雙臺(tái)子河口文蛤碳、氮、磷收支的季節(jié)變化
2018-03-02 05:24:16張安國(guó)王麗麗馬恭博張學(xué)輝袁秀堂國(guó)家海洋環(huán)境監(jiān)測(cè)中心遼寧大連60河北省地礦局第四水文工程地質(zhì)大隊(duì)河北滄州06000盤(pán)山縣水產(chǎn)技術(shù)推廣站遼寧盤(pán)錦400
中國(guó)環(huán)境科學(xué) 2018年2期
關(guān)鍵詞:生長(zhǎng)
張安國(guó),王麗麗,袁 蕾,馬恭博,張學(xué)輝,雷 帥,袁秀堂* (.國(guó)家海洋環(huán)境監(jiān)測(cè)中心,遼寧 大連60;.河北省地礦局第四水文工程地質(zhì)大隊(duì),河北 滄州 06000;.盤(pán)山縣水產(chǎn)技術(shù)推廣站,遼寧 盤(pán)錦 400)
雙殼貝類(lèi)在河口、淺海和深海等區(qū)域均有分布,但以沿岸海域?yàn)樽钪匾獥⒌?其種類(lèi)多,數(shù)量大.雙殼貝類(lèi)具有較強(qiáng)的濾水能力,通過(guò)大量的濾水和攝食來(lái)攝取海水中的有機(jī)顆粒物質(zhì),加快其沉降速率,同時(shí)減少海水中的浮游生物量.雙殼貝類(lèi)還通過(guò)排泄過(guò)程將大量的營(yíng)養(yǎng)鹽排入水體中,對(duì)浮游植物生長(zhǎng)具有重要意義;特別是無(wú)機(jī)形態(tài)的N()和P()的再循環(huán),成為維持海洋初級(jí)生產(chǎn)力的重要營(yíng)養(yǎng)鹽來(lái)源[1-8].因此,濾食性雙殼貝類(lèi)通過(guò)生物沉積和呼吸排泄作用加快了C、N、P等生源要素在水層與底層間的流動(dòng)和交換[4-5],在近海沿岸生態(tài)系統(tǒng)的物質(zhì)循環(huán)中扮演著重要角色[9].
文蛤(Meretrix meretrix)屬于灘涂埋棲性雙殼貝類(lèi),是我國(guó)沿海灘涂重要的經(jīng)濟(jì)貝類(lèi)之一.文蛤分布較廣,主要棲息在河口潮間帶以下和潮下帶靠上的沙質(zhì)或泥質(zhì)灘涂[10].雙臺(tái)子河口位于我國(guó)渤海遼東灣頂部,灘涂面積約為6.7×104hm2,是我國(guó)北方灘涂經(jīng)濟(jì)貝類(lèi)的主要棲息場(chǎng)所之一[11].因此,現(xiàn)場(chǎng)研究該河口灘涂?jī)?yōu)勢(shì)種類(lèi)和重要經(jīng)濟(jì)貝類(lèi)——文蛤的C、N、P等生源要素的收支狀況,對(duì)評(píng)價(jià)灘涂埋棲性貝類(lèi)在河口生態(tài)系統(tǒng)物質(zhì)循環(huán)中的作用具有重要意義.
目前,國(guó)內(nèi)外關(guān)于雙殼貝類(lèi)生源要素收支的研究報(bào)道較少,僅見(jiàn)于我國(guó)山東煙臺(tái)四十里灣櫛孔扇貝(Chlamys farreri)[12]、浙江樂(lè)清灣和三門(mén)灣太平洋牡蠣(Crassostrea gigas)、僧帽牡蠣(Ostrea cucullata)、泥蚶(Tegillarca granosa)、縊蟶(Sinonovacula constricta)的C收支[13];遼寧雙臺(tái)子河口四角蛤蜊(Mactra veneriformis)[14]、莊河海域菲律賓蛤仔(Ruditapes philippinarum)[15]、山東榮成桑溝灣櫛孔扇貝、太平洋牡蠣和蝦夷扇貝(Patinopecten yesoensis)的C、N、P收支[16]、長(zhǎng)江口縊蟶、光滑河藍(lán)蛤C、N收支[17];日本東京灣四角蛤蜊的N收支[18]和荷蘭Oosterschelde河口海域貽貝(Mytilus edulis)C、N、P收支[19-20].而關(guān)于文蛤的C、N、P收支研究較少,僅見(jiàn)在實(shí)驗(yàn)室條件下文蛤的C收支狀況[21],未見(jiàn)基于現(xiàn)場(chǎng)條件的文蛤C、N、P收支及生長(zhǎng)余力(SFG)的研究.課題組前期[11]在雙臺(tái)子河口海域現(xiàn)場(chǎng)測(cè)定了文蛤的主要生理生態(tài)學(xué)參數(shù),為構(gòu)建文蛤C、N、P等生源要素收支和估算其SFG提供了基礎(chǔ)數(shù)據(jù).本研究作為該文的延續(xù),利用現(xiàn)場(chǎng)測(cè)定的生理生態(tài)學(xué)參數(shù)估算文蛤的SFG,并構(gòu)建現(xiàn)場(chǎng)條件下C、N、P收支方程,進(jìn)而對(duì)不同季節(jié)文蛤的C、N、P收支狀況和SFG進(jìn)行分析.本研究結(jié)果有助于從生源要素角度分析評(píng)價(jià)灘涂埋棲性貝類(lèi)在河口生態(tài)系統(tǒng)的物質(zhì)循環(huán)中所扮演的生態(tài)角色,也可為經(jīng)濟(jì)貝類(lèi)的碳通量以及碳交易提供基礎(chǔ)數(shù)據(jù).
1 研究海域和研究方法
1.1 研究海域
選擇我國(guó)渤海雙臺(tái)子河口的盤(pán)山海域?yàn)楝F(xiàn)場(chǎng)試驗(yàn)研究區(qū)域(圖1).該海域的灘涂底質(zhì)以泥沙質(zhì)及泥質(zhì)為主,海水中有機(jī)顆粒物含量豐富,是文蛤生長(zhǎng)及繁殖的主要場(chǎng)所.試驗(yàn)地點(diǎn)位于雙臺(tái)子河口灘涂高潮線(xiàn)附近面積6hm2、水深1.5~2.0m的池塘中.該池塘通過(guò)潮水溝渠與河口海域相通,并根據(jù)當(dāng)?shù)爻毕?guī)律定時(shí)納入新鮮的海水,保證池塘中海水水質(zhì)條件與自然海區(qū)水質(zhì)條件基本一致.
