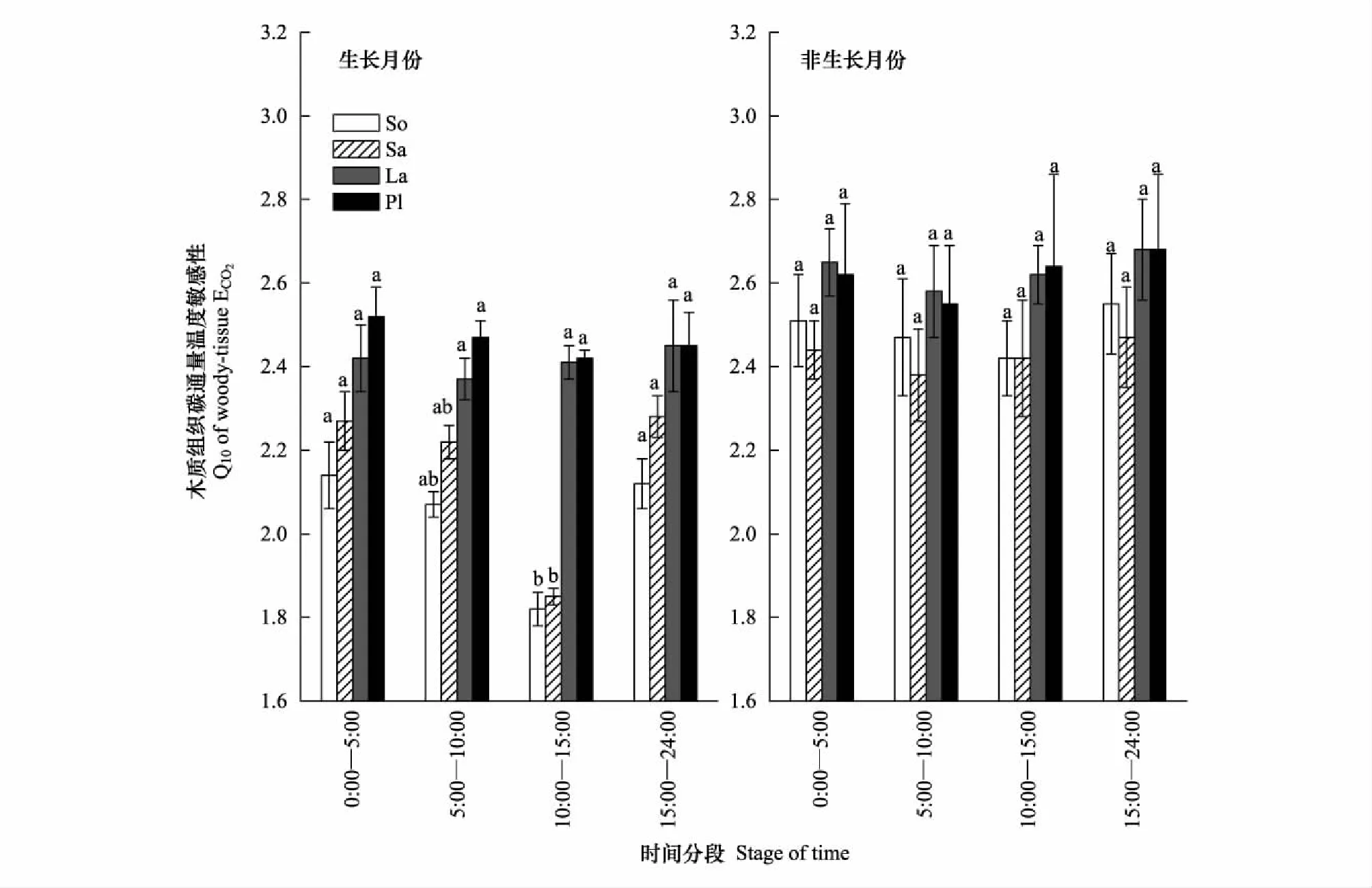
圖4 溫度敏感性系數(Q10)的日不對稱分析Fig.4 Daily asymmetry of temperature-sensitivity coefficient (Q10)
典型生長月份(6—8月)和非生長月份內Q10的日不對稱比較,不同小寫字母代表一天內不同時段之間存在差異(P<0.05),圖中數據為平均值±標準差(n=3)
3 結果與討論
本研究所選取的4個樹種,枝干表面CO2通量速率(ECO2)均表現為單峰型季節變化節律(圖1),與先前Ceschia等和Gruber等在歐洲山毛櫸(FagussylvaticaL.)和瑞士五針松(Pinus cembra L.)中得到的結論相一致[16,40]。通過對ECO2和木質組織溫度(Tw)的回歸分析發現,兩者之間存在極強的正相關關系(表2)。因此,Tw是導致ECO2季節變化的因素之一,這與前人的研究結果吻合[8,19,28]。同時,本研究也發現,Tw的本身并不能完全解釋ECO2的季節變異性,溫度歸一化處理后的ECO2(歸一到0℃,用E0表示)仍呈現單峰或雙峰型季節變化節律(圖1)。對于研究中的4個樹種,ECO2與對應月份內枝干直徑增長速率有明顯的強相關關系(P<0.05)(表2),該結果表明ECO2除了受溫度影響以外,還與樹木本身的生長狀況密切相關[41]。這可能是由于在生長期間,植物體通過加強呼吸速率來為細胞分化以及隨后的胞壁加厚以及木質化等過程提供更多的呼吸底物,以實現正常生長[42]。此外,樹木生長期間皮孔對水汽的滲透性增加,也可能會導致植物體內CO2向周邊大氣的排放增多。以上結果表明,枝干呼吸的季節變化除了受組織溫度影響以外,還與樹木生長狀況甚至樹皮的組織結構密切相關[43]。
本研究中國槐,旱柳,華北落葉松和側柏的年均Q10值分別為2.32,2.09,2.57和2.49,略高于生態系統模型中的默認值2.0[36]。ECO2對不同時間尺度上的溫度變化,有不同程度的敏感性[44]。本研究發現,ECO2在短時間內(日間尺度)的溫度響應特征與長時間內(年內尺度)的溫度響應特征存在明顯的不同。年際尺度上的生長代謝和晝夜尺度上的液流變化等因素會直接影響ECO2與溫度之間的相關關系[45],因此精細尺度上的溫度敏感性計算更能反應ECO2對溫度的響應[44,46]。通過對每個月份內的Q10對比研究發現,ECO2對溫度的敏感性存在明顯的季節變化。夏季ECO2對溫度的敏感性顯著小于冬季(圖2),Q10隨樹干組織溫度的升高而降低,與之前的研究結果相一致[25]。這可能是因為在低溫環境下,酶活性是抑制呼吸的主要因素,而高溫環境下底物供應成為限制因素,植物為了適應這種轉變而做出的生理生態響應[47]。
