紫紅紅球菌9α–羥化酶loop對(duì)酶活性的影響
2018-02-28 06:32:40申雁冰王九彬
天津科技大學(xué)學(xué)報(bào) 2018年1期
劉 揚(yáng),申雁冰,王九彬,王 敏
(工業(yè)發(fā)酵微生物教育部重點(diǎn)實(shí)驗(yàn)室,天津市工業(yè)微生物重點(diǎn)實(shí)驗(yàn)室,天津科技大學(xué)生物工程學(xué)院,天津 300457)
雄甾–4–烯–3,17–二酮(androst-4-ene-3,17-dione,AD)是合成甾體激素類藥物的關(guān)鍵中間體,它的 9α羥基化產(chǎn)物9α–羥基雄甾–4–烯–3,17–二酮(9α-hydroxyandrost-4-ene-3,17-dione,9α-OH-AD)是生產(chǎn)9α位上有鹵素的多種皮質(zhì)激素的重要前體[1]. 能夠進(jìn)行 9α–羥化的微生物有很多,目前應(yīng)用較多的菌種是紅球菌(Rhodococcus sp.)和分枝桿菌(Mycobacterium sp.).3–甾 酮 –9α–羥 化 酶 (3-ketosteroid-9α-hydroxylase,KSHAB)負(fù)責(zé)將 AD 轉(zhuǎn)化生成 9α-OHAD,由末端加氧酶(KSHA)與鐵硫還原酶(KSHB)雙組分構(gòu)成.
羥化酶的加氧酶組分 KSHA在底物進(jìn)出酶活性中心出口處,存在一個(gè) loop,像一個(gè)“開關(guān)”,將底物阻擋在活性中心通道外[2].在本課題組前期對(duì)分枝桿菌轉(zhuǎn)化植物甾醇生成AD的研究中發(fā)現(xiàn),分枝桿菌(Mycobacterium neoaurum)TCCC11028的 3–甾酮–Δ1–脫氫酶靠近底物活性中心處的 loop上的 Ser138突變成性質(zhì)不同的氨基酸 Leu138后,顯著影響了該酶對(duì) AD的活性[3].已有文獻(xiàn)證明柔性 loop的開關(guān)運(yùn)動(dòng)是調(diào)控酶催化作用的關(guān)鍵因素,比如磷酸丙糖異構(gòu)酶與脂肪酶的晶體結(jié)構(gòu)研究表明,柔性 loop 作為“開關(guān)”調(diào)節(jié)底物進(jìn)入活性位點(diǎn),進(jìn)而影響酶對(duì)底物的活性[4–6].
盡管目前有2個(gè)KSHA的蛋白晶體結(jié)構(gòu)得到解析,但是KSHA的loop與酶活性之間的關(guān)系尚不清楚.本研究以已知晶體結(jié)構(gòu)的紫紅紅球菌(Rhodococcus rhodochrous)DSM43269的 KSHA5 loop為研究對(duì)象,應(yīng)用定點(diǎn)突變技術(shù)探究loop對(duì)酶活性的影響,解析各氨基酸對(duì) loop開關(guān)運(yùn)動(dòng)、調(diào)節(jié)底物通道的重要性,研究成果加深對(duì) 9α–羥化酶的認(rèn)識(shí)有著重要意義,同時(shí)也為進(jìn)一步研究其催化機(jī)理、改造酶蛋白活性提供理論依據(jù).
1 材料與方法
1.1 材料
1.1.1 菌株、質(zhì)粒與培養(yǎng)基
2.3.3 5-HT 3RA的應(yīng)用情況 本研究中患者應(yīng)用的 5-HT3RA 包括 Tro、Pal和 Ond。“指南”推薦 Tro 只在第1天靜脈應(yīng)用或口服5mg。有59例(53.15%)連續(xù)多日應(yīng)用Tro,療程過長(zhǎng)。應(yīng)用Pal存在的主要問題與Tro一致,有11例(9.91%)應(yīng)用Pal療程過長(zhǎng)。有4例將Pal與Tro重復(fù)應(yīng)用,因藥物與受體的結(jié)合存在飽和現(xiàn)象,重復(fù)用藥并不能增強(qiáng)療效,反而會(huì)增大不良反應(yīng)的發(fā)生率,兩藥重復(fù)應(yīng)用為不合理用藥。Ond應(yīng)用主要存在超致吐級(jí)別應(yīng)用、給藥劑量偏小和療程不合理。Ond為短效的5-HT3RA,應(yīng)用Ond的例數(shù)較Pal和Tro少。見表6。
用限制性內(nèi)切酶 NcoⅠ和 HindⅢ酶切質(zhì)粒,鑒定重組質(zhì)粒的正確性,如圖2所示.