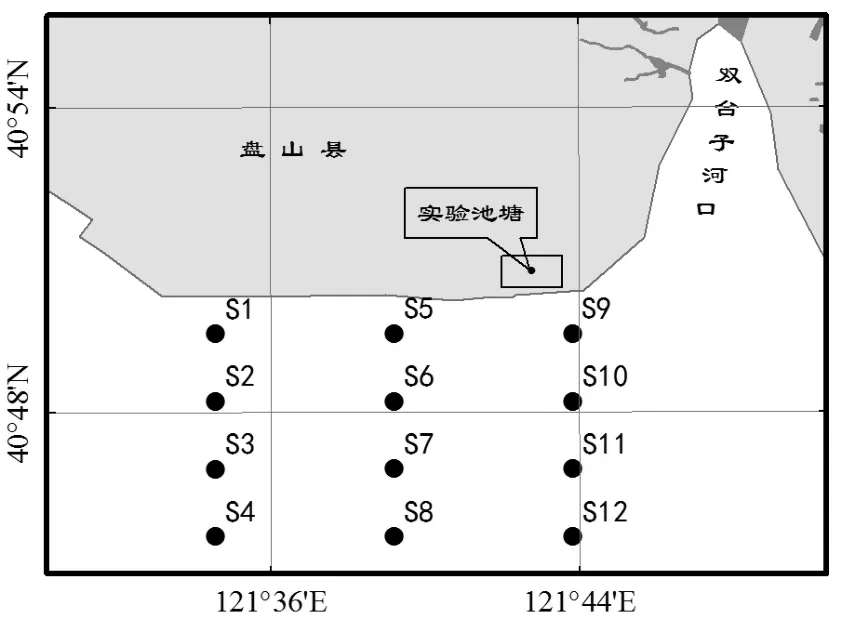
圖1 雙臺(tái)子河口試驗(yàn)地點(diǎn)Fig.1 Experimental sites in Shuangtaizi estuary
1.2 文蛤主要生理生態(tài)參數(shù)和試驗(yàn)池塘環(huán)境參數(shù)測(cè)定
分別于2010年7月(夏季)、10月(秋季)、12月(冬季)及2011年4月(春季)進(jìn)行文蛤主要生理生態(tài)學(xué)參數(shù)(生物沉積速率、耗氧率、排氨率和排磷率等)和試驗(yàn)池塘相關(guān)環(huán)境參數(shù)的測(cè)定.文蛤主要生理生態(tài)學(xué)參數(shù)和試驗(yàn)池塘環(huán)境參數(shù)的各季節(jié)具體數(shù)值見(jiàn)文獻(xiàn)[11].
1.3 文蛤真糞與假糞比例的估算
研究表明,當(dāng)海水中顆粒物濃度足夠高時(shí)可引起貝類(lèi)產(chǎn)生假糞.因此,為準(zhǔn)確判斷河口海域文蛤個(gè)體是否產(chǎn)生假糞,以及產(chǎn)生的假糞與真糞的比例,本試驗(yàn)參照四角蛤蜊真糞與假糞的測(cè)定方法[19](并略作改動(dòng))來(lái)現(xiàn)場(chǎng)測(cè)定二齡和三齡文蛤個(gè)體真糞與假糞的比例.具體試驗(yàn)方法如下:現(xiàn)場(chǎng)條件下將文蛤個(gè)體分別放入盛有試驗(yàn)池塘海水的塑料燒杯(體積為2L)中,并將塑料燒杯放置于試驗(yàn)池塘邊緣,池塘海水不沒(méi)過(guò)燒杯頂部為止,以保持試驗(yàn)溫度與池塘海水溫度(16℃)一致.試驗(yàn)共設(shè)13個(gè)塑料燒杯,其中10個(gè)放置文蛤,另3個(gè)不放文蛤作為對(duì)照,試驗(yàn)重復(fù)3次.經(jīng)觀(guān)察發(fā)現(xiàn),文蛤的假糞一般呈顆粒團(tuán)狀,出現(xiàn)在進(jìn)水管一端;真糞呈線(xiàn)狀,在出水管一端.24h后分別收集文蛤的真糞及假糞,將糞便(真糞和假糞)分別虹吸過(guò)濾至事先于450℃下灼燒6h并稱(chēng)量的GF/F濾膜(直徑47mm)上,-20℃冷凍保存待測(cè).將試驗(yàn)樣品帶回實(shí)驗(yàn)室后將濾膜在65℃條件下烘干至恒重,用精密電子天平(精確到0.1mg)分別準(zhǔn)確稱(chēng)量文蛤產(chǎn)生的真糞和假糞重量.試驗(yàn)時(shí)取試驗(yàn)池塘水樣過(guò)濾,以測(cè)定水體的總懸浮顆粒物(TPM),即用上述經(jīng)過(guò)處理的GF/F過(guò)濾膜兩張分別過(guò)濾250~500mL水樣,用蒸餾水脫鹽并將濾膜在 65℃條件下干燥24h后,稱(chēng)量以測(cè)定試驗(yàn)池塘水中的TPM含量.
1.4 文蛤?yàn)V水率、攝食率的估算
根據(jù)生物沉積法的原理進(jìn)行文蛤生理生態(tài)學(xué)參數(shù)的測(cè)定:

式中:IFR為文蛤?qū)o(wú)機(jī)物的過(guò)濾速率,Rb為文蛤的生物沉積速率.
文蛤的濾水率(CR):
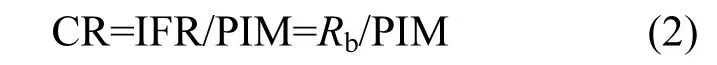
式中:PIM為文蛤所過(guò)濾海水中顆粒無(wú)機(jī)物濃度.Rb的測(cè)定方法及結(jié)果均引自文獻(xiàn)[11].
文蛤的總濾食速率(FR):

式中:TPM為文蛤所過(guò)濾海水中的總顆粒物濃度.
當(dāng)海水中顆粒物濃度足夠高以引起貝類(lèi)假糞的產(chǎn)生時(shí),則文蛤的攝食率(IR):

式中:RR為文蛤?qū)λ^(guò)濾的食物拒絕攝食的速率,即文蛤假糞的排出速率.因此,當(dāng)顆粒物濃度很低并不足以引起假糞的產(chǎn)生時(shí),則攝食率IR=FR.
當(dāng)文蛤產(chǎn)生假糞時(shí),文蛤假糞的排出速率:
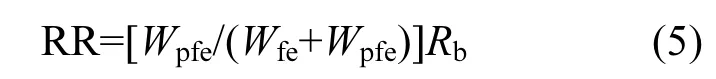
攝食率:

式中:Wfe為文蛤產(chǎn)生的真糞重量,g;Wpfe為文蛤產(chǎn)生的假糞重量,g.
文蛤?qū)︻w粒有機(jī)物質(zhì)C、N、P的攝食率(RI)計(jì)算公式如下:
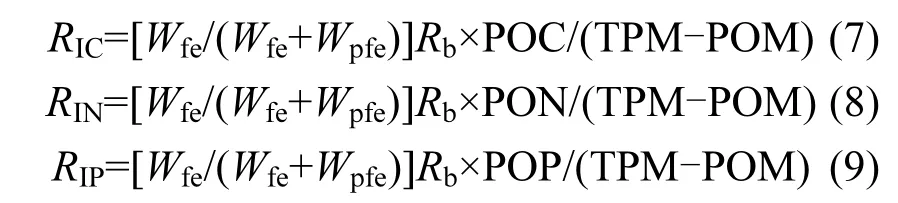
式中:TPM為試驗(yàn)海域水體的總懸浮顆粒物含量,mg/L;POM為顆粒有機(jī)質(zhì)含量,mg/L;POC為顆粒有機(jī)碳含量,mg/L;PON為顆粒有機(jī)氮含量,mg/L;POP為顆粒有機(jī)磷含量,μg/L.