同時,ECO2對溫度的敏感性在不同月份內表現為不同的日間變化動態,在樹木的生長月份,ECO2對溫度的敏感性呈現出明顯的垂直梯度分區。例如,6—8月份,國槐的Q10值從樹干基本的1.92增至一級分支的2.45(圖2)。一定程度上,皮層的薄厚程度會直接影響到CO2氣體向周邊大氣的擴散,從而改變ECO2對溫度的敏感性[48],使得ECO2對溫度的響應呈現出自上而下的梯度變化。另一方面,不同徑級與大氣之間的傳熱快慢也會影響到ECO2對溫度的敏感性。對于細徑而言,枝干與大氣之間可以快速達到熱量平衡,從而表現為細徑級的溫度敏感性明顯升高[49]。除此之外,木質組織的代謝活性也會影響到ECO2對溫度的敏感性,本研究發現一級分枝的代謝活性明顯高于其他徑級(圖1),高的代謝活性可以促進CO2氣體的外部釋放,進而改變木質組織對外界環境的響應[50]。
在一天中,木質組織與大氣溫度之間的平衡存在明顯的時間分區(圖3)。中午時段(10:00—15:00),木質組織與大氣之間的傳熱最為迅速,但相應的溫度敏感性最低(圖4)。這可能是由于午間溫度迅速升高導致:有研究發現植物呼吸對溫度的敏感性會隨溫度的升高而降低,例如,Tjoelker等發現葉片溫度每升高1℃,溫度敏感性系數Q10降低0.04[51]。除此之外,光合作用的“午間抑制”也可能是造成Q10值在白天下降的原因之一。光合作用的抑制和氣孔的關閉都會造成ECO2下降,進而改變ECO2對溫度的敏感性[52]。在絕大多數生態系統模型中,Q10被設置為常數2.0,與白天的Q10數值相似,但遠遠低于晚上的Q10值,因此會對晚上的碳排放估測造成誤差。因此,本研究表明,在對木質組織碳排放進行估算時,應根據問題所涉及的時-空間尺度,選擇合適的Q10值進行自下而上的碳推算。
[1] Ryan M G, Hubbard R M, Clark D A, Sanford R L Jr. Woody-tissue respiration forSimaroubaamaraandMinquartiaguianensis, two tropical wet forest trees with different growth habits. Oecologia, 1994, 100(3): 213- 220.
