黃芩甙對新城疫病毒感染雞胚成纖維細胞Toll樣受體2、3、4、7 mRNA表達的影響
2018-01-05 02:45:32許銳光胡力文翁嘉華鄧俊良胡延春
浙江農業學報 2017年12期
許銳光,賈 艷,胡力文,翁嘉華,鄧俊良,胡延春,*
(1.四川農業大學 動物醫學院/動物疫病與人類健康四川省重點實驗室,四川 成都 611130; 2.嘉興職業技術學院 農業與環境分院,浙江 嘉興 314036)
黃芩甙對新城疫病毒感染雞胚成纖維細胞Toll樣受體2、3、4、7 mRNA表達的影響
許銳光1,賈 艷2,胡力文1,翁嘉華1,鄧俊良1,胡延春1,*
(1.四川農業大學 動物醫學院/動物疫病與人類健康四川省重點實驗室,四川 成都 611130; 2.嘉興職業技術學院 農業與環境分院,浙江 嘉興 314036)
旨在探討黃芩甙對新城疫病毒感染雞胚成纖維細胞Toll樣受體2、3、4、7 mRNA表達的影響。新城疫病毒(NDV)感染雞胚成纖維細胞,使用MTT法檢測黃芩甙對雞胚成纖維細胞的最大安全濃度,使用熒光定量PCR方法檢測不同濃度黃芩甙對染毒細胞Toll樣受體2、3、4、7 mRNA在不同時間表達的影響。結果表明,黃芩甙組和病毒對照組細胞TLR2、TLR4和TLR7 mRNA的表達水平起初受到抑制,TLR2和TLR4 mRNA在24 h內呈上升趨勢,TLR7 mRNA在16 h內呈上升趨勢,TLR3 mRNA的表達水平呈波動性變化。黃芩甙在24 h時能夠促進NDV感染細胞TLR2和TLR4 mRNA的表達,且呈劑量依賴性,從而發揮免疫調節作用,具有抗NDV的功能。
黃芩甙;新城疫病毒;Toll樣受體;實時熒光定量PCR
新城疫病毒(Newcastlediseasevirus, NDV)也被稱為亞洲雞瘟病毒、偽雞瘟病毒或禽肺腦炎病毒,屬于副黏病毒科,禽腮腺炎病毒屬(Avulavirus),禽副黏病毒I型(APMV-1)[1]。最急性NDV感染雞群,死亡率高達90%,其每年對全球養禽業都造成巨大的損失[2]。接種疫苗是目前本病主要的防治措施[3],因大量抗病毒西藥被禁止應用于畜禽臨床中,故選擇一種天然化合物作為替代十分有應用前景[4]。許多中藥本身具有調節免疫功能、抑制病毒增殖和感染[5]等作用,以及副作用小、毒性低等優良特性。但目前中藥在動物醫學領域的應用并不廣泛。中藥抗病毒不只單純著眼于直接的抗病毒作用,更重視“病毒-機體-藥物”的關系,不僅直接殺滅病毒,阻止病毒的復制、轉染,更重要的是能夠增強機體的免疫力,激發調動一切的免疫防御系統發揮抗病毒作用[5]。黃芩甙(C21H18O11,圖1)是提取自中藥黃芩的干燥根中的一種主要活性成分,它具有抗炎[6]、抗病毒[7]、抗菌[8]和調節免疫[9]等生物活性。Jia等[10]的研究表明,黃芩甙具有體外直接抗NDV作用。
通過模式識別受體來識別病原微生物的特異性成分是機體發揮免疫應答抵御病原微生物侵入的前提和基礎。Toll樣受體(TLRs)是一類重要模式識別受體,在脊椎動物中具有顯著的保守性,能夠識別細菌、病毒等微生物的各種組分[11]。當TLRs被配體激活后,信號通過相關分子的傳導,激發一系列的免疫應答反應,產生炎癥細胞因子,抵御包括病毒在內的病原微生物的侵入[12]。目前,在禽體內已發現的TLRs為TLR1、TLR2、TLR3、TLR4、TLR5、TLR7、TLR15和TLR21[13]。其中,TLR2、TLR3、TLR4、TLR7在NDV感染機體時發揮作用。本文通過研究黃芩甙對新城疫病毒感染雞胚成纖維細胞Toll樣受體2、3、4、7 mRNA表達的影響,旨在揭示黃芩甙能夠通過調節機體的免疫力,激發調動免疫防御系統發揮抗NDV作用的機理,以期為臨床抗NDV新藥的研發提供理論和試驗依據。