1.5 文蛤C、N、P排遺率的估算
文蛤?qū)︻w粒有機(jī)物質(zhì)C、N、P的(排遺率)生物沉積速率:
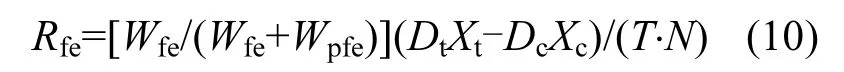
式中:Dt為文蛤捕集器中沉積物的質(zhì)量,g;Dc為對(duì)照組捕集器中沉積物的質(zhì)量,g;Xt為文蛤捕集器所收集沉積物的C、N、P的百分含量;Xc為對(duì)照捕集器所收集沉積物中C、N、P的百分含量;T為時(shí)間,d;N為試驗(yàn)用文蛤個(gè)數(shù).
1.6 文蛤C、N、P排泄速率的估算
文蛤耗氧率(Ro)、排氨率和排磷率的測(cè)定方法及結(jié)果均引自文獻(xiàn)[11].根據(jù)1μmol O2=0.85μmol CO2,碳排泄速率:
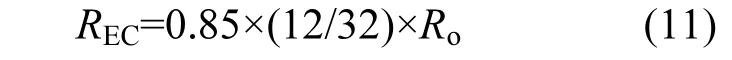
將排氨率和排磷率分別轉(zhuǎn)化為氮排泄速率(RN)和磷排泄速率(RP).
1.7 文蛤C、N、P收支模型
將上述相關(guān)參數(shù)轉(zhuǎn)換成文蛤C、N、P元素收支模型[19]:
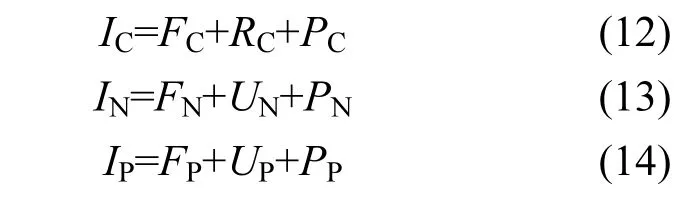
式中:IC為文蛤攝取的有機(jī)碳量;FC為文蛤通過(guò)糞便排出的碳量;RC為文蛤呼吸代謝消耗的碳量;PC為文蛤用于生長(zhǎng)的碳量;IN為文蛤攝取的有機(jī)氮量;FN為文蛤通過(guò)糞便排出的氮量;UN為文蛤排泄消耗的氮量;PN為文蛤用于生長(zhǎng)的氮量;IP為文蛤攝取的有機(jī)磷量;FP為文蛤通過(guò)糞便排出的磷量;UP為文蛤排泄消耗的磷量;PP為文蛤用于生長(zhǎng)的磷量.
1.8 文蛤C、N、P 的SFG及生長(zhǎng)效率的估算
在測(cè)定文蛤?qū)、N、P的攝食、排糞、排泄和吸收的基礎(chǔ)上對(duì)這3種元素的SFG和總生長(zhǎng)效率及凈生長(zhǎng)效率進(jìn)行計(jì)算.
本試驗(yàn)中,文蛤?qū)、N、P的生長(zhǎng)余力分別為:
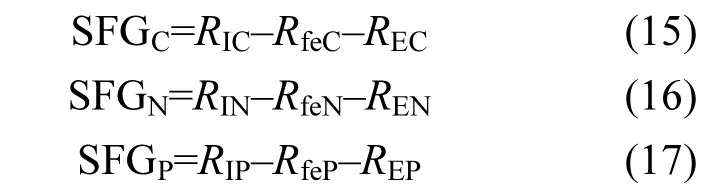
式中:RfeC、RfeN、RfeP分別為文蛤糞便C、N、P排出速率(mg/ind·d),REC、REN、REN分別為文蛤C、N、P排泄速率(mg/ind·d).
參照文獻(xiàn)[13,15]分別計(jì)算文蛤?qū)、N、P生源要素的總生長(zhǎng)效率(K1)和凈生長(zhǎng)效率(K2).
1.9 數(shù)據(jù)標(biāo)準(zhǔn)化和統(tǒng)計(jì)分析
應(yīng)用SPSS19.0軟件包對(duì)實(shí)驗(yàn)數(shù)據(jù)進(jìn)行單因素(季節(jié))方差分析,并結(jié)合Duncan法進(jìn)行多重比較.同時(shí)利用SPSS19.0軟件對(duì)文蛤生理生態(tài)學(xué)參數(shù)與水溫等環(huán)境因子進(jìn)行Pearson相關(guān)分析,并以P=0.05作為差異顯著的標(biāo)準(zhǔn).
2 結(jié)果與分析
2.1 文蛤真糞與假糞比例
文蛤真糞和假糞的實(shí)驗(yàn)結(jié)果分析可知,二齡和三齡文蛤個(gè)體在溫度為16℃、TPM 為48.5mg/L的條件下產(chǎn)生的真糞所占比例分別為86%和100%.
2.2 文蛤的CR和IR
由表1可以看出,雙臺(tái)子河口文蛤的CR和IR均呈顯著的季節(jié)性變化(P<0.05),夏季最高,冬季最低.文蛤?qū)、N、P生源要素的攝食速率表現(xiàn)為RIC>RIN>RIP.相關(guān)分析結(jié)果顯示,文蛤的CR和IR均與海水溫度呈顯著的相關(guān)關(guān)系,文蛤的CR與TPM無(wú)顯著相關(guān)性,而攝食率則與TPM呈極顯著關(guān)系(表2).文蛤的RIC與海水溫度及POC濃度呈極顯著的正相關(guān)關(guān)系,而與TPM含量無(wú)顯著相關(guān)性.文蛤的RIN與海水溫度及PON含量呈極顯著相關(guān)性,與TPM含量呈顯著相關(guān)關(guān)系.文蛤的RIP與海水中TPM及POP含量均呈極顯著的相關(guān)關(guān)系,而與海水溫度無(wú)顯著相關(guān)性.
2.3 文蛤的RE和Rfe
如表1所示,文蛤的RE和Rfe均呈現(xiàn)明顯的季節(jié)性變化(P<0.05).文蛤的REC(除夏季外)、REN、REP均低于其RfeC、RfeN、RfeP.相關(guān)性分析結(jié)果顯示,文蛤的REC和RfeC均與海水溫度呈顯著的相關(guān)關(guān)系,與TPM均無(wú)顯著相關(guān)性,REC與POC也具有顯著相關(guān)性,而RfeC與POC無(wú)顯著相關(guān)性(見(jiàn)表2).文蛤的REN與海水溫度、TPM及PON均呈顯著相關(guān)性,RfeN則僅與海水溫度有顯著的相關(guān)關(guān)系,而與TPM和PON均無(wú)顯著相關(guān)性.文蛤的REP與海水溫度和TPM呈極顯著的相關(guān)關(guān)系,與POP無(wú)顯著相關(guān)性;RfeP則與海水溫度、TPM和POP含量均無(wú)顯著相關(guān)關(guān)系.