[2] Maier C A, Albaugh T J, Allen H L, Dougherty P M. Respiratory carbon use and carbon storage in mid-rotation loblolly pine (PinustaedaL.) plantations: the effect of site resources on the stand carbon balance. Global Change Biology, 2004, 10(8): 1335- 1350.
[3] 石立新, 王傳寬, 許飛, 王興昌. 四個溫帶樹種樹干呼吸的時間動態及其影響因子. 生態學報, 2010, 30(15): 3994- 4003.
[4] Lavigne M B, Ryan M G. Growth and maintenance respiration rates of aspen, black spruce and jack pine stems at northern and southern BOREAS sites. Tree Physiology, 1997, 17(8/9): 543- 551.
[5] 朱娟, 班遠沖. 陸地生態系統呼吸溫度敏感性及影響因素研究. 環境科學與管理, 2017, 42(1): 158- 161.
[6] Amthor J S. Plant respiratory responses to the environment and their effects on the carbon balance//Wilkinson R E, ed. Plant-Environment Interactions. New York: Marcel Dekker, 1994: 501- 554.
[7] Valentini R, De Angelis P, Matteucci G, Monaco R, Dore S, Mucnozza G E S. Seasonal net carbon dioxide exchange of a beech forest with the atmosphere. Global Change Biology, 1996, 2(3): 199- 207.
[8] Zha T S, Kellom?ki S, Wang K Y, Ryypp? A, Niinist? S. Seasonal and annual stem respiration of Scots pine trees under boreal conditions. Annals of Botany, 2004, 94(6): 889- 896.
[9] Maier C A, Clinton B D. Relationship between stem CO2efflux, stem sap velocity and xylem CO2concentration in young loblolly pine trees. Plant, Cell & Environment, 2006, 29(8): 1471- 1483.
[10] Teskey R O, Saveyn A, Steppe K, McGuire M A. Origin, fate and significance of CO2in tree stems. New Phytologist, 2008, 177(1): 17- 32.
[11] Chambers J Q, Tribuzy E S, Toledo L C, Crispim B F, Higuchi N, dos Santos J, Araújo A C, Kruijt B, Nobre A D, Trumbore S E. Respiration from a tropical forest ecosystem: partitioning of sources and low carbon use efficiency. Ecological Applications, 2004, 14(sp4): 72- 88.
[12] Harris N L, Hall C A S, Lugo A E. Estimates of species- and ecosystem-level respiration of woody stems along an elevational gradient in the Luquillo Mountains, Puerto Rico. Ecological Modelling, 2008, 216(3/4): 253- 264.
[13] Robertson A L, Malhi Y, Farfan-Amezquita F, Aragao L E O C, Espejo J E S, Robertson M A. Stem respiration in tropical forests along an elevation gradient in the Amazon and Andes. Global Change Biology, 2011, 16(12): 3193- 3204.