質(zhì)粒 pGEM-T Easy購(gòu)自 Promega公司,質(zhì)粒pET-28a(+)由本實(shí)驗(yàn)室保存.
LB 培養(yǎng)基(g/L):蛋白胨 10,酵母提取物 5,NaCl 10,pH 7.5.固體培養(yǎng)基在此基礎(chǔ)上添加2%,的瓊脂粉.
1.1.2 試劑
黑龍江省龍江電器集團(tuán)有限公司成立于1996年,為中小型企業(yè),現(xiàn)有職工370人。龍江電器集團(tuán)有限公司自2002年起開始推行企務(wù)公開工作。近年來,該公司以落實(shí)“三個(gè)代表”重要思想,貫徹黨的依靠方針為指導(dǎo),著力在規(guī)范、鞏固、深化、創(chuàng)新、實(shí)效上下功夫。隨著企務(wù)公開的不斷開展,企業(yè)民主管理渠道不斷拓寬,廣大職工建功立業(yè)、奉獻(xiàn)企業(yè)的熱情不斷高漲,該公司保持長(zhǎng)周期安全穩(wěn)定可持續(xù)發(fā)展。該公司先后榮獲國(guó)家“五一勞動(dòng)獎(jiǎng)?wù)隆薄⑹ 皬S務(wù)公開民主管理先進(jìn)單位”、省、市“文明單位”等多項(xiàng)榮譽(yù)。
限制性內(nèi)切酶(NcoⅠ、HindⅢ和 DpnⅠ),NEB公司;蛋白質(zhì) Marker,北京全式金公司;TaKaRa LA TaqTMwith GC Buffer、DNA Marker,Takara 公司;KOD Plus,Toyobo公司;T4連接酶,Promega公司;質(zhì)粒小提試劑盒、膠回收試劑盒、基因組提取試劑盒,Omega公司;其他藥品和試劑均為分析純.
1.2 KSHA氨基酸比對(duì)與蛋白結(jié)構(gòu)
KSHAB以NADH為輔酶,將來自NADH的電子最終傳遞到 O2,使底物被羥化.可以通過檢測(cè)340,nm下 NADH被氧化時(shí)吸光度的變化表征KSHA5及其各突變體的活性,摩爾吸光系數(shù) ε采用6.22,mmol/(L·cm).酶活力單位定義為 1,min氧化1,nmol NADH所需要的酶量.KSH測(cè)定體系為含終濃度為 105,μmol/L的 NADH 和 250,μmol/L的底物AD(溶于 100%,異丙醇)的 200,μL 酶液.測(cè)定時(shí),先向酶標(biāo)板中加入198,μL蛋白酶液(質(zhì)量濃度為 60,μg/mL)和 1,μL NADH 溶液(母液濃度為 21,mmol/L),最后加入 1,μL 的底物(母液濃度為 50,mmol/L),啟動(dòng)反應(yīng),在酶標(biāo)儀上(33,℃)實(shí)時(shí)監(jiān)測(cè)反應(yīng).
KSHA5無底物的3D結(jié)構(gòu)(4QDF)與有底物AD的 3D結(jié)構(gòu)(4QDC)從 Protein Data Bank數(shù)據(jù)庫(kù)下載,用Discovery Studio 2.5軟件分析有無底物存在時(shí)KSHA5的結(jié)構(gòu).
1.3 表達(dá)載體的構(gòu)建
根據(jù) KSHAB的末端加氧酶 kshA5與鐵硫還原酶 kshB基因設(shè)計(jì)特異性引物,引入 NcoⅠ和 HindⅢ兩個(gè)酶切位點(diǎn),kshA5的上、下游引物分別為kshA5-TF1:5′-CCATGGTGTCCATCGACACCGCACGG-3′,kshA5-T-R1:5′-AAGCTT GGGGGTCGCGGTGGAGC C-3′;kshB 的上、下游引物分別為 kshB-T-F1:5′-CCATGGTGACAGCCGTCCAGGCACC-3′,kshB-TR1:5′-AAGCTT GAACTCGATGCGCACGTGGT-3′(其中標(biāo)注下劃線的堿基為添加的酶切位點(diǎn)).
另外,通過不同類型的KSHA的比對(duì),還發(fā)現(xiàn)在loop上存在兩個(gè)非常保守的氨基酸,R217與 D219,用軟件Discovery Studio 2.5分析了兩者的相互作用,發(fā)現(xiàn)它們之間存在著鹽橋相互作用,這可能起到固定loop位置的作用,使loop能更貼合到底物入口處.因此,將兩者之一進(jìn)行突變,把 R217變成 A217,使鹽橋作用喪失,讓loop比較松弛地靠近入口,使入口變大,考察其對(duì)活性的影響.