2.4 文蛤?qū)z取C、N、P的分配
由圖2可見(jiàn),文蛤在春季及秋季時(shí)的碳收支順序依次為糞便碳>呼吸碳>生長(zhǎng)碳,在夏季時(shí)為生長(zhǎng)碳>呼吸碳>糞便碳,冬季時(shí)為生長(zhǎng)碳>糞便碳>呼吸碳.呼吸碳及糞便碳的分配比例均在春季達(dá)到最高值,生長(zhǎng)碳所占比例則在夏季最高,在春季最低,且為負(fù)值.文蛤攝取氮元素用于生理過(guò)程的各組分比例依次為生長(zhǎng)氮>糞便氮>排泄氮,其中文蛤用于生長(zhǎng)的氮所占比例最高(65%),并在春季及夏季時(shí)較高,在春季時(shí)達(dá)到最高值,兩季節(jié)間差異不顯著;在秋季時(shí)較低,在冬季時(shí)達(dá)到最低值,且秋冬季兩者間差異不顯著.通過(guò)糞便形式排出的氮量占攝食氮的比例較高(33%),并且季節(jié)間差異顯著,在冬季時(shí)達(dá)到最高值,在春季時(shí)達(dá)到最低值.用于排泄消耗的氮量占攝食氮的比例較低(2%),在夏季時(shí)達(dá)到最高,秋季次之,春季較低,冬季最低,且春季和冬季間差異不顯著.文蛤攝食磷量用于生理過(guò)程的各組分比例在各季節(jié)中(除冬季外)依次為生長(zhǎng)磷>糞便磷>排泄磷,其中生長(zhǎng)磷所占比例超過(guò)50%,在春季達(dá)到最高值,夏季時(shí)最低,但與秋季和冬季間差異不顯著,文蛤通過(guò)糞便排出的磷所占比例在秋季時(shí)最高,在春季時(shí)最低,用于排泄消耗的磷所占攝食磷的比例在夏季最高,冬季最低,且季節(jié)間變化顯著.
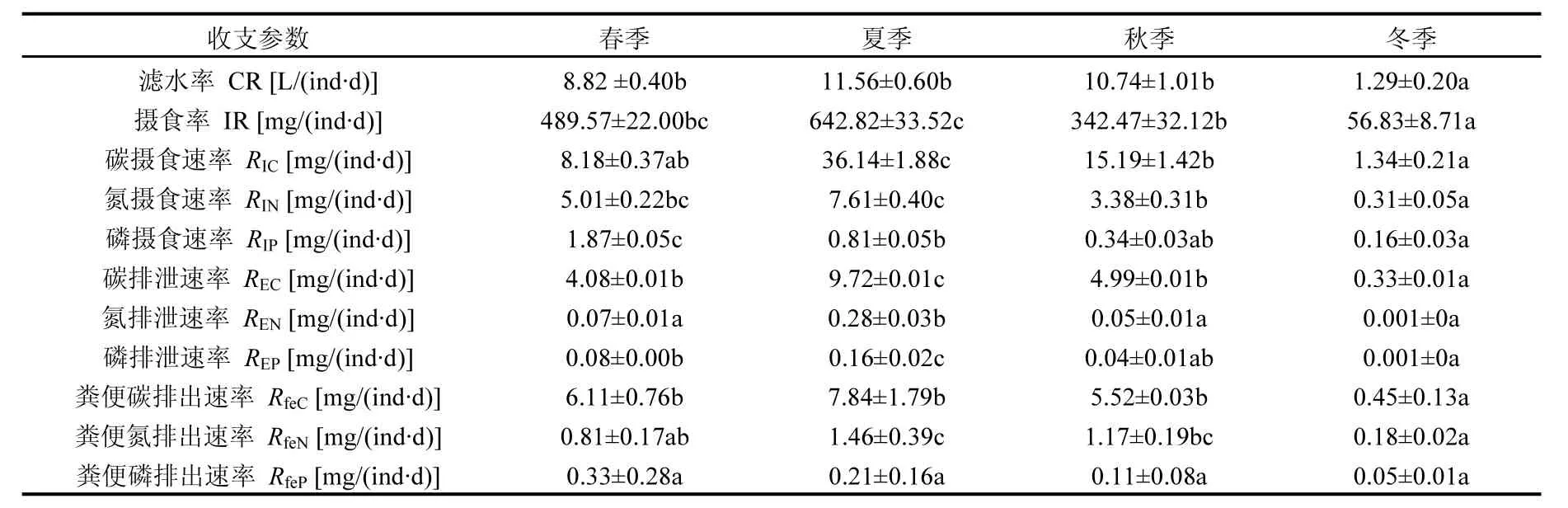
表1 文蛤CR、IR、C、N、P收支參數(shù)的季節(jié)變化Table 1 Clearance rate, ingestion rate, carbon, nitrogen, and phosphorus parameters of Meretrix meretrix in different seasons (mean±SE)

表2 文蛤生理生態(tài)學(xué)參數(shù)與環(huán)境因子的相關(guān)系數(shù)Table 2 Correlation coefficients between physio-ecological parameters of Meretrix meretrix and environmental variables
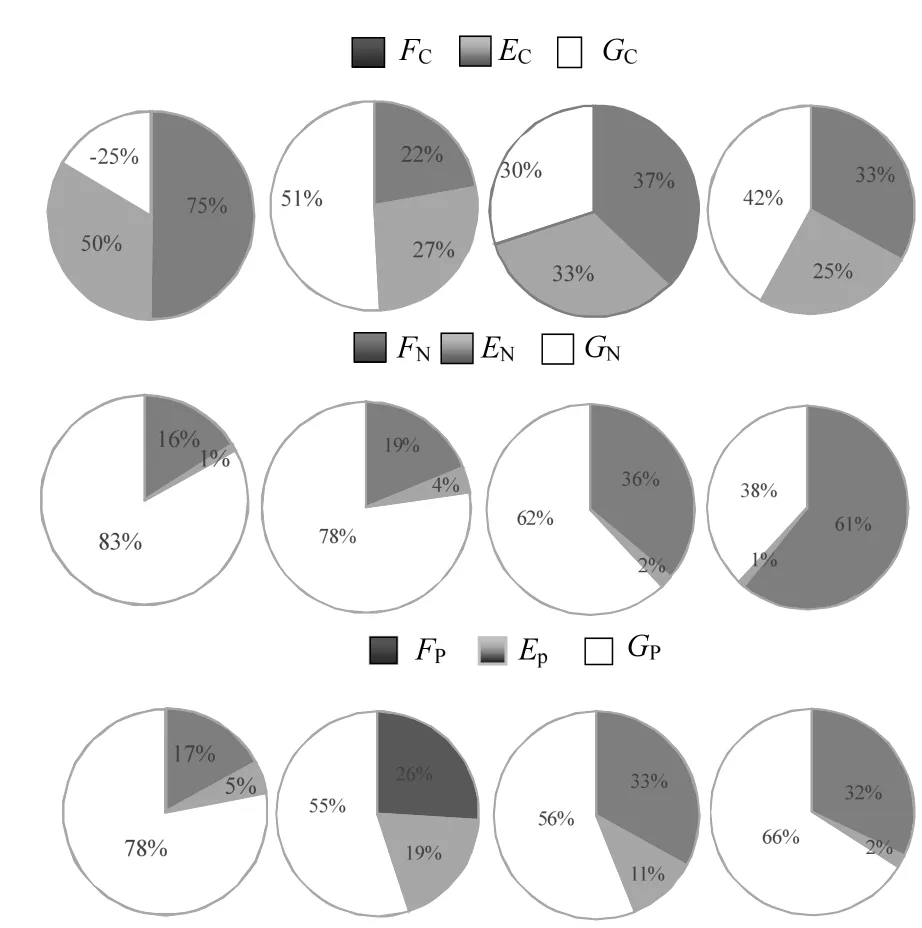
圖2 文蛤攝取的C、N、P元素在各生理生態(tài)過(guò)程中配給比例的季節(jié)變化Fig.2 Carbon, nitrogen, and phosphorus allocation into different physio-ecological process of Meretrix meretrix
2.5 文蛤的SFGC、SFGN、SFGP
由圖3可見(jiàn),文蛤的SFGC呈現(xiàn)明顯的季節(jié)變化,春季時(shí)最低,且為負(fù)值[-2.11mg C/(ind·d)],夏季時(shí)達(dá)到最高[18.59mg C/(ind?d)],秋季及冬季時(shí)均較低,分別為4.69和0.56mg C/(ind·d).相關(guān)性分析結(jié)果顯示(表2),文蛤的SFGC與海水溫度和POC含量呈極顯著的相關(guān)關(guān)系,與TPM無(wú)顯著相關(guān)性.