[14] Katayama A, Kume T, Komatsu H, Ohashi M, Matsumoto K, Ichihashi R, Kumagai T, Otsuki K. Vertical variations in wood CO2efflux for live emergent trees in a Bornean tropical rainforest. Tree Physiology, 2014, 34(5): 503- 512.
[15] Stockfors J, Linder S. Effect of nitrogen on the seasonal course of growth and maintenance respiration in stems of Norway spruce trees. Tree Physiology, 1998, 18(3): 155- 166.
[16] Ceschia é, Damesin C, Lebaube S, Pontailler J Y, Dufrêne é. Spatial and seasonal variations in stem respiration of beech trees (Fagussylvatica). Annals of Forest Science, 2002, 59(8): 801- 812.
[17] Wieser G, Bahn M. Seasonal and spatial variation of woody tissue respiration in aPinuscembratree at the alpine timberline in the central Austrian Alps. Trees, 2004, 18(5): 576- 580.
[18] Maunoury F, Berveiller D, Lelarge C, Pontailler J Y, Vanbostal L, Damesin C. Seasonal, daily and diurnal variations in the stable carbon isotope composition of carbon dioxide respired by tree trunks in a deciduous oak forest. Oecologia, 2007, 151(2): 268- 279.
[20] Brito P, Morales D, Wieser G, Jiménez M S. Spatial and seasonal variations in stem CO2efflux ofPinuscanariensisat their upper distribution limit. Trees, 2010, 24(3): 523- 531.
[21] Tarvainen L, R?ntfors M, Wallin G. Vertical gradients and seasonal variation in stem CO2efflux within a Norway spruce stand. Tree Physiology, 2014, 34(5): 488- 502.
[22] Paembonan S A, Hagihara A, Hozumi K. Long-term measurement of CO2release from the aboveground parts of a hinoki forest tree in relation to air temperature. Tree Physiology, 1991, 8(4): 399- 405.
[23] Stockfors J. Temperature variations and distribution of living cells within tree stems: implications for stem respiration modeling and scale-up. Tree Physiology, 2000, 20(15): 1057- 1062.
[24] Ryan M G. Effects of climate change on plant respiration. Ecological Applications, 1991, 1(2): 157- 167.
[25] Atkin O K, Tjoelker M G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends in Plant Science, 2003, 8(7): 343- 351.
[27] Zhu L W, Zhao P, Ni G Y, Cao Q P, Zhou C M, Zeng X P. Individual- and stand-level stem CO2efflux in a subtropicalSchimasuperbaplantation. Biogeosciences, 2012, 9(10): 3729- 3737.
[28] Ryan M G, Gower S T, Hubbard R M, Waring R H, Gholz H L, Cropper W P Jr, Running S W. Woody tissue maintenance respiration of four conifers in contrasting climates. Oecologia, 1995, 101(2): 133- 140.
[29] Amthor J S. The McCree-de Wit-Penning de Vries-Thornley respiration paradigms: 30 years later. Annals of Botany, 2000, 86(1): 1- 20.
[30] Melillo J M, McGuire A D, Kicklighter D W, Moore B, Vorosmarty C J, Schloss A L. Global climate change and terrestrial net primary production. Nature, 1993, 363(6426): 234- 240.
[31] Turnbull M H, Whitehead D, Tissue D T, Schuster W S F, Brown K J, Griffin K L. Responses of leaf respiration to temperature and leaf characteristics in three deciduous tree species vary with site water availability. Tree Physiology, 2001, 21(9): 571- 578.
[32] Atkin O K, Zhang Q S, Wiskich J T. Effect of temperature on rates of alternative and cytochrome pathway respiration and their relationship with the redox poise of the Quinone pool. Plant Physiology, 2002, 128(1): 212- 222.
[33] Covey-Crump E M, Attwood R G, Atkin O K. Regulation of root respiration in two species ofPlantagothat differ in relative growth rate: the effect of short- and long-term changes in temperature. Plant, Cell & Environment, 2002, 25(11): 1501- 1513.