圖1 黃芩甙的化學結構式Fig.1 Chemical structure of Baicalin
1 材料與方法
1.1 試驗材料
藥物:黃芩甙,購自上海金穗生物科技有限公司,純度≥98%。
病毒:新城疫病毒(La Sota株)由四川農業大學預防獸醫系黃勇教授提供。
試驗試劑:DMEM培養基購自Hyclone;噻唑藍(MTT)和二甲基亞砜(DMSO)購自Sigma;胎牛血清購自杭州四季青生物工程材料有限公司;TRIpure總RNA快速提取試劑盒購自成都百菲特科技有限公司;TUREscript 1st Strand cDNA Synthesis Kit和BestarSybrGreen qPCR mastermix購自北京艾德萊生物科技有限公司。
儀器設備:37XC009312型倒置顯微鏡,上海江南光學儀器廠;Thermo311型二氧化碳培養箱,Thermo Fisher Scientific;YXQ-LS-50S11型高壓滅菌鍋,上海博迅實業有限公司醫療責任設備廠;1645050型電泳儀,iMark型酶標儀,CFX96熒光定量PCR儀和GelDoc XR+型凝膠成像系統,BIO-RAD公司。
1.2 方法
1.2.1 細胞和病毒
按文獻[4]方法制備雞胚成纖維細胞(CEF)。
將NDV La Sota株(病毒含量為100 EID50·mL-1)接種于9日齡雞胚尿囊腔,每胚接種0.2 mL,24 h內死亡雞胚棄之。收集24 h后死亡雞胚尿囊液,0.22 μm濾膜過濾除菌、加雙抗(青霉素G和硫酸鏈霉素各100單位·mL-1)、分裝,-80 ℃保存。
1.2.2 NDV病毒對CEF細胞TCID50的測定
將病毒液用DMEM培養基對數稀釋成10-1,10-2,…,10-8。吸取每一稀釋度的病毒液100 μL加入吸去培養基的單層CEF 96孔板的相應孔中,每個稀釋度重復4孔,在37 ℃、5% CO2培養箱中吸附1.5 h。吸棄病毒液,每孔加入200 μL維持液,培養72 h,在倒置顯微鏡下觀察致細胞病變效應(CPE),并記錄結果。用Reed-Muench方法計算病毒的TCID50。
1.2.3 黃芩甙對CEF細胞安全濃度的測定
用含2%胎牛血清的DMEM培養基,即維持液稀釋Baicalin 至2 mg·mL-1,過濾除菌。倍比稀釋8個梯度加至吸去培養基的單層CEF 96孔板的相應孔中,100 μL·孔-1,每個稀釋度重復4孔,同時設細胞對照孔和空白對照孔, 37 ℃、5%CO2培養。加藥后每12 h 觀察1次細胞單層完好程度, 培養72 h 后, 按MTT法[14]用酶標儀測定570 nm 波長處D570值。選擇細胞D570值不顯著小于細胞對照組的單體最大質量濃度作為該單體最大安全濃度[15]。
1.2.4 熒光定量PCR引物合成
根據文獻[16]提供的序列合成特異性引物,如表1。
1.2.5 黃芩甙對NDV感染的CEF細胞TLR2、TLR3、TLR4、TLR7 mRNA的調控試驗
(1)樣品準備
取處于對數期CEF細胞,用細胞生長液調整細胞密度為 5×105個·孔-1接種到 6 孔細胞培養板。待CEF細胞長至單層時,棄舊培養液,加入