文蛤的SFGN在春季和夏季時(shí)較高,分別為4.13 和5.87mg N/(ind·d),且兩季節(jié)間差異不顯著,秋季時(shí)較低,冬季時(shí)達(dá)到最低值,僅為0.13mg N/(ind·d).相關(guān)性分析結(jié)果顯示(表2),文蛤的SFGN與海水溫度、TPM及PON含量均呈極顯著的相關(guān)關(guān)系.
文蛤的SFGP呈現(xiàn)明顯的季節(jié)變化特征(見(jiàn)圖3),春季時(shí)達(dá)到峰值1.45mg P/(ind·d),冬季最低,僅為0.11mg P/(ind·d),夏季及秋季時(shí)均較低,分別為0.46 和0.20mg P/(ind·d),且夏季、秋季及冬季間差異不顯著.相關(guān)性分析結(jié)果表明(表2),文蛤的SFGP與TPM及POP呈顯著相關(guān)性,而與海水溫度相關(guān)性不顯著.
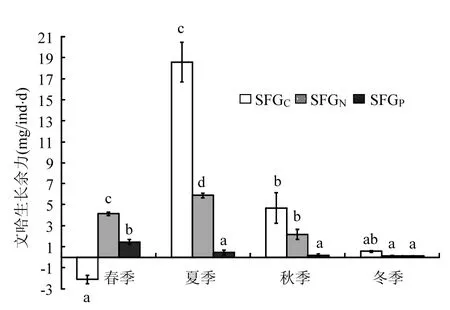
圖3 文蛤C、N、P元素生長(zhǎng)余力的季節(jié)變化Fig.3 Seasonal variations of carbon, nitrogen, and phosphorus scope for growth (SFG) of Meretrix meretrix
2.6 文蛤的KC、KN、KP
文蛤的KC1和KC2呈現(xiàn)明顯的季節(jié)性(圖4),即均在夏季達(dá)到最高值,春季達(dá)到最低值,且為負(fù)值.相關(guān)分析結(jié)果顯示(表2),文蛤的KC1和KC2與海水中POC濃度均呈顯著的相關(guān)關(guān)系,與海水溫度和TPM濃度均無(wú)顯著相關(guān)性.
由圖4可知,文蛤的KN1季節(jié)變化顯著,在春季時(shí)達(dá)到最高,夏季時(shí)較高,且兩季節(jié)間差異不顯著,秋季較低,冬季時(shí)最低,且秋冬季節(jié)間差異不顯著.文蛤的KN2均較高,且季節(jié)間變化不顯著.相關(guān)分析結(jié)果顯示(見(jiàn)表2),文蛤的KN1與海水溫度、TPM和PON濃度均呈顯著相關(guān)性,而KN2與海水溫度、TPM和PON濃度均無(wú)顯著相關(guān)性.
如圖4所示,文蛤的KP1也呈現(xiàn)明顯的季節(jié)變化,即文蛤的KP1在春季時(shí)最高(78%),夏季時(shí)最低(55%),但與秋季及冬季間差異不顯著.文蛤的KP2在春季最高,夏季最低.文蛤的KP1、KP2均與POP濃度呈顯著的正相關(guān)關(guān)系;均與TPM濃度無(wú)顯著相關(guān)性;KP2與海水溫度呈極顯著的負(fù)相關(guān)關(guān)系,而KP1則與溫度關(guān)系不顯著.
實(shí)驗(yàn)結(jié)果表明,文蛤的KC2、KN2、KP2年均值分別為13%、97%及85%.即基本表現(xiàn)為KN2>KP2>KC2.
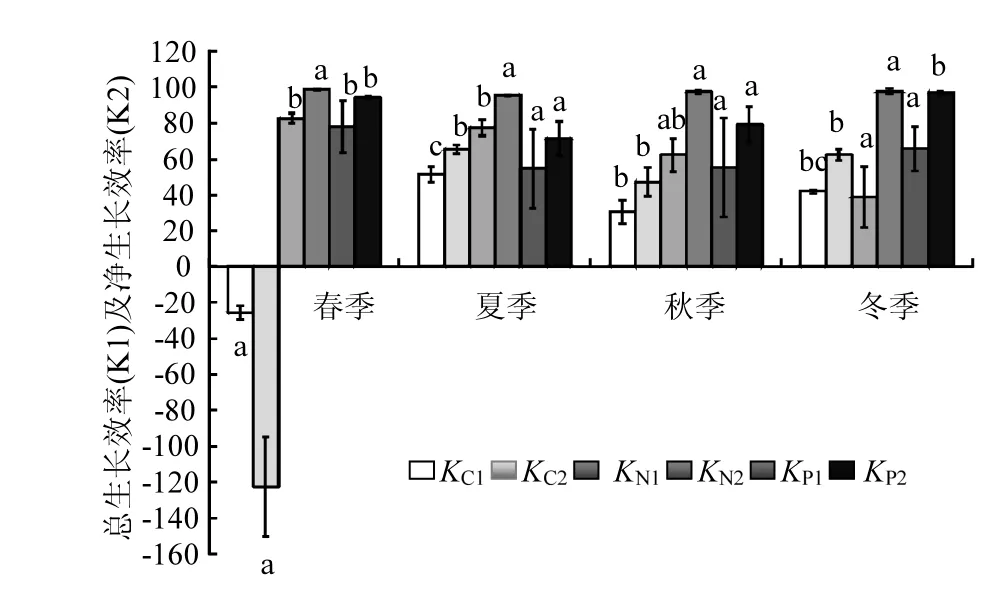
圖4 文蛤?qū)、N、P元素總生長(zhǎng)效率及凈生長(zhǎng)效率的季節(jié)變化Fig.4 Seasonal variations of total growth efficiency and net growth efficiency of Meretrix meretrix
3 討論
3.1 文蛤CR及IR的季節(jié)變化
研究表明,雙殼貝類(lèi)CR和IR主要受溫度等環(huán)境因子的影響,在適宜的溫度范圍內(nèi),其CR和IR隨溫度升高而上升,但超出適宜溫度范圍時(shí)則出現(xiàn)下降[22-26].本研究結(jié)果表明,文蛤生長(zhǎng)的適宜水溫為5.5~32℃,最適水溫為15~27℃[27].在本研究中,春季、夏季及秋季時(shí)海水溫度分別為12、17和28℃,因此實(shí)驗(yàn)文蛤處于較適宜的生活環(huán)境中,而水溫的升高使得海水粘滯性變小且水中生物餌料量也更加豐富,促使攝食生理活動(dòng)處于較高水平,從而表現(xiàn)出較高的CR和IR,并在夏季達(dá)到最高值.冬季時(shí),由于海水溫度降至最低值,且水溫超出了文蛤的適溫范圍,從而導(dǎo)致文蛤的濾水和攝食能力減弱.這與其他學(xué)者的研究結(jié)果基本一致[21,28-29].