在4個(gè)組別溫度升高的第1天、第3天和第10天,每個(gè)組別的每個(gè)重復(fù)中隨機(jī)選擇1只雞,選擇使用數(shù)字體溫計(jì)測(cè)量肉雞直腸溫度。同時(shí),在同一時(shí)間內(nèi),采集該重復(fù)的另1只雞迅速使其安樂死,在3 min內(nèi)取出下丘腦組織放入速凍管中-80 ℃保存,20 d內(nèi)測(cè)定下丘腦中熱休克蛋白70的濃度[2]。
1.4 loop上氨基酸的定點(diǎn)突變
以重組表達(dá)載體pET28-kshA5為模板,采用全質(zhì)粒 PCR的方法進(jìn)行突變子的構(gòu)建,R217的上、下游突變引物分別為 TBR217A-F:5′-ACCGGTGCGGAGG ACGTCATCTCCG-3′,TBR217A-R:5′-GTCCTCCGCA CCGGTCGAGTGCAT-3′;T224的上、下游突變引物分別為 TBT224V-F:5′-TCCGGCGTGAACTACGACG ACCCCAAC-3′,TBT224V-R:5′-GTAGTTCACGCCGG AGATGACGTCC-3′;N225的上、下游突變引物分別為 TBN225L-F:5′-GGCACCCTGTACGACGACCCCA ACG-3′,TBN225L-R:5′-GTCGTACAGGGTGCCGGA GATGACG-3′;Y226的上、下游突變引物分別為TBY226F-F:5′-ACCAACTTTGACGACCCCAACGCC G-3′,TBY226F-R:5′-GTCGTCAAAGTTGGTGCCGGA GATGACG-3′;D227的上、下游突變引物分別為TBD227S-F:5′-AACTACAGCGACCCCAACGCCGAA C-3′,TBD227S-R:5′-GGGGTCGCTGTAGTTGGTGCC GGAGATG-3′;D228的上、下游突變引物分別為TBD228S-F:5′-TACGACAGCCCCAACGCCGAACTG C-3′,TBD228S-R:5′-GTTGGGGCTGTCGTAGTTGGT GCCGGAG-3′(其中斜體部分的堿基為突變位點(diǎn)).PCR 反應(yīng)體系(50,μL):pET28-KSHA5 模板 1,μL,10×buffer for KOD plus 5,μL,dNTP mixture 6,μL,KOD plus 1,μL,25,mmol/L MgSO43,μL,10,μmol/L上、下游特異性引物各 2,μL,ddH2O 補(bǔ)齊至50,μL.反應(yīng)條件:94,℃ 5,min;94,℃ 30,s,58,℃45,s,68,℃ 7,min,25 個(gè)循環(huán);68,℃ 10,min.反應(yīng)完畢后,直接向 PCR的反應(yīng)體系中添加限制性內(nèi)切酶DpnⅠ及其 buffer,37,℃消化模板 2,h,取 10,μL 消化液轉(zhuǎn)化到 E.coli DH5α 感受態(tài)細(xì)胞中.挑取陽(yáng)性克隆,測(cè)序正確后,轉(zhuǎn)化E.coli BL21感受態(tài)細(xì)胞,得到各突變體的工程菌.
1.5 蛋白的誘導(dǎo)表達(dá)與純化
挑取陽(yáng)性克隆單菌落接種于 5,mL含 50,μg/mL卡那霉素的LB液體培養(yǎng)基中,37,℃、200,r/min過夜培養(yǎng).取培養(yǎng)液按體積比 1∶50轉(zhuǎn)接于 50,mL含50,μg/mL 卡納霉素的 LB 液體培養(yǎng)基中,37,℃、200,r/min培養(yǎng),表達(dá) KSHA5(或其突變體)蛋白的菌液培養(yǎng)至 A600=0.4~0.6,表達(dá) KSHB蛋白的菌液長(zhǎng)至 A600=0.2,加入 IPTG 至終濃度 0.5,mmol/L,30,℃、200,r/min 繼續(xù)培養(yǎng) 24,h.
分別收集含KSHA5(或其突變體蛋白)與 KSHB的重組大腸桿菌菌體,加入破碎緩沖液(50,mmol/L Tris-HCl(pH 7.4),500,mmol/L NaCl,20,mmol/L 咪唑,0.1,mmol/L EDTA)徹底洗去培養(yǎng)基,再用破碎緩沖液重浮菌體,使各突變體細(xì)胞濃度保持一致.按體積比 1∶2取 KSHA5(或突變體)細(xì)胞懸浮液與KSHB細(xì)胞懸浮液,充分混勻,超聲破碎細(xì)胞17,min(功率 32%,,工作 5,s,間隔 5,s).蛋白純化的過程參見文獻(xiàn)[7].純化出的蛋白不需要透析(因?yàn)橥肝鲆矔?huì)使蛋白的活性受到損失),用 Bradford法定量總蛋白,并用軟件 Quantity One分析確定 KSHA與KSHB含量的比值,通過添加純 KSHB蛋白調(diào)整各突變蛋白與KSHB的含量比為4∶1.