100TCID50的NDV,37 ℃吸附 1.5 h,棄去病毒液,黃芩甙由最大安全濃度倍比稀釋5個濃度梯度加入6孔板,并設置細胞對照和病毒對照,每組重復4次。分別于4、8、16、24、48、72 h后終止試驗,冰上吸出培養液,PBS 洗3次,每孔中加入1 mL Trizol,用細胞刮刮凈細胞,儲存于-80 ℃冰箱,用于 RNA 提取。
(2)細胞總 RNA 的提取
根據TRIpure總RNA提取試劑盒說明書進行操作,提取細胞總RNA。運用變性瓊脂糖凝膠電泳檢測提取的總RNA的完整性,130 V穩壓,1×TBE Buffer 1%變性瓊脂糖凝膠電泳20~30 min。
(3)總 RNA 的反轉錄反應
根據TUREscript 1st Strand cDNA Synthesis Kit說明書進行反轉錄反應合成cDNA。所得產物-20 ℃冰箱長期保存。
(4)實時熒光定量 PCR 反應
取合成的cDNA作為DNA模板,每組設3個平行重復孔,按照BestarSybrGreen qPCR mastermix試劑盒說明書進行PCR擴增。反應程序為:95 ℃ 2 min;94 ℃ 15 s,60 ℃ 30 s,72 ℃ 30 s,共40個循環。
1.2.6 數據分析
由 SPSS 20.0軟件進行數據統計分析和差異顯著性分析。黃芩甙對CEF細胞安全濃度的測定數據用Mean±SE表示。在進行相對熒光定量時以18sRNA內參基因作為標準。基因相對表達量的計算用2-ΔΔCt法。
2 結果與分析
2.1 NDV的TCID50測定結果
表1引物序列及擴增片段長度
Table1The primers sequences and amplification fragments sizes

目的基因TargetmRNA上游引物Forwardprimers(5′-3′)下游引物Reverseprimers(5′-3′)產物大小Productsize/bpTLR2GATTGTGGACAACATCATTGACTCAGAGCTGCTTTCAAGTTTTCCC294TLR3TCAGTACATTTGTAACACCCCGCCGGCGTCATAATCAAACACTCC256TLR4AGTCTGAAATTGCTGAGCTCAAATGCGACGTTAAGCCATGGAAG190TLR7TTCTGGCCACAGATGTGACCCCTTCAACTTGGCAGTGCAG21918SrRNATCAGATACCGTCGTAGTTCCTTCCGTCAATTCCTTTAAGTT154
根據CPE情況計算NDV的TCID50,結果得出TCID50=10-5.5·mL-1,本試驗使用的病毒濃度為100TCID50,故本試驗使用的病毒濃度為10-3.5·mL-1。
2.2 黃芩甙對CEF細胞的安全濃度
根據MTT試驗結果,選擇合適的植物單體作用于CEF細胞的藥物濃度。黃芩甙對CEF細胞的安全濃度測定結果圖 2。黃芩甙對CEF細胞的最大安全濃度為0.25 mg·mL-1。
2.3 總RNA的完整性
結果如圖3所示,總RNA的18S RNA和28S RNA條帶比較清晰、明亮、規則,并且18S RNA和28S RNA之間無拖帶,說明所提總RNA完整性較高,可用來合成cDNA。

A,黃芩甙組; B,病毒對照組; C,細胞對照組A, Baicalin group; B, NDV control group; C, cell control group圖3 總RNA變性瓊脂糖凝膠電泳Fig.3 Denatured agarose gel electrophoresis test of total RNA extracted from CEFs