3.2 文蛤的SFGC、SFGN、SFGP
研究發(fā)現(xiàn),貝類(lèi)的SFG作為其體內(nèi)能量供需機(jī)制可以反映出環(huán)境因子和生理活動(dòng)對(duì)其生長(zhǎng)的瞬間影響,同時(shí)周年實(shí)驗(yàn)結(jié)果也證明生長(zhǎng)余力和實(shí)際測(cè)量的生長(zhǎng)狀況相一致[19].通過(guò)表3可以看出,菲律賓蛤仔個(gè)體的SFGC、SFGN、SFGP均高于文蛤和四角蛤蜊.本文推測(cè)造成貝類(lèi)不同種類(lèi)間特定SFG的差異一方面可能是由于實(shí)驗(yàn)海區(qū)環(huán)境條件不同造成的.資料表明[11,14-15],菲律賓蛤仔實(shí)驗(yàn)個(gè)體所在的莊河海域與文蛤和四角蛤蜊實(shí)驗(yàn)個(gè)體所在的雙臺(tái)子河口海域環(huán)境條件差異比較明顯,兩者的溫度年均值分別為11.5和14.2℃, 鹽度分別為31.3和20.5,TPM含量分別為14.3和46.8mg/L.因此,實(shí)驗(yàn)海域的環(huán)境條件差異,從而造成菲律賓蛤仔的生長(zhǎng)余力高于文蛤和四角蛤蜊.另一方面,不同種類(lèi)間特定生長(zhǎng)余力的差異可能是由于實(shí)驗(yàn)貝類(lèi)種類(lèi)不同.由于實(shí)驗(yàn)貝類(lèi)文蛤和四角蛤蜊均處于雙臺(tái)子河口海域,所處環(huán)境條件基本一致,但是文蛤和四角蛤蜊個(gè)體規(guī)格不同(文蛤殼高平均為41.48mm,軟體干重為1.02g;四角蛤蜊殼高平均為28.74mm,軟體干重為0.29g).因此,實(shí)驗(yàn)個(gè)體不同造成了兩者SFG的差異.
SFG反映了貝類(lèi)獲取和利用能量物質(zhì)的一種平衡,其受貝類(lèi)的個(gè)體大小、繁殖時(shí)期、水溫以及餌料條件等因素的影響[12,26,30-32].研究發(fā)現(xiàn),櫛孔扇貝和長(zhǎng)牡蠣的SFGC在夏季時(shí)出現(xiàn)負(fù)值主要與低餌料濃度有關(guān)[12,16,30],這可能與夏季桑溝灣溫度偏高(26.7℃)、餌料濃度偏低(5.34mg/L)有關(guān),海水溫度超過(guò)了貝類(lèi)適宜的生長(zhǎng)溫度,其自身生理代謝活動(dòng)降低[16].菲律賓蛤仔的SFG隨餌料濃度的增加而顯著增加[31];四角蛤蜊的SFGC與POC濃度呈顯著的相關(guān)關(guān)系[14].本研究結(jié)果顯示,雙臺(tái)子河口文蛤的SFGC也與該海域水體中的POC呈顯著的正相關(guān)關(guān)系.這說(shuō)明餌料濃度是影響該河口海域文蛤生長(zhǎng)的重要因子之一.文蛤的SFGC在春季時(shí)為負(fù)值,這種現(xiàn)象主要與該季節(jié)海水中POC含量較低有關(guān).如前所述,在春季時(shí),文蛤處于比較適宜的生活環(huán)境,但海水中POC含量較低,導(dǎo)致文蛤?qū)τ袡C(jī)碳源的攝食率較低,而此時(shí)呼吸速率卻隨著溫度升高而迅速增加,即要消耗較大部分的碳量來(lái)維持自身的代謝,進(jìn)而導(dǎo)致SFGC在春季時(shí)出現(xiàn)負(fù)值.本研究結(jié)果顯示,文蛤的SFGC和SFGN均與海水溫度有顯著的相關(guān)關(guān)系.通過(guò)上述分析可以看出,在溫度適宜的季節(jié)(如春季)時(shí),海區(qū)中餌料濃度可能是影響文蛤生長(zhǎng)的主要因子;在餌料充足季節(jié)(如夏季)時(shí),溫度的變化會(huì)對(duì)文蛤的生長(zhǎng)產(chǎn)生較大的影響.總之,在河口海域中文蛤的生長(zhǎng)主要受到溫度及餌料濃度共同作用的影響.另外,盡管雙臺(tái)子河口海域文蛤的碳生長(zhǎng)在春季出現(xiàn)負(fù)值,但對(duì)整個(gè)雙臺(tái)子河口海域而言,文蛤在春季屬于碳源,并且從周年平均水平上來(lái)說(shuō),碳的生長(zhǎng)能為正,屬于元素的匯,通過(guò)文蛤等濾水性貝類(lèi)的攝食代謝等生理活動(dòng),貝類(lèi)完成了碳源向碳匯轉(zhuǎn)換的過(guò)程.
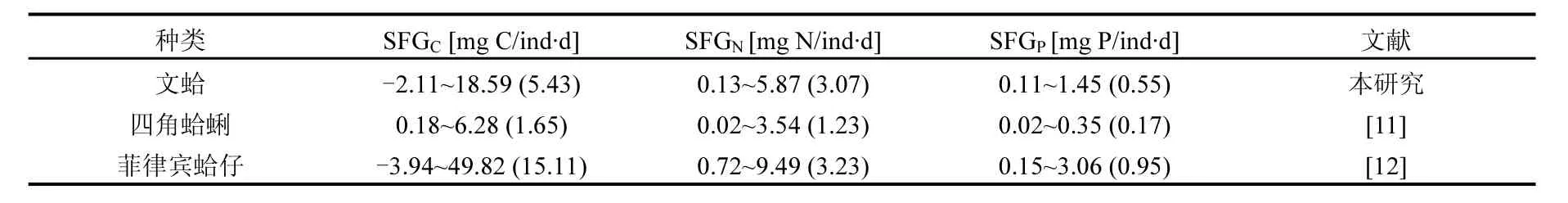
表3 文蛤與其他貝類(lèi)生長(zhǎng)余力比較Table 3 Comparison of scope for growth of Meretrix meretrix and other bivalves
3.3 文蛤的C、N、P生長(zhǎng)效率
研究表明,生長(zhǎng)效率也常被用來(lái)描述貝類(lèi)生長(zhǎng)狀況,但由于測(cè)定條件的差異,變化范圍較大.雙殼貝類(lèi)的K1變化范圍為2%~54%, K2變化范圍為3%~86%[12-13,19,30],本研究中文蛤的K1和K2大多數(shù)數(shù)值也處于這一范圍內(nèi).Smaal等[19]指出貽貝的KC2在秋季及冬季出現(xiàn)負(fù)值,主要是由于該季節(jié)餌料缺乏造成的.與上述結(jié)果類(lèi)似,在本研究中文蛤的KC1和KC2在春季時(shí)出現(xiàn)負(fù)值,也是由于該季節(jié)海水中POC含量較低,造成對(duì)C元素的攝食率較低,其攝取的有機(jī)碳源絕大部分通過(guò)自身的呼吸作用被消耗掉,從而導(dǎo)致用于生長(zhǎng)的碳量為負(fù)值.冬季時(shí)雙臺(tái)子河口海域海水溫度降至最低,文蛤生理代謝活動(dòng)緩慢,文蛤?qū)元素的攝食速率也較低,但碳呼吸消耗速率和糞便碳排出速率均降至全年最低值,因此,攝取的碳量仍有部分用于自身的生長(zhǎng).
