轉染HBx改變Notch信號誘導肝癌HepG2細胞株多藥耐藥性研究
2018-02-05 09:30:21徐桐紅于冬艷
解放軍醫藥雜志 2018年1期
徐桐紅,于冬艷,任 玲
肝癌多藥耐藥(MDR)是影響化療效果及導致死亡的主要原因[1-2]。肝癌對化療藥物相對不敏感,對MDR發生機制不清楚。HBx具有多種調控功能,可激活腫瘤侵襲相關原癌基因和轉錄因子,是肝癌發病獨立危險因素[3-5]。Notch信號通路是高度保守的細胞間信號轉導通路。研究表明,Notch信號通路不僅對胚胎期腎臟發育起至關重要的作用,且參與多種腫瘤的發生和發展[6],其能通過血管內皮生長因子促進腫瘤血管發生,從而促進腫瘤細胞增殖[7]。多藥耐藥蛋白(MRP)過表達是肝癌產生MDR的重要機制[8-9]。本研究采用基因轉染將HBx基因轉入肝癌HepG2細胞中,觀察HBx對HepG2細胞中Notch信號通路和MRP表達的影響,并探討轉染HBx對肝癌細胞增殖和MDR的影響及其可能的作用機制,為肝癌的防治提供理論基礎。
1 材料與方法
1.1細胞株 人肝癌HepG2細胞株購自中科院上海生命科學院細胞庫,本所傳代保存,置于37℃、CO2體積分數為5%、含10%胎牛血清RPMI-1640培養基(Thermo Fisher Scientific,MA,USA)的恒溫培養箱內培養。
1.2試劑 DMEM高糖細胞培養液(美國Gibco公司,批號:12100046),DMEMF12(美國Gibco公司,批號:12400024),HBSS緩沖液(上海研卉生物技術有限公司,批號:BS0101),Trizol裂解液(美國賽默飛世爾科技,批號:15596026),Taqman MicroRNA反轉錄試劑盒(美國賽默飛世爾科技,批號:4366596),Lipofectamine 2000轉染試劑盒(美國賽默飛世爾科技,批號:722255),Annexin-V-FITC/PI細胞凋亡檢測試劑盒(上海前塵生物科技有限公司,批號:40302ES20),B27補充劑(美國GIBCO,批號:17504-044),CCK-8試劑盒(上海鈺博,批號:YB1198),小牛血清(北京索萊寶公司,批號:01-045),胰蛋白酶(北京瑞達恒輝科技發展有限公司,批號:HB-0103-01),RPMI-1640培養基(美國GIBCO公司,批號:11875-093)。
1.3實驗方法
1.3.1重組X質粒轉染肝癌HepG2細胞:在10%胎牛血清的DMEN中培養HepG2細胞,胰酶消化后,接種于直徑25 cm培養瓶中,此時計數細胞數為1×106個/L,細胞在6 h后貼壁,之后進行細胞轉染。在37℃預熱的無血清培養基加入5 μg DNA和15 μl Trans Fast Reagent,隨機高速渦旋混勻,在室溫下靜置15 min。培養基被小心移出,之后慢慢向培養瓶中加入混勻的DNA/脂質體的混合物,在37℃孵育24 h后,輕輕加入4 ml含血清的培養基(完全培養基),不必移去DNA/脂質體混合物,37℃繼續孵育48 h;以HepG2細胞中轉染重組x質粒命名為轉染HBx細胞組(HepG2-HBx組)、未轉染重組x質粒命名為轉染空載體細胞組(HepG2-con組),未進行任何處理的為空白細胞組(HepG2組)。
1.3.2實時定量聚合酶鏈反應(PCR)檢測HBx mRNA的表達:TRIzol試劑提取并收集處于對數生長期的3組肝癌HepG2細胞的HBx mRNA,RNA完整性利用1%的瓊脂糖變性凝膠電泳進行檢測,純度和濃度利用紫外可見分光光度計檢測;在70℃(10 min)、冰育(2 min)、42℃(60 min)、70℃(10 min)反應條件下,利用逆轉錄試劑盒逆轉錄得到cDNA;在95℃(5 s)、60℃(20 s)、72℃(5 s)進行qRT-PCR反應,40個循環,重復試驗3次。