2.4 黃芩甙對染毒細胞TLR2、TLR3、TLR4、TLR7 mRNA表達作用的影響
2.4.1 黃芩甙對染毒細胞TLR2 mRNA表達作用的影響
黃芩甙組和病毒對照組細胞TLR2 mRNA的表達水平起初受到抑制,極顯著低于細胞對照組(P<0.01),在24 h內呈上升趨勢,于24 h時極顯著高于細胞對照組(P<0.01),最大相對表達量分別提高了149%(黃芩甙組)和67%(病毒對照組),并在24 h后呈下降趨勢。在24 h時黃芩甙組的TLR2 mRNA的表達水平與濃度成正相關,并在濃度為1×2-2~1×2-4mg·mL-1時極顯著高于病毒對照組(P<0.01),在濃度為1×2-5mg·mL-1時顯著高于病毒對照組(P<0.05),在濃度為1×2-5mg·mL-1時與病毒對照組無顯著差異。故黃芩甙能夠促進NDV感染細胞TLR2 mRNA的表達且促進能力與濃度呈正相關,從而能夠通過免疫調節發揮抗病毒作用(圖4-A)。
2.4.2 黃芩甙對染毒細胞TLR3 mRNA表達作用的影響
黃芩甙組和病毒對照組細胞TLR3 mRNA的表達水平呈波動性變化,無明顯規律。而不同濃度的黃芩甙組的TLR3 mRNA表達水平與病毒對照組無顯著差異(P>0.05)。故黃芩甙對NDV感染細胞TLR3 mRNA的表達無顯著影響(圖4-B)。
2.4.3 黃芩甙對染毒細胞TLR4 mRNA表達作用的影響
黃芩甙組和病毒對照組細胞TLR4 mRNA的表達水平起初受到抑制,極顯著低于細胞對照組(P<0.01),在24 h內呈上升趨勢,于24 h時極顯著高于細胞對照組(P<0.01),最大相對表達量分別提高了93%(黃芩甙組)和52%(病毒對照組)并在24 h后呈下降趨勢。在24 h時黃芩甙組的TLR4 mRNA的表達水平與濃度成正相關,并在濃度為1×2-2~1×2-4mg·mL-1時極顯著高于病毒對照組(P<0.01),在濃度為1×2-5mg·mL-1時顯著高于病毒對照組(P<0.05),在濃度為1×2-6mg·mL-1時與病毒對照組無顯著差異。故黃芩甙能夠促進NDV感染細胞TLR4 mRNA的表達且促進能力與濃度呈正相關,從而能夠通過免疫調節發揮抗病毒作用(圖4-C)。
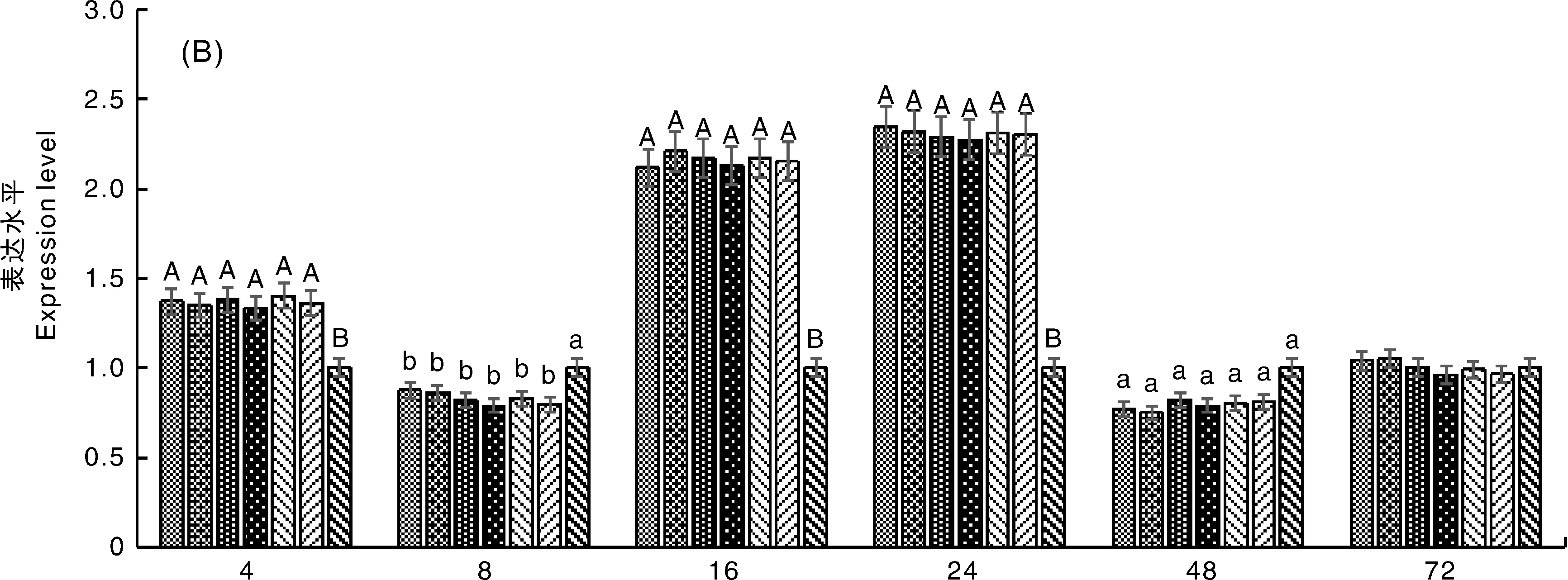


在同一時間,無相同大寫字母表示差異極顯著(P<0.01),無相同小寫字母表示差異顯著(P<0.05)At the same time, the difference between data with different capital letter is very significant(P<0.01), and the difference between data with different small letters is significant (P<0.05)圖4 不同處理組CEF細胞內TLR2(A)、TLR3(B)、TLR4(C)、TLR7(D) mRNA的表達水平Fig.4 Expression levels of TLR2(A)、TLR3(B)、TLR4(C)、TLR7(D) in NDV-infected CEFs of different treatment groups
2.4.4 黃芩甙對染毒細胞TLR7 mRNA表達作用的影響