1.6 蛋白活性測(cè)定
不同類型的KSHA蛋白的氨基酸序列用Clustal X比對(duì).
2 結(jié)果與分析
2.1 蛋白分析與突變位點(diǎn)的選擇
首先對(duì) R.rhodochrous DSM43269的 5個(gè)不同類型KSHA的氨基酸進(jìn)行了比對(duì),結(jié)果如圖1所示.

圖1 KSHA5的氨基酸比對(duì)及結(jié)構(gòu)Fig. 1 Alignment of amino acid of KSHA5 and its structure
從圖 1(a)上可以看到,這 5種KSHA呈現(xiàn)高度相似,但loop區(qū)的氨基酸差別很大,α5螺旋、部分β折疊,以及二者之間的部分氨基酸也呈現(xiàn)很大差別,對(duì)照?qǐng)D1(b)和1(c)可以看到,α5螺旋和這部分β折疊的位置與 loop距離較近,這些位置的氨基酸都位于底物進(jìn)出活性中心的通道處.每種 KSHA的 loop的氨基酸不同,對(duì)應(yīng)的α5螺旋和這部分 β折疊的氨基酸就不同,推測(cè)它們之間可能存在協(xié)同變化的關(guān)系;而且,loop靠近底物入口處的氨基酸 T224–D228(圖1(b)和1(c)中被橢圓形標(biāo)中的loop區(qū)域),在有無底物AD時(shí)變動(dòng)非常明顯,那么它可能參與到底物的運(yùn)送過程進(jìn)而調(diào)控酶活,因此,本研究將對(duì)loop的這部分區(qū)域進(jìn)行定點(diǎn)突變,分別突變成性質(zhì)不同的氨基酸 T224V、N225L、Y226F、D227S與D228S,探究這些氨基酸的作用.
2.2 穿刺結(jié)果 所有病例中,28例經(jīng)穿刺活檢病理結(jié)合特殊染色直接確診(見圖2),但其中2例僅提示真菌感染,未明確特異病原體,其余26例明確了感染病原體,診斷敏感度、特異度分別為:93.3%、86.7%,診斷時(shí)限為送檢后1h至1周。確診的肺真菌病分別為:肺隱球菌病19例,肺曲霉菌病4例,肺接合菌病(傾向毛霉菌病)2例,肺馬爾尼菲藍(lán)狀菌病1例。2例未確診,1例提示肉芽腫性炎,經(jīng)氟康唑治療有效,臨床診斷肺隱球菌病,另1例外科術(shù)后病理證實(shí)肺毛霉菌。
提取 R.rhodochrous DSM43269基因組作為擴(kuò)增模板,擴(kuò)增羥化酶 kshAB基因序列.PCR 反應(yīng)體系(50,μL):基因組模板 1,μL,2×GC BufferⅠ25,μL,2.5,mmol/L dNTPs 8,μL,LA Taq DNA 聚合酶0.5,μL,10,μmol/L 上、下游特異性引物各 2,μL,ddH2O 補(bǔ)齊至 50,μL.反應(yīng)條件:94,℃ 5,min;94,℃30,s,60,℃ 45,s,72,℃ 2,min,30 個(gè)循環(huán);72,℃10,min.將PCR產(chǎn)物進(jìn)行回收純化,與pGEM-T Easy載體連接并送金唯智公司測(cè)序,測(cè)序正確后,經(jīng)酶切、連接獲得表達(dá)載體 pET28-kshA5與 pET28-kshB.膠回收純化、酶切與連接等參照試劑說明書.
2.2 定點(diǎn)突變結(jié)果
紫紅紅球菌(Rhodococcus rhodochrous)DSM43269購(gòu)自中國(guó)普通微生物菌種保藏管理中心;大腸桿菌(Escherichia coli)DH5α、BL21(DE3)均由本實(shí)驗(yàn)室保存.