1.3.3蛋白免疫印跡法(Western-blot)檢測肝癌HepG2細胞中HBx、Notch-1和MRP表達:轉染組和未轉染組進行細胞總蛋白提取,通過Bradford方法調整相同的蛋白濃度后,行電泳獲得根據分子量分離的蛋白條帶,根據pierce公司Western-blot試劑盒的指導說明進行,蛋白NC膜電轉,進行蛋白封閉和一抗結合,一抗結合后洗脫,再進行二抗孵育標記兔抗大鼠的多克隆抗體。ECL試劑盒顯色后,暗室發光拍照,以β-actin為內參進行系統軟件分析。
1.3.4CCK-8法檢測細胞增殖活性:取3組細胞以1×106個/L接種于96孔板中,每孔100 μl,培養48 h后每孔加入10 μl CCK-8試劑,4 h后在450 nm波長下檢測各孔吸光度(OD)值,重復3次取均值。以OD值為縱坐標,時間為橫坐標,繪制生長曲線。
1.3.5流式細胞儀檢測細胞周期:收集轉染48 h后的3組細胞,用冷PBS洗滌細胞后加入體積分數70%冷乙醇固定過夜。采用Cell Quet軟件分析細胞周期分布情況。實驗重復3次。
1.3.6細胞耐藥性檢測:將3組處于對數生長期的HepG2細胞制成濃度為1×106個/L的細胞懸液,每孔100 μl,接種于96孔板,每孔設3個復孔。孵育24 h后,換用濃度為0.5、1、2.5、5、10、25、50、100 μmol/L絲裂霉素、阿霉素培養液,0.5、1、2.5、5、10、25、50、100、250、500 μmol/L 5-氟尿嘧啶、順鉑、奧沙利鉑培養液。孵育48 h后,去除含藥液,每孔加100 μl不含藥培養基和10 μl CCK-8,設空白孔。孵育2 h后,酶標儀測光密度值,設a為加藥組的OD值,b為不加藥細胞組的OD值,c為無細胞的空白組的OD值,藥物對細胞的抑制率(%)=[1-(a-c)/(b-c)]×100%。計算5種藥物抑制率為50%的濃度(IC50)和耐藥指數(RI),RI=耐藥細胞IC50/親本細胞IC50。
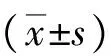
2 結果
2.1HBx mRNA的表達 將PCR擴增產物進行凝膠電泳,可見HepG2-HBx組有目的片段出現,而其他2組未見HBx mRNA的表達,見圖1。
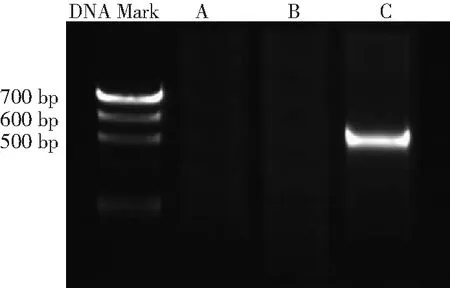
圖13組肝癌細胞HepG2聚合酶鏈反應擴增產物凝膠電泳結果
A.HepG2組;B.HepG2-con組;C.HepG2-HBx組
2.2HBx、Notch-1和MRP蛋白表達 HepG2-HBx組有HBx蛋白表達,HepG2組和HepG2-con組未見表達。Notch-1和MRP蛋白在HepG2-HBx組條帶最大,HepG2-con組次之,HepG2組最小。見圖2。
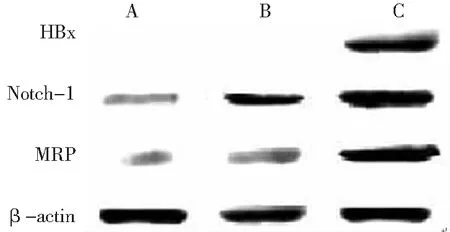
圖23組肝癌HepG2細胞中HBx、Notch-1和多藥耐藥蛋白的表達情況

A.HepG2組;B.HepG2-con組;C.HepG2-HBx組
2.3轉染HBx后各組細胞的生長曲線 與HepG2組和HepG2-con組比較,HepG2-HBx組細胞增殖顯著加快(P<0.05)。見表1。
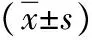