黃芩甙組和病毒對照組細胞TLR7 mRNA的表達水平起初受到抑制,極顯著低于細胞對照組(P<0.01),在16 h內呈上升趨勢,于16 h時極顯著高于細胞對照組(P<0.01),相對表達量提高了約120%,并在16 h后呈下降趨勢。不同濃度黃芩甙組的TLR7 mRNA表達水平與病毒對照組無顯著差異。故黃芩甙對NDV感染細胞TLR7 mRNA的表達無顯著影響(圖4-D)。
3 討論
了解NDV感染和免疫系統的相互作用對于理解新城疫的發病機制是十分重要的。TLRs是細胞對抗病原入侵的首道防線,能夠作為模式識別受體識別病原。到目前為止,已經有13種TLRs被發現,它們可以調節產生各種炎性細胞因子、化學增活素和干擾素[17]。其中,TLR2、TLR3、TLR4和TLR7在因病毒感染產生的天然免疫反應中發揮重要的作用。
TLR2可以激活MyD88信號通路,激活NF-κB和AP-1轉錄因子和干擾素調節因子(IRFs),從而協調調節宿主對微生物感染的防御[18]。越來越多的證據表明,TLR/MyD88信號轉導通路在機體對微生物病原體的先天免疫和適應性免疫應答的激活發揮著核心作用[19-20]。本試驗結果表明,在NDV感染早期,TLR2 mRNA的表達被NDV抑制,但隨著NDV的增殖,TLR2 mRNA的表達量增加并于24 h達峰值。故NDV能夠激活TLR2的復制,從而激活MyD88信號轉導通路協同調節宿主防御病毒。本研究中,黃芩甙能增加TLR2 mRNA在NDV感染細胞的表達并增強上述免疫作用。
TLR3屬于I型跨膜蛋白,是存在于細胞內的富含亮氨酸的重復序列(LRR),并且是一個Toll-IL-1受體(TIR)的同源信號域[21]。TLR3受體可以識別體外轉錄的dsRNA[22]。Cheng等[23]的研究結果表明,TLR3能夠積極參與NDV感染時的前期炎癥反應,并且導致抗病毒細胞因子分泌。本研究結果表明,NDV感染可引起TLR3 mRNA表達的變化,但它沒有規律,并且黃芩甙對TLR3 mRNA在NDV感染的細胞中的表達無明顯影響。
和TLR2一樣,TLR4也能夠激活MyD88信號轉導通路[24]。一些TRIF相關的接頭分子(TRAM)只能被TLR4所利用,MyD88適配器是一個連接適配器,聯通TLR4和TRAM[25]。受體復合物的形成激活信號轉導通路,產生 NF-κB、IRF3、IRF5 和IRF7轉錄因子,導致特異性免疫途徑基因表達從而調節機體的免疫機能[17]。本試驗結果表明,在NDV感染早期,TLR4 mRNA的表達被NDV抑制。但隨著NDV的增殖,TLR4 mRNA的表達量增加并于24 h達峰值。故NDV能夠激活TLR4的復制。TLR4可以激活MyD88信號轉導通路協同調節宿主防御病毒。本研究中,黃芩甙能增加TLR4 mRNA在NDV感染細胞的表達,并增強上述免疫作用。
TLR7能夠識別病毒的ssRNA,并且是機體對ssRNA病毒外周免疫反應的重要媒介[26]。它主要參與促炎性細胞因子的產生,但具體機制尚不清楚[27]。本研究結果表明,在NDV感染早期,TLR7 mRNA的表達被NDV抑制,但隨著NDV的增殖,TLR7 mRNA的表達量增加并于16 h達峰值。故NDV能夠激活TLR7的復制從而調節宿主防御病毒。本研究中,黃芩甙對TLR7基因在病毒感染細胞的表達無明顯影響。