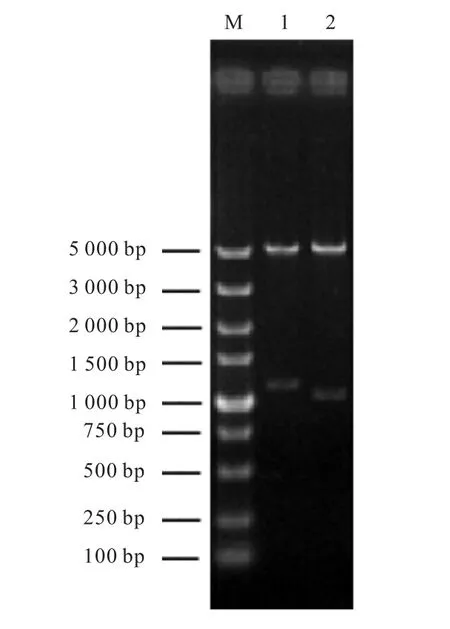
圖2 重組質(zhì)粒pET28-kshA5與pET28-kshB酶切電泳圖Fig. 2 Agarose gel electrophoresis of pET28-kshA5 and pET28-kshB treated with restriction endonuclease
突變位點(diǎn)和突變策略確定后,選用全質(zhì)粒 PCR的方法進(jìn)行基因突變,均擴(kuò)增出大約7,000,bp的條帶(圖3).經(jīng)測(cè)序證明都突變成了相應(yīng)的目的堿基.
兩個(gè)泳道在1,000,bp以上位置出現(xiàn)的條帶,分別與 kshA5和 kshB基因片段大小一致,證明重組質(zhì)粒pET28-kshA5與pET28-kshB構(gòu)建成功.
由于軌縫錯(cuò)臺(tái)值主要通過第一個(gè)測(cè)點(diǎn)(間隙信號(hào)4)與其他3個(gè)測(cè)點(diǎn)比較計(jì)算得到,因此在懸浮架采集的4路間隙信號(hào)中取兩端的間隙信號(hào)4進(jìn)行分析。由圖6的軌縫錯(cuò)臺(tái)變化仿真圖可知,間隙信號(hào)4基本反映了軌道錯(cuò)臺(tái)情況,通過錯(cuò)臺(tái)值能夠準(zhǔn)確表示高低突變。由于監(jiān)測(cè)探頭3、4之間有一定距離,因此錯(cuò)臺(tái)值出現(xiàn)一段平臺(tái)。
蚌埠市農(nóng)田水利建設(shè)成效明顯,為保障農(nóng)業(yè)和農(nóng)村經(jīng)濟(jì)發(fā)展發(fā)揮了重要基礎(chǔ)作用。但農(nóng)田水利建設(shè)基礎(chǔ)仍然薄弱,建設(shè)滯后、標(biāo)準(zhǔn)偏低、管理粗放的問題不同程度存在,相對(duì)滯后的農(nóng)田水利設(shè)施總體上依然是農(nóng)村經(jīng)濟(jì)發(fā)展的制約因素,影響農(nóng)業(yè)生產(chǎn)、農(nóng)民切身利益、水資源節(jié)約和國(guó)家糧食安全。
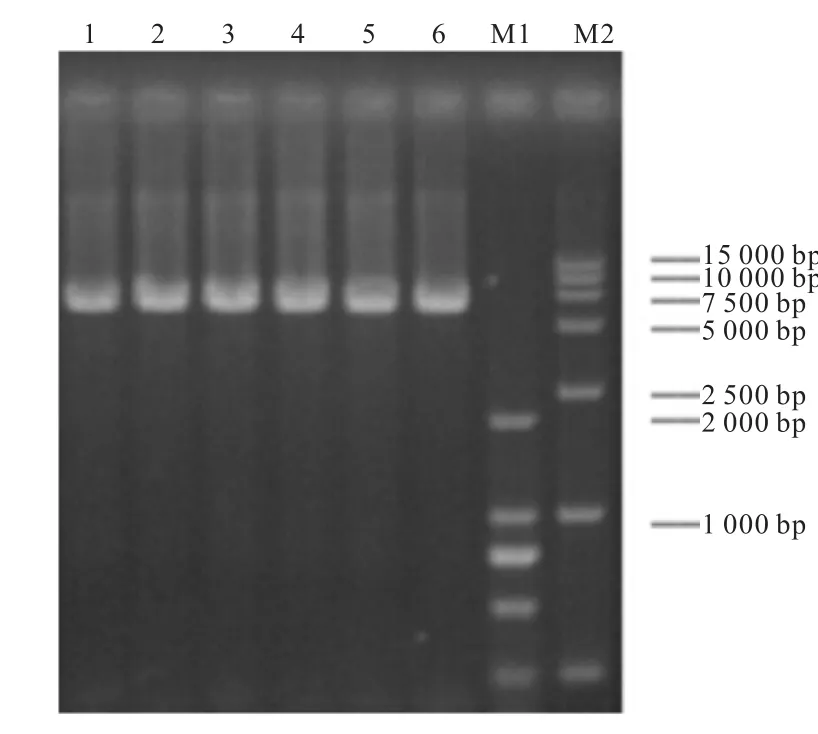
圖3 loop的定點(diǎn)突變Fig. 3 Site-directed mutagenesis of the loop
2.3 酶活分析
由于KSHAB酶是由加氧酶成分KSHA與還原酶成分KSHB同時(shí)存在才能發(fā)揮羥化酶KSHAB的活性,而且已有報(bào)道證明加氧酶組分KSHA以O(shè)2為最終電子受體,在空氣中純化 KSHA組分,會(huì)使KSHA組分失去活性,還原酶組分 KSHB,除了作為KSHAB酶催化體系的關(guān)鍵組分,同時(shí)還可以為KSHA的純化提供還原性的環(huán)境,避免在有氧情況下KSHA 酶活性的丟失[7–8].因此,本實(shí)驗(yàn)采用 A 組分與 B組分共純化的方式進(jìn)行蛋白純化,蛋白純化的結(jié)果如圖4所示.