表1 3組肝癌HepG2細胞培養不同時間吸光度值比較
注:與HepG2組比較,aP<0.05,bP<0.01;與HepG2-con組比較,cP<0.05
2.4轉染HBx后細胞周期變化 與HepG2組和HepG2-con組比較,HepG2-HBx組G0/G1期細胞顯著減少,S期細胞增加(P<0.05,P<0.01),G2/M期細胞變化不大(P>0.05)。見表2。
2.5耐藥性 與HepG2組和HepG2-con組比較,HepG2-HBx組中肝癌細胞HeG2對5種藥物耐藥指數增高(P<0.05,P<0.01)。見表3。
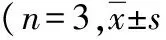
表2 3組肝癌HepG2細胞周期變化情況,%)
注:與HepG2組比較,aP<0.05,bP<0.01;與HepG2-con組比較,cP<0.05,dP<0.01
表3 3組肝癌HepG2細胞對藥物的敏感性比較
注:IC50為藥物抑制率為50%的濃度,RI為耐藥指數;與HepG2組比較,aP<0.05,bP<0.01;與HepG2-con組比較,cP<0.05,dP<0.01
3 討論
HBV慢性感染是世界范圍內原發性肝細胞癌(HCC)的主要發病原因之一[10-11],HBx基因致HCC發生的機制是目前研究的熱點。HBx基因具有強大的生物學功能,包括反式激活病毒基因組和宿主細胞基因的轉錄、增強轉錄因子DNA結合特性、抑制p53蛋白活性、參與細胞信號轉導途徑和細胞凋亡的調節等。這些功能可能與HCC的發生具有密切的關系[12]。一些體內及體外研究也發現,HBx可誘導細胞的惡性轉化,甚至參與癌變發生[13]。
在腫瘤的發生、發展過程中,Notch信號出現紊亂時,能夠直接引起腫瘤的發生和發展,且可通過多條信號通路的相互作用,間接誘導腫瘤的形成[14]。本研究qPCR檢測結果顯示,轉染HBx的HepG2細胞內HBx mRNA和蛋白表達明顯,提示轉染構建過表達HBx的肝癌細胞成功。本研究還發現,轉染HBx人肝癌HepG2細胞Notch-1蛋白表達高于HepG2組和HepG2-con組,提示HepG2細胞中HBx過表達誘導了細胞內Notch-1表達水平的增加,造成細胞內Notch信號通路的紊亂。細胞增殖和周期檢測結果顯示,轉染HBx后細胞增殖能力顯著提高,S期細胞增加,提示Notch-1的活化誘導了HepG2細胞增殖的發生。
MDR是導致腫瘤治療失敗和復發的主要原因之一[15-16]。有報道指出,檢測患者體內MRP的表達可以作為監測惡性腫瘤細胞原發性耐藥的主要指標[17]。腫瘤細胞產生MRP與藥物外排泵ABC轉運蛋白的過度表達與細胞凋亡水平異常改變、細胞DNA修復活性增強、胞內酶異常改變和器官微環境改變有密切關系[18]。此外,Notch-1信號與腫瘤細胞MDR的形成有關,過表達Notch-1能增加與MDR相關的ABBCC1基因的表達,誘導MDR的發生[19]。本研究發現,轉染HBx的HepG2細胞耐藥指數明顯增高,提示轉染HBx HepG2細胞,可能通過誘導Notch-1和MRP的過表達,最終導致MDR的發生。此外,在本研究基礎上檢測Notch通路的活化情況及由此引起凋亡抑制蛋白livin表達,同時研究livin的表達后MDR相關蛋白表達的變化,并觀察HBx、Notch通路活化、livin蛋白及MDR蛋白表達之間的相關性是下一步的研究重點,為乙型肝炎相關性HCC臨床治療提供新靶點。
[1] 韓冬,肖亮,占小波,等.多藥聯合方案建立人肝癌多藥耐藥細胞株[J].實用醫學雜志,2015,31(22):3691-3693.