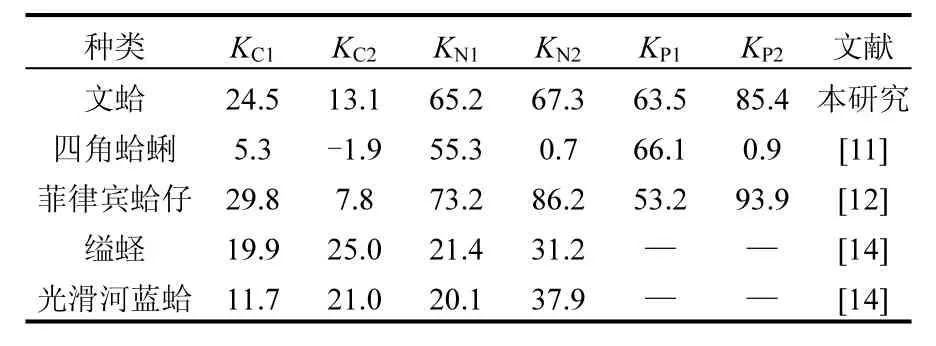
表4 文蛤與其他貝類(lèi)生長(zhǎng)效率比較Table 4 Comparison of growth efficiency of Meretrix meretrix and other bivalves
貝類(lèi)的凈生長(zhǎng)效率越高,則表明其攝取的物質(zhì)中大部分用于自身的生長(zhǎng)和繁殖,僅有少部分用于維持代謝[19].本研究發(fā)現(xiàn),文蛤的KC2年均值最低,為13%,而KN2、KP2年均值則很高,分別為97%和85%.即基本表現(xiàn)為KN2>KP2>KC2,這與貽貝[19-20]、櫛孔扇貝[30]、四角蛤蜊[14,18]、縊蟶和光滑河藍(lán)蛤[17]等雙殼貝類(lèi)基本一致(表4),說(shuō)明文蛤等濾食性貝類(lèi)更趨向于對(duì)N、P元素的富集,即與C元素相比,攝取的氮量和磷量大部分用于其自身生長(zhǎng)和繁殖.
3.4 文蛤生理生態(tài)過(guò)程對(duì)河口生態(tài)系統(tǒng)的影響
濾食性貝類(lèi)是C、N、P主要生源要素循環(huán)的重要媒介之一,在海洋生態(tài)系統(tǒng)的物質(zhì)循環(huán)中發(fā)揮著重要作用.例如,在菲律賓蛤仔養(yǎng)殖區(qū),生產(chǎn)6000t的蛤仔,可從水體中分別攝取約137t顆粒氮及19t顆粒磷,約91t的總?cè)芙獾?2t的總?cè)芙饬字匦逻M(jìn)入生態(tài)系統(tǒng)循環(huán)中,且有16t的氮及0.9t的磷通過(guò)收獲的方式直接從養(yǎng)殖生態(tài)系統(tǒng)物質(zhì)循環(huán)中輸出[33].四角蛤蜊每年從雙臺(tái)子河口海水中轉(zhuǎn)移約6710.3、7422.2、1378.4t的POC、PON和POP[14].作者調(diào)查發(fā)現(xiàn),2016年雙臺(tái)子河口海域?yàn)┩课母虻姆植济芏燃s為2.3個(gè)/m2,其中二齡以上的文蛤個(gè)體所占比例為81%,即1.9個(gè)/m2.根據(jù)上述相關(guān)參數(shù),本研究估算出每年每公頃雙臺(tái)子河口海域文蛤可過(guò)濾海水56.2t,顆粒有機(jī)物攝食量達(dá)2656.9kg,攝取的C、N、P總量分別為105.5、28.3和5.5kg,形成可移出的碳匯為37.7kg,同時(shí)用于生長(zhǎng)的N、P總量分別為21.3和3.8kg.通過(guò)分析可以看出,文蛤等濾食性貝類(lèi)通過(guò)攝食水體中的單細(xì)胞藻類(lèi)和顆粒有機(jī)物來(lái)滿(mǎn)足自身的生長(zhǎng)所需,攝食的能量主要通過(guò)呼吸作用和排泄代謝回到水體,部分通過(guò)糞便直接排除體外,作為生物沉積沉降到海底,沉降到海底的C、N、P元素部分通過(guò)生物再懸浮重回水體(圖5).濾食性貝類(lèi)增殖不僅增加經(jīng)濟(jì)效益,還可以有效地去除海水里的碳元素,并對(duì)河口海域的生態(tài)系統(tǒng)能量流動(dòng)和物質(zhì)循環(huán)產(chǎn)生較大影響.
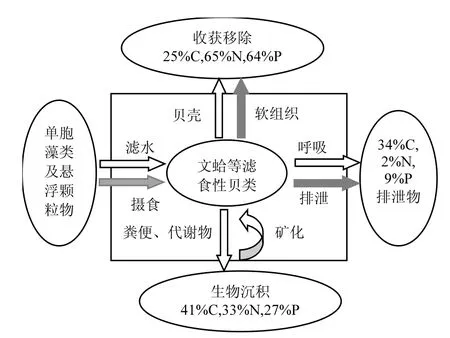
圖5 雙臺(tái)子河口文蛤?qū)、N、P生源要素?cái)z食和利用Fig.5 The ingestion and utilization of carbon, nitrogen,and phosphorus for Meretrix meretrix in Shuangtaizi estuary
4 結(jié)論
4.1 文蛤SFGC、SFGN、SFGP均呈現(xiàn)明顯的季節(jié)變化,即在夏季最高;但SFGC在春季時(shí)最低,且為負(fù)值,SFGN及SFGP全年均為正值.
4.2 文蛤的C、N、P生長(zhǎng)效率季節(jié)變化顯著,且C、N、P凈生長(zhǎng)效率基本表現(xiàn)為KN2> KP2> KC2.
4.3 文蛤在春季及秋季時(shí)的C收支順序依次為糞便碳>呼吸碳>生長(zhǎng)碳,在夏季時(shí)為生長(zhǎng)碳>呼吸碳>糞便碳,冬季時(shí)為生長(zhǎng)碳>糞便碳>呼吸碳.文蛤N收支順序?yàn)樯L(zhǎng)氮>糞便氮>排泄氮.文蛤P收支順序?yàn)?除冬季外)依次為生長(zhǎng)磷>糞便磷>排泄磷.
4.4 文蛤等濾食性貝類(lèi)更趨向于對(duì)N、P元素的富集,即與C元素相比,攝取的氮量和磷量大部分用于其自身生長(zhǎng)和繁殖.
4.5 文蛤資源增殖不僅增加經(jīng)濟(jì)效益,還可以有效地去除海水里的碳元素,并對(duì)河口海域的生態(tài)系統(tǒng)能量流動(dòng)和物質(zhì)循環(huán)產(chǎn)生較大影響.
[1] Cockcroft A C. Nitrogen excretion by the surf zone bivalvesDanax SerraandD. Sordulus[J]. Marine Ecology Progress Series,1990,60:57-65.
[2] 周 毅,楊紅生,張福綏.海水雙殼貝類(lèi)的N、P排泄及其生態(tài)效應(yīng) [J]. 中國(guó)水產(chǎn)科學(xué), 2003,10(2):165-168.