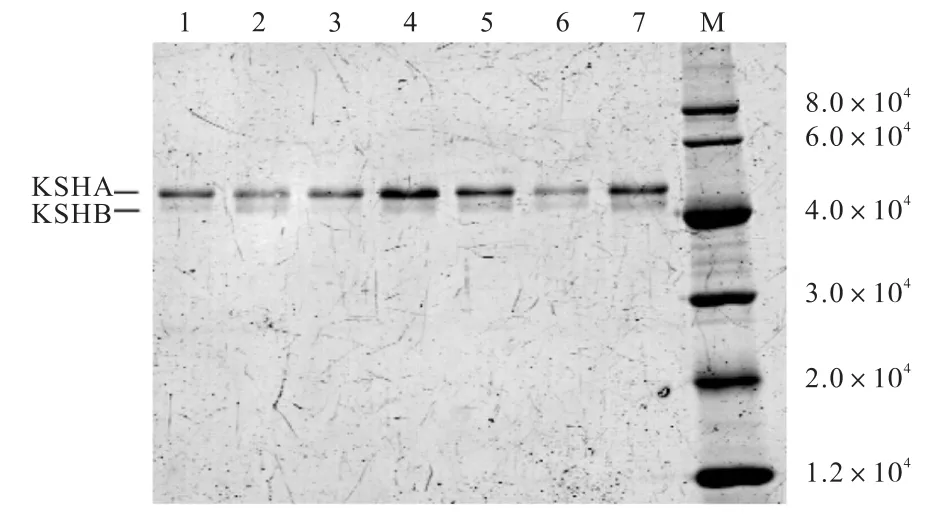
圖4 蛋白純化后的SDS-聚丙烯酰胺凝膠電泳圖譜Fig. 4 SDS-polyacrylamide gel electrophoresis of the purified proteins
由圖 4可知,所有蛋白均在相對(duì)分子質(zhì)量 4×104以上位置處顯示兩條帶,其大小與 KSHA、KSHB組分的大小一致,證明所有蛋白均純化成功.用軟件Quantity One分析所得各純化蛋白的A組分與B組分含量的比例,使用Trace Tracking方法以及高斯建模模擬各條帶的灰度值來確定A組分與B組分含量的比例.通過添加純KSHB蛋白,使各純化蛋白的A組分與 B組分含量的比例一致,測(cè)定酶活力的結(jié)果見表1.
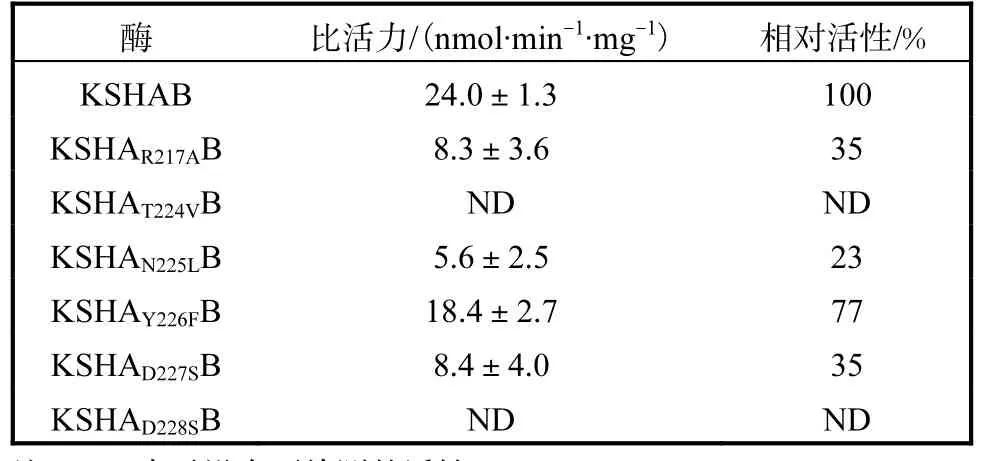
表1 突變體的活性分析Tab. 1 Assessment of the activity of different mutants
loop經(jīng)突變后的酶的活性較突變前的都有所降低,還有一些突變甚至給酶造成了完全失活的結(jié)果.將第217位的 R 變成 A后,活性降低為原來的35%,,堿性氨基酸變化成非極性氨基酸,喪失原來與第219位D之間的鹽橋作用,使loop更加松弛地靠在活性中心處,導(dǎo)致 loop上靠近底物入口處的氨基酸與α5螺旋、β折疊等附近區(qū)域的相互作用減弱,推送底物進(jìn)入活性中心的能力下降,可見,KSHA蛋白中該位置上的R高度保守是非常重要的.