[2] 徐鋮,羅華榮,甘梅富.非小細胞肺癌中EGFR及多藥耐藥蛋白與化療療效的關系[J].中國生化藥物雜志,2017,37(5):93-95,97.
[3] Hoare J, Henkler F, Dowling J J,etal. Subcellular localisation of the x protein in hbv infected hepatocytes[J].J Med Virol, 2001,64(4):419-426.
[4] Su Q, Schroder C H, Hofmann W J,etal. Expression of hepatitis B virus X protein in HBV-infected human livers and hepatocellularcarcinomas[J].Hepatology, 1998,27(4):1109-1120.
[5] 李偉,劉傳苗,趙守松.原發性肝細胞癌患者肝組織中乙型肝炎病毒x基因的變異[J].實用醫學雜志,2009,25(2):180-182.
[6] Dormoy V, Jacqmin D, Lang H,etal. From development to cancer: lessons from the kidney to uncover new therapeutic targets[J].Anficancer Res, 2012,32(9):3609-3617.
[7] MaⅡIard I, Fang T, Pear W S. ReguIation of Iymphoid deveIopment, differentiation, and function bytheNotch pathway[J].Annu ReUImmunol, 2005,23:945-974.
[8] 孫海兵,張開光,李琴,等.TRAIL逆轉人胃腺癌SGC7901/ADR細胞多藥耐藥的機制探討[J].山東醫藥,2017,57(13):9-12.
[9] 王海娟,張美芝,劉小菊,等.雷公藤內酯醇逆轉腫瘤多藥耐藥研究進展[J].中國實驗方劑學雜志,2016,22(10):229-234.
[10] 余咸輝,鄢業鴻,董春華,等.HBx在乙肝相關型肝細胞癌中的作用機制[J].廣東醫學,2016,37(23):3614-3618.
[11] 王文耀,張鴻飛,唐淼,等.miR-21 在肝癌中的表達及其與 PTEN 的關系[J].中國普通外科雜志,2017,26(3):395-400.
[12] Huang J, Deng Q, Wang Q,etal. Exome sequencing of hepatitis B virus-associated hepatocellular carcinoma[J].Nat Genet, 2012,44(10):1117-1123.
[13] Chen L, Hu L, Li L,etal. Dysregulation of β-catenin by hepatitis B virus X protein in HBV-infected human hepatocellular carcinomas[J].Front Med China, 2010,4(4):399-411.
[14] Bridges E, Oon C E, Harris A. Notch regulation of tumor angiogenesis[J].Future Oncol, 2011,7(4):569-588.
[15] 嚴林海,韋尉元,肖強,等.尾型同源盒基因2過表達對人胃癌耐順鉑細胞SGC7901/DDP多藥耐藥性的影響[J].中華消化外科雜志,2014,13(2):145-148.
[16] 羅文,楊杰,廉超,等.尾型同源盒基因2沉默對人胃癌耐順鉑細胞SGC7901/DDP多藥耐藥性的逆轉[J].中華消化外科雜志,2014,13(1):55-61.
[17] 宋祖麗,李振光,王凈凈,等.穴位埋藥線對難治性癲癇大鼠癲癇樣波的發放及大腦海馬和顳葉皮質多藥耐藥相關蛋白MRP-1、P-gp表達的影響[J].湖南中醫藥大學學報,2016,36(10):76-81.
[18] Yergeri M, Kapse-Mistry S, Srivastava R,etal. Nanodmg delivery in reversing muttidrug resistance in cancer cells[J].Front Pharmacol, 2014,5(159):1-6.
[19] 李艷霞.Notch-1通路在人胰腺癌細胞上皮-間質轉化和腫瘤干性中的作用機制研究[D].武漢:武漢大學,2016.
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
海峽科技與產業(2016年3期)2016-05-17 04:32:12
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年17期)2015-03-01 02:03:58
中國當代醫藥(2015年16期)2015-03-01 02:03:11