[3] 王曉宇,周 毅,楊紅生.膠州灣菲律賓蛤仔(Ruditapes philippinarum)呼吸排泄作用的現(xiàn)場(chǎng)研究 [J]. 海洋與湖沼,2011,42(5):722-727.
[4] Prins T C, Smaal A C. The role of the blue musselMytilus edulisin the cycling of nutrients in the Oosterschelde estuary (The Netherlands) [J]. Hydrobiologia, 1994,282-283:413-429.
[5] Zhou Y, Yang H S, Zhang T, et al. Density-dependent effects on seston dynamics and rates of filtering and biodeposition of the suspension-cultured scallopChlamys farreriin a eutrophic bay(northern China): an experimental study in semi-in situ flowthrough systems [J]. Journal of Marine System, 2006,59:143-158.
[6] 臧家業(yè),趙晨英,劉 軍.乳山灣鄰近海域有機(jī)碳的分布與底界面過(guò)程 [J]. 中國(guó)環(huán)境科學(xué), 2017,37(3):1089-1102.
[7] 趙晨英,臧家業(yè),劉 軍,等.黃渤海氮磷營(yíng)養(yǎng)鹽的分布、收支與生態(tài)環(huán)境效應(yīng) [J]. 中國(guó)環(huán)境科學(xué), 2016,36(7):2115-2127.
[8] 許春玲,李緒錄,林 梵.深圳灣及鄰近沿岸沉積物中碳、氮和磷的分布 [J]. 中國(guó)環(huán)境科學(xué), 2017,37(7):2691-2698.
[9] 蔣 紅,崔 毅,陳碧鵑,等.乳山灣菲律賓蛤仔可溶性氮、磷排泄及其與溫度的關(guān)系 [J]. 中國(guó)水產(chǎn)科學(xué), 2006,13(2):237-242.
[10] 王如才,王昭萍,張建中.海水貝類(lèi)養(yǎng)殖學(xué) [M]. 青島:青島海洋大學(xué)出版社, 1993:322-324.
[11] 張安國(guó),袁秀堂,侯文久,等.文蛤的生物沉積和呼吸排泄過(guò)程及其在雙臺(tái)子河口水層-底棲系統(tǒng)中的耦合作用 [J]. 生態(tài)學(xué)報(bào),2014,34(22):6573-6582.
[12] 周 毅,楊紅生,張 濤,等.四十里灣櫛孔扇貝的生長(zhǎng)余力和C、N、P元素收支 [J]. 中國(guó)水產(chǎn)科學(xué), 2002,9(2):161-166.
[13] 柴雪良,張炯明,方 軍,等.樂(lè)清灣、三門(mén)灣主要濾食性養(yǎng)殖貝類(lèi)碳收支的研究 [J]. 上海水產(chǎn)大學(xué)學(xué)報(bào), 2006,15(1):52-58.
[14] Zhang A G, Yuan X T, Hou W J, et al. Carbon, nitrogen, and phosphorus budgets of the surf clamMactra veneriformis(Reeve)based on a field study in the Shuangtaizi Estuary, Bohai Sea of China [J]. Journal of Shellfish Research, 2013,32(2):275-284.
[15] 張升利,張安國(guó),袁秀堂,等.底播增殖菲律賓蛤仔碳、氮、磷收支[J]. 應(yīng)用生態(tài)學(xué)報(bào), 2015,26(4):1244-1252.
[16] 牛亞麗.桑溝灣濾食性貝類(lèi)碳、氮、磷、硅元素收支的季節(jié)變化研究 [D]. 舟山:浙江海洋學(xué)院, 2014.
[17] 呂昊澤.縊蟶、光滑河藍(lán)蛤和河蜆對(duì)鹽度的適應(yīng)性及碳、氮收支研究 [D]. 上海:上海海洋大學(xué), 2014.
[18] Hiwatari T, Kohata K, Iijima A. Nitrogen budget of the bivalveMactra veneriformis, and its significance in benthic-pelagic systems in the Sanbanse area of Tokyo bay [J]. Estuarine, Coastal and Shelf Science, 2002,55:299-308.
[19] Smaal A C, Vonck A P M A. Seasonal variation in C, N, and P budgets and tissue composition of the musselMytilus edulis[J].Marine Ecology Progress Series, 1997,153:167-179.
[20] Jansen H M, Strand Q, Verdegem M, et al. Accumulation, release and turnover of nutrients (C-N-P-Si) by the blue musselMytilus edulisunder oligotrophic conditions [J]. Journal of Experimental Marine Biology and Ecology, 2012,416-417:185-195.
[21] 范建勛.文蛤能量代謝的研究 [D]. 寧波:寧波大學(xué), 2010.
[22] Kuang S, Fang J, Sun H, et al. Seasonal studies of filtration rate and absorption efficiency in the scallopChlamys farreri[J].Journal of Shellfish Research, 1997,16:39-45.
[23] 常亞青,王子臣.皺紋盤(pán)鮑的個(gè)體能量收支 [J]. 應(yīng)用生態(tài)學(xué)報(bào),1998,9(5):511-516.
[24] 董 波,薛欽昭,李 軍.溫度對(duì)菲律賓蛤仔濾食率、清濾率和吸收率的影響 [J]. 海洋水產(chǎn)研究, 2000,21(1):37-42.
[25] 栗志民,劉志剛,鄧海東.溫度和鹽度對(duì)企鵝珍珠貝清濾率、濾食率、吸收率的影響 [J]. 水產(chǎn)學(xué)報(bào), 2011,35(1):96-103.
[26] Guzma′n-Agu¨ero J E, Nieves-Soto M, Hurtado M A, et al.Feeding physiology and scope for growth of the oysterCrassostrea corteziensis(Hertlein, 1951) acclimated to different conditions of temperature and salinity [J]. Aquaculture International, 2013,21:283-297.
[27] 赫崇波,陳洪大.灘涂養(yǎng)殖文蛤生長(zhǎng)和生態(tài)習(xí)性的初步研究 [J].水產(chǎn)科學(xué), 1997,16(5):17-20.
[28] Tang B J, Liu B Z, Yang H S, et al. Oxygen consumption and ammonia-N excretion ofMeretrix meretrixin different temperature and salinity [J]. Chinese Journal of Oceanology and Limnology, 2005,23(4):469-474.
[29] 范建勛,林志華,肖國(guó)強(qiáng),等.饑餓對(duì)3種不同規(guī)格文蛤耗氧率和排氨率的影響 [J]. 海洋科學(xué), 2009,33(10):73-76.
[30] 周 毅,楊紅生,張福綏.櫛孔扇貝生理生態(tài)學(xué)特征的實(shí)驗(yàn)研究[J]. 應(yīng)用生態(tài)學(xué)報(bào), 2003,14(3):227-233.
[31] 董 波,李 軍,王海燕,等.不同溫度與餌料濃度下菲律賓蛤仔的能量收支 [J]. 中國(guó)水產(chǎn)科學(xué), 2003,10(5):398-403.
[32] Helson J G, Gardner J P A. Variation in scope for growth: a test of food limitation among intertidal mussels [J]. Hydrobiologia,2007,586:373-392.
[33] Nizzoli D, Bartoli M, Viaroli P. Nitrogen and phosphorous budgets during a farming cycle of the Manila clamRuditapes philippinarum: Anin situexperiment [J]. Aquaculture, 2006,261:98-108.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