T224側(cè)鏈帶有一個(gè)羥基,具有形成氫鍵的能力,從圖 5(a)與 5(b)上看,在結(jié)構(gòu)位置上,它更靠近α5螺旋.在底物進(jìn)入活性通道時(shí),T224有可能與α5螺旋上某個(gè)氨基酸作用,能較好地打開底物通道,順利讓底物通過,而其突變成 V后,極性發(fā)生較大改變.V是非極性氨基酸,容易埋在疏水性活性中心,不利于 loop打開通道,這可能是突變后監(jiān)測(cè)不到酶活的原因.同樣,第 225位 N變?yōu)?L后,活性也大大下降,Y226F也有明顯下降.突變前兩者的氨基酸具有形成氫鍵的能力,在蛋白質(zhì)結(jié)構(gòu)中是位于內(nèi)外部皆可的氨基酸,而突變后的非極性氨基酸更傾向于伸入疏水性活性中心,不利于打開loop這個(gè)“蓋子”,這也是 loop區(qū)該處位置在有無底物存在時(shí)發(fā)生很大變動(dòng)的原因.沒有底物時(shí),它們可以靠近活性中心,起到保護(hù)活性中心的作用;當(dāng)有底物存在時(shí),這些氨基酸又向外運(yùn)動(dòng),打開活性中心,在與周圍區(qū)域的相互作用力下,給底物營(yíng)造合適的環(huán)境,推送底物進(jìn)入活性中心.D227(圖5(c)與5(d))和D228從結(jié)構(gòu)上看,有無底物存在時(shí),雖然它們的位置變動(dòng)較大,但它們的側(cè)鏈更多地暴露在外面.D227和 D228是酸性氨基酸,可電離產(chǎn)生鹽橋,與溶液中離子相互作用,可能起到支撐固定loop結(jié)構(gòu)的作用,讓T224–Y226能與其周圍的氨基酸產(chǎn)生相互作用,調(diào)節(jié)底物通道;而突變成絲氨酸后,靈活性增強(qiáng),絲氨酸可轉(zhuǎn)動(dòng)到活性中心處,也可暴露在外,削弱了對(duì)loop的固定作用.
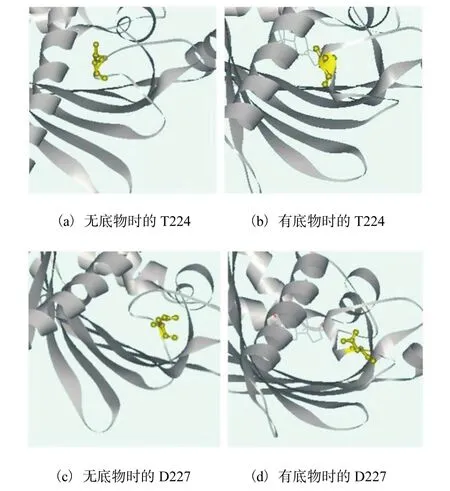
圖5 loop上靠近底物進(jìn)入酶活性中心入口處的氨基酸Fig. 5 Amino acids of the loop near the entrance of the substrate entering the enzyme active center
3 討 論
本文運(yùn)用定點(diǎn)突變技術(shù),確定了KSHA5的loop對(duì)酶活的重要性;loop上的氨基酸對(duì)穩(wěn)定 loop運(yùn)動(dòng)結(jié)構(gòu)和調(diào)節(jié)底物通道都有著不同的作用,尤其是靠近底物入口處的氨基酸,如 T224、N225和 Y226,它們與周圍α5螺旋和β折疊存在一定相互作用,共同調(diào)節(jié)底物通道與底物運(yùn)送能力;D227和D228起到支撐固定loop結(jié)構(gòu)的作用.磷酸丙糖異構(gòu)酶的loop上的A176就與其附近的β折疊上的Y208能產(chǎn)生相互作用,調(diào)節(jié) loop的開關(guān)運(yùn)動(dòng)[4].分枝桿菌(M.neoaurum)TCCC11028的3–甾酮–Δ1–脫氫酶靠近底物活性中心處的loop上的Ser138能形成氫鍵,在底物AD進(jìn)入活性中心后,能用氫鍵固定住 loop,保持活性中心的疏水性;而突變成 Leu后,氫鍵作用喪失,從而導(dǎo)致活性中心暴露,最終影響了酶對(duì)底物的活性[3].