眾所周知,TLR2和TLR4均可以參與機體對病毒糖蛋白誘導產生促炎癥反應[17]。在機體中,TLR2是TLR4的輔助受體,TLR2和TLR4基因的表達呈正相關關系[28]。在本研究中,TLR2和TLR4的mRNA具有相似的表達變化趨勢,在NDV感染早期,其mRNA的表達均被NDV抑制。但隨著NDV的增殖其mRNA表達量均增加并于24 h達峰值。這是與前人的研究結果一致。
本研究結果表明,NDV感染影響了TLR2、TLR3、TLR4、TLR7 mRNA的表達,黃芩甙可以極顯著地促進NDV感染細胞TLR2和TLR4 mRNA的表達,發揮免疫調節作用,具有抗NDV的功能。
[1] 蔡寶祥. 家畜傳染病學[M]. 北京:中國農業出版社, 2001.
[2] PALYA V, KISS I, TATR-KIS T, et al. Advancement in vaccination against Newcastle disease: recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions[J].AvianDiseases, 2017, 56(2):282-287.
[3] 羅玉均,陳建紅,張濟培. 雞新城疫的診斷與防制[J]. 養禽與禽病防治, 2004 (12):7.
LUO Y J, CHEN J H, ZHANG J P. Diagnosis and control of Newcastle disease[J].PoultryHusbandryandDiseaseControl, 2004 (12):7.(in Chinese)
[4] ZHANG Y Q, SUN Z A, LIU J G, et al. Flavone ingredients can synergistically inhibit NDV infecting cell and improve ND vaccine’s protective rate[J].InternationalJournalofBiologicalMacromolecules, 2012, 51(3):201-208.
[5] CHEN Y, ZHU J. Anti-HBV effect of individual traditional Chinese herbal medicineinvitroandinvivo: an analytic review[J].JournalofViralHepatitis, 2013, 20(7):445-452.
[6] GUO M, ZHANG N, LI D, et al. Baicalin plays an anti-inflammatory role through reducing nuclear factor-κB and p38 phosphorylation inS.aureus-induced mastitis[J].InternationalImmunopharmacology, 2013, 16(2):125-130.
[7] NAYAK M K, AGRAWAL A S, BOSE S, et al. Antiviral activity of baicalin against influenza virus H1N1-pdm09 is due to modulation of NS1-mediated cellular innate immune responses[J].JournalofAntimicrobialChemotherapy, 2014, 69(5):1298-1310.