[34] Loveys B R, Atkinson L J, Sherlock D J, Roberts R L, Fitter A H, Atkin O K. Thermal acclimation of leaf and root respiration: an investigation comparing inherently fast- and slow-growing plant species. Global Change Biology, 2003, 9(6): 895- 910.
[35] Lavigne M B. Comparing stem respiration and growth of jack pine provenances from northern and southern locations. Tree Physiology, 1996, 16(10): 847- 852.
[36] Yang Y, Zhao M, Xu X T, Sun Z Z, Yin G D, Piao S L. Diurnal and seasonal change in stem respiration ofLarixprincipis-rupprechtiitrees, northern China. PLoS One, 2014, 9(2): e89294.
[37] Damesin C, Ceschia E, Le Goff N, Ottorini J M, Dufrêne E. Stem and branch respiration of beech: from tree measurements to estimations at the stand level. New Phytologist, 2002, 153(1): 159- 172.
[38] McGuire M A, Cerasoli S, Teskey R O. CO2fluxes and respiration of branch segments of sycamore (PlatanusoccidentalisL.) examined at different sap velocities, branch diameters, and temperatures. Journal of Experimental Botany, 2007, 58(8): 2159- 2168.
[39] Kim M H, Nakane K, Lee J T, Bang H S, Na Y E. Stem/branch maintenance respiration of Japanese red pine stand. Forest Ecology and Management, 2007, 243(2/3): 283- 290.
[40] Gruber A, Wieser G, Oberhuber W. Intra-annual dynamics of stem CO2efflux in relation to cambial activity and xylem development inPinuscembra. Tree Physiology, 2009, 29(5): 641- 649.
[41] 孫濤, 劉瑞鵬, 李興歡, 毛子軍, 黃唯, 董利利. 模擬氮沉降對東北地區興安落葉松樹干呼吸的影響. 生態學報, 2015, 35(11): 3684- 3691.
[42] Rossi S, Deslauriers A, Morin H. Application of the Gompertz equation for the study of xylem cell development. Dendrochronologia, 2003, 21(1): 33- 39.
[43] 許飛, 王傳寬, 王興昌. 東北東部14個溫帶樹種樹干呼吸的種內種間變異. 生態學報, 2011, 31(13): 3581- 3589.
[44] Yuste J C, Janssens I A, Carrara A, Ceulemans R. AnnualQ10of soil respiration reflects plant phenological patterns as well as temperature sensitivity. Global Change Biology, 2004, 10(2): 81- 94.
[45] Sampson D A, Janssens I A, Yuste J C, Ceulemans R. Basal rates of soil respiration are correlated with photosynthesis in a mixed temperate forest. Global Change Biology, 2007, 13(9): 2008- 2017.
[46] Janssens I A, Pilegaard K. Large seasonal changes inQ10of soil respiration in a beech forest. Global Change Biology, 2003, 9(6): 911- 918.
[47] Atkin O K, Bruhn D, Hurry V M, Tjoelker M G.EvansreviewNo.2: the hot and the cold: unravelling the variable response of plant respiration to temperature. Functional Plant Biology, 2005, 32(2): 87- 105.
[48] Eklund L, Lavigne M B. Restricted lateral gas movement inPinusstrobusbranches. Trees, 1995, 10(2): 83- 85.
[49] 龐卓, 余新曉, 朱建剛. 樹干自然溫度梯度變化對熱擴散法測算樹干液流速率的影響. 生態學報, 2010, 30(3): 635- 644.
[50] Gaumont-Guay D, Black T A, Barr A G, Jassal R S, Nesic Z. Biophysical controls on rhizospheric and heterotrophic components of soil respiration in a boreal black spruce stand. Tree Physiology, 2008, 28(2): 161- 171.
[51] Tjoelker M G, Oleksyn J, Reich P B. Modelling respiration of vegetation: evidence for a general temperature-dependentQ10. Global Change Biology, 2001, 7(2): 223- 230.
[52] Peters J, Morales D, Jimenez M S. Gas exchange characteristics ofPinuscanariensisneedles in a forest stand on Tenerife, Canary Islands. Trees, 2003, 17(6): 492- 500.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