在信息時(shí)代的大環(huán)境下,仍有許多施工單位的建設(shè)項(xiàng)目在管理項(xiàng)目檔案時(shí),采用傳統(tǒng)的管理辦法,在檔案資料的收集和整理過程中,單純依靠人工操作來進(jìn)行管理,不僅加大了管理人員的工作量,工作效率也受到很大的影響。
由于近年投入不足和管理缺位,面廣量大的田間溝渠被填埋、損毀嚴(yán)重,農(nóng)田灌溉“最后一公里”的問題非常突出,直接影響著河道治理工程的效益。要抓住全市10個(gè)縣(市、區(qū))列入中央財(cái)政小農(nóng)水重點(diǎn)縣的機(jī)遇,以縣為單位,以耕地灌區(qū)化為目標(biāo),將所有耕地以灌區(qū)為單元進(jìn)行規(guī)劃設(shè)計(jì)、投資建設(shè)、運(yùn)行管理,使之形成由可靠水源和引、輸、配水渠道及相應(yīng)排水溝道組成的灌溉系統(tǒng),形成田成方、林成網(wǎng)、渠相通、路相連、旱能澆、澇能排的高標(biāo)準(zhǔn)農(nóng)田建設(shè)格局。
loop在蛋白質(zhì)結(jié)構(gòu)中的運(yùn)動(dòng)較為靈活,底物進(jìn)出活性中心時(shí) loop旋轉(zhuǎn)擺動(dòng)的動(dòng)態(tài)過程,以及在這個(gè)運(yùn)動(dòng)過程中 loop上各氨基酸對(duì)底物的某種相互作用,還有底物進(jìn)入活性中心過程中l(wèi)oop與α5螺旋和部分β折疊的相互作用,在目前的晶體結(jié)構(gòu)信息中是無法獲知的,僅用無底物或者底物在活性中心時(shí)這兩個(gè)時(shí)刻解析的晶體結(jié)構(gòu),不能準(zhǔn)確判斷氨基酸的具體作用.而本研究運(yùn)用定點(diǎn)突變技術(shù)揭示了 loop上氨基酸的具體作用,這將使后面的研究工作更加具有針對(duì)性.那么,在下一步工作中,我們會(huì)把工作重點(diǎn)放到loop的T224、N225和Y226這3個(gè)氨基酸上,并結(jié)合其附近的α5螺旋與β折疊,進(jìn)行組合突變,研究三者對(duì)底物通道的調(diào)控,進(jìn)而能提高酶的羥化活性.本研究成果有助于理解 9α–羥化酶及其催化機(jī)理,也為采用蛋白質(zhì)工程改造 9α–羥化酶活性提供理論依據(jù).
[1] Angelova B,Mutafov S,Avramova T,et al. Effect of nitrogen source in cultivation medium on the 9αhydroxylation of pregnane steroids by resting Rhodococcus sp. cells[J]. Biotechnology & Biotechnology Equipment,2005,19(3):113–116.
[2] Penfield J S,Worrall L J,Strynadka N C,et al. Substrate specificities and conformational flexibility of 3-ketosteroid 9α-hydroxylases[J]. The Journal of Biological Chemistry,2014,289(37):25523–25536.
[3] Xie R,Shen Y,Qin N,et al. Genetic differences in ksdD influence on the ADD/AD ratio of Mycobacterium neoaurum[J]. Journal of Industrial Microbiology and Biotechnology,2015,42(4):507–513.
[4] Derreumaux P,Schlick T. The loop opening/closing motion of the enzyme triosephosphate isomerase[J]. Biophysical Journal,1998,74(1):72–81.
[5] Jennens M L,Lowe M E. A surface loop covering the active site of human pancreatic lipase influences interfacial activation and lipid binding[J]. The Journal of Biological Chemistry,1994,269(41):25470–25474.
[6] Dugi K A,Dichek H L,Santamarina-Fojo S. Human hepatic and lipoprotein lipase:The loop covering the catalytic site mediates lipase substrate specificity[J]. The Journal of Biological Chemistry,1995,270(43):25396–25401.
[7] 姚抗. 分枝桿菌甾醇代謝機(jī)制的解析以及其代謝工程改造應(yīng)用于制備重要甾藥中間體的研究[D]. 上海:華東理工大學(xué),2014.
[8] Petrusma M,Dijkhuizen L,van der Geize R. Rhodococcus rhodochrous DSM43269 3-ketosteroid 9α-hydroxylase,a two-component iron-sulfur-containing monooxygenase with subtle steroid substrate specificity[J].Applied and Environmental Microbiology,2009,75(16):5300–5307.