[8] ZHU J L, WANG J F, SHENG Y, et al. Baicalin improves survival in a murine model of polymicrobial sepsis via suppressing inflammatory response and lymphocyte apoptosis[J].PLoSOne, 2012, 7(5):e35523.
[9] 劉曉婷. 黃芩苷對流感病毒H1N1感染A549細胞的作用及免疫調控機制的研究[D]. 北京:北京中醫藥大學, 2016.
LIU X T. Effect of Baicalin on influenza virus H1N1 infection in A549 cells and its immunoregulation mechanism[D]. Beijing: Beijing University of Chinese Medicine, 2016.(in Chinese with English abstract)
[10] JIA Y, XU R G, HU Y C, et al. Anti-NDV activity of baicalin from a traditional Chinese medicineinvitro[J].JournalofVeterinaryMedicalScience, 2016, 78(5):819-824.
[11] KUMAR H, KAWAI T, AKIRA S. Biochemical and biophysical research communications toll-like receptors and innate immunity[J].BiochemicalandBiophysicalResearchCommunications, 2009, 388(4):621-625.
[12] 何曉霞, 白海. Toll樣受體的研究進展[J]. 西北國防醫學雜志,2010,31(6):445-446.
HE X X, BAI H. Advances in toll like receptors[J].MedicalJournalofNationalDefendingForcesinNorthwestChina, 2010,31(6):445-446.(in Chinese)
[13] 丁娜, 孫英杰, 丁鏟. 禽Toll樣受體研究進展[J]. 中國動物傳染病學報,2011,19(6):77-83.
DING N, SUN Y J, DING C. Research progress in avian toll-like receotors[J].ChineseJournalofVeterinaryParasitology, 2011, 19(6):77-83.(in Chinese with English abstract)
[14] VERMA A, PRASAD K N, SINGH A K, et al. Evaluation of the MTT lymphocyte proliferation assay for the diagnosis of neurocysticercosis[J].JournalofMicrobiologicalMethods, 2010, 81(2):175-178.
[15] CHEN M Z, XIE H G, YANG L W, et al.Invitroanti-influenza virus activities of sulfated polysaccharide fractions fromGracilarialemaneiformis[J].VirologicaSinica, 2010, 25(5):341-351.
[16] 張婷婷. 兩種不同來源新城疫病毒的感染對雞免疫作用的分子機制[D]. 哈爾濱:東北農業大學, 2015.
ZHANG T T. The molecular mechanism of infection ofNewcastleDiseaseVirusfrom different hosts on theimmune function of chickens[D]. Harbin: Northeast Agricultural University, 2015.(in Chinese with English abstract)
[17] AKIRA S, UEMATSU S, TAKEUCHI O. Pathogen recognition and innate immunity[J].Cell, 2006, 124(4):783-801.
[18] KURTJONES E A, BELKO J, YU C, et al. The role of toll-like receptors in herpes simplex infection in neonates[J].JournalofInfectiousDiseases, 2005, 191(5):746-748.
[19] ZHOU S, HALLE A, KURTJONES E A, et al.Lymphocyticchoriomeningitisvirus(LCMV) infection of CNS glial cells results in TLR2-MyD88/Mal-dependent inflammatory responses[J].JournalofNeuroimmunology, 2008, 194(1/2):70.
[20] SEGOVIA J, SABBAH A, MGBMENA V, et al. TLR2/MyD88/NF-κB pathway, reactive oxygen species, potassium efflux activates NLRP3/ASC inflammasome during respiratory syncytial virus infection[J].PLoSOne, 2012, 7(1):e29695.
[21] PAULINE MENAGER P R F M. Toll-like receptor 3 (TLR3) plays a major role in the formation of rabies virus Negri Bodies[J].PLoSPathogens, 2009, 5(2):e1000315.
[22] ALEXOPOULOU L, HOLT A C, MEDZHITOV R, et al. Recognition of double-stranded RNA and activation of NF-kappaB by toll-like receptor 3[J].Nature, 2001, 413(6857):732-738.
[23] CHENG J, SUN Y, ZHANG X, et al. Toll-like receptor 3 inhibitsNewcastlediseasevirusreplication through activation of pro-inflammatory cytokines and the type-1 interferon pathway[J].ArchivesofVirology, 2014, 159(11):2937-2948.
[24] TAKEUCHI O, AKIRA S. MyD88 as a bottle neck in Toll/IL-1 signaling[J].CurrentTopicsinMicrobiology&Immunology, 2002, 270:155-167.
[25] YAMAMOTO M, SATO S, HEMMI H, et al. TRAM is specifically involved in the toll-like receptor 4-mediated MyD88-independent signaling pathway[J].NatureImmunology, 2003, 4(11):1144-1150.
[26] TOWN T, BAI F W, WANG T, et al. Toll-like receptor 7 mitigates lethal West Nile encephalitis via interleukin 23-dependent immune cell infiltration and homing[J].Immunity, 2009, 30(2):242-253.
[27] BAKER D G, WOODS T A, BUTCHI N B, et al. Toll-like
receptor 7 suppresses virus replication in neurons but does not affect viral pathogenesis in a mouse model of Langat virus infection[J].JournalofGeneralVirology, 2013,94(Pt 2):336-347.
[28] KLEIN K P, TAN L, WERKMAN W, et al. The role of toll-like receptors in regulating the immune response against respiratory syncytial virus[J].CriticalReviewsinImmunology, 2009, 29(6):531-550.
EffectofBaicalinonexpressionofToll-likereceptor2,3,4,7inNewcastlediseasevirus-infectedCEFs
XU Ruiguang1, JIA Yan2, HU Liwen1, WENG Jiahua1, DENG Junliang1, HU Yanchun1,*
(1.KeyLaboratoryofAnimalDiseaseandHumanHealthofSichuanProvince,CollegeofVeterinaryMedicine,SichuanAgriculturalUniversity,Chengdu611130,China; 2.AgriculturalandEnvironmentalBranch,JiaxingVocationalTechnicalCollege,Jiaxing314036,China)
The purpose of this study was to investigate the changes of the expression ofTLR2,TLR3,TLR4 andTLR7 gene in NDV-infected cells and the effect of Baicalin on them. NDV infection of chicken embryo fibroblasts (CEFs) was performed. Cytotoxicity of Baicalin was tested by the MTT method. Quantitative expressions ofTLR2,TLR3,TLR4 andTLR7 gene were analyzed by real-time fluorescence quantitative PCR (RTFQ PCR). The results showed that the expressions ofTLR2,TLR4 andTLR7 genes were suppressed by NDV in the early infection stage, while their expression were enhanced with the replication of NDV and the expression ofTLR2 andTLR4 gene reached the peak level at 24 h. As well as the expression ofTLR7 gene reached the peak level at 16 h. The infection of NDV could change the expression ofTLR3 gene, but it was not regular. Besides, Baicalin could increase the expression ofTLR2 andTLR4 genes in NDV-infected cells and show the dosage-dependence, which enhanced immune function and possessed anti-NDV activity.
Baicalin;Newcastlediseasevirus(NDV); Toll-like receptors (TLRs);real-time fluorescence quantitative PCR (RTFQ PCR)
浙江農業學報ActaAgriculturaeZhejiangensis, 2017,29(12): 1986-1993
http://www.zjnyxb.cn
許銳光,賈艷,胡力文,等. 黃芩甙對新城疫病毒感染雞胚成纖維細胞Toll樣受體2、3、4、7 mRNA表達的影響[J].浙江農業學報, 2017,29(12): 1986-1993.
10.3969/j.issn.1004-1524.2017.12.05
2017-05-22
四川省科技支撐項目(2015SZ0201);教育部創新團隊項目(IRT0848);四川省學術和技術帶頭人培養基金項目(2014,2016)
許銳光(1993—),女,河北滄州人,碩士研究生,研究方向為動物環境公害性疾病。E-mail: 597033704@qq.com
*通信作者,胡延春,E-mail: hychun114@163.com
S831.7
A
1004-1524(2017)12-1986-08
(責任編輯盧福莊)
