BCI 50 hm2森林大樣地2010年活立木空間結構分析
2018-01-03 00:51:48周紅敏王宏翔趙中華惠剛盈
林業科學研究 2017年6期
彭 輝 ,周紅敏 ,王宏翔 ,趙中華 ,惠剛盈*
(1.中國林業科學研究院林業研究所,國家林業局林木培育重點實驗室,北京 100091;2.浙江鳳陽山自然保護區管理處,浙江 龍泉 323700; 3.龍泉市林業科學研究院,浙江 龍泉 323700)
BCI50hm2森林大樣地2010年活立木空間結構分析
彭 輝1,2,周紅敏3,王宏翔1,趙中華1,惠剛盈1*
(1.中國林業科學研究院林業研究所,國家林業局林木培育重點實驗室,北京 100091;2.浙江鳳陽山自然保護區管理處,浙江 龍泉 323700; 3.龍泉市林業科學研究院,浙江 龍泉 323700)
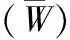
BCI樣地;空間結構;二元分布
森林結構是森林動態變化過程中測度時點的林分狀態的高度概括和度量。隨著森林可持續經營對森林精確信息的需求增加,森林空間結構研究越顯重要[1-3]。林分空間結構決定林木間的競爭態勢及其空間生態位,在很大程度上決定林分的穩定性、發展的可能性和經營空間的大小[4-7]。目前,在森林結構解析中,基于相鄰木空間關系的林分空間結構分析方法已被廣泛應用,由于其可釋性、簡潔性和可操作性等優點,已經廣泛應用在森林類型的研究中,為結構化經營提供了科學依據[8]。森林固定監測大樣地在前所未有的尺度上為研究種群動態等生態學規律和現象提供了良好的契機[9],是當前生態學研究中最活躍的領域之一[10-13]。基于最近相鄰木關系的林分空間結構參數角尺度、混交度和大小比數等在國內外關于林分空間結構分析、林木競爭與優勢度計算、物種多樣性測度以及結構恢復重建與優化調整等研究目前主要是以1 hm2或更小面積的樣地作為研究對象,在全球的森林大樣地中應用較少。各國學者對位于巴拿馬的BCI 50 hm2的大樣地進行了大量的研究,在生物多樣性的維持機制、物種的點格局和群落動態等方面產生了巨大的影響[14-17]。但對BCI 50 hm2大樣地空間結構一元及二元分布特征的分析卻鮮有報道。因此,作者以BCI 50 hm2大樣地為研究對象,利用空間結構參數一元分布和二元分布,量化評價BCI 50 hm2大樣地(2010年第7次普查)的活立木空間結構特征,探討空間結構參數在大樣地中的應用前景以及對空間結構參數理論體系進行進一步驗證。
1 材料
分析采用BCI樣地第7次(2010年)的普查數據資料。數據來源網絡以R.data形式保存的公開數據(http://dx.doi.org/10.5479/data.bci.20130603),數據的詳細信息及全部物種名錄詳見:https://repository.si.edu/handle/10088/20925。

2 研究方法
BCI樣地數據存儲的格式為 Plotname.full#.rdata,#代表調查的次數,如第7次調查(2010年)文件的名稱為bci.full7.rdata。每次調查的數據框(dataframe)包含著相同的行數(number of rows), 其中“treeID”行包含了所有調查過的樹木的編號,無論其補員(Recruitment)、存活(Alive)或者死亡(Dead)都會永久保留其標識號,因為行數上的匹配,歷次的調查數據可以方便的進行比較分析。本研究以樣地中DBH≥1 cm的活立木(Alive)為分析對象,并剔除數據中的缺失值(NA)。BCI樣地第7次調查中活立木有297個物種,共201 434株林木。其中冠層包含180個物種7 410株林木,占總物種數的60%,占總株數的3.68%;下層有290個物種194 024株林木,占總物種數的97.6%,占總株數的96.3%。
在熱帶森林中,一般以胸徑(DBH)超過一定標準作為冠層的劃分,例如Hubbell和Fosetr[21]以20 cm劃分冠層,Steege等以30 cm作為冠層[24],La Frankie等以40 cm作為冠層[25]等,本文將BCI樣地第7次調查數據中所有活立木個體分為冠層林木(DBH≥20 cm)和下層林木(1 cm≤DBH<20 cm)進行空間結構分析。
采用R 3.3.2軟件分析,Excel 2013作圖,分析BCI樣地整體林分以及冠層和下層活立木的一元和二元分布空間特征。空間結構二元分布是基于最近 4 株相鄰木的空間關系構建的林分空間結構分析方法,可從兩個方面同時解析林分空間結構的微觀信息,結合空間結構一元分布對林分宏觀特征的解析,其理論基礎是基于有直觀明了的空間解析能力以及彼此獨立且有相同的結構劃分的角尺度 (W) ,混交度(Mingling,M) 和大小比數(U) 3個空間結構參數,其計算方法詳見文獻[26-27],為避免邊緣效應對林分結構的影響,計算時緩沖區設置為5 m。
3 結果與分析
3.1 BCI大樣地空間結構一元分布特征
3.2 BCI大樣地空間結構的二元分布特征
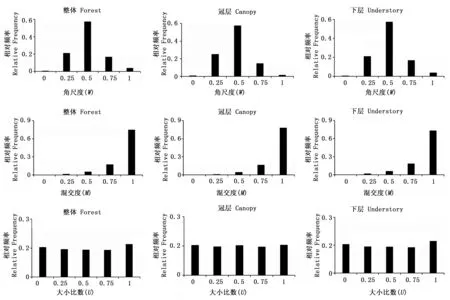
圖1 BCI樣地第7次(2010年)普查空間結構一元分布Fig.1 The univariate distribution of spatial structure in BCI plot
3.2.3 BCI林分的混交度和角尺度的二元分布 在W-M的組合中(圖2C),混交度的整體分布趨勢隨著角尺度的增大表現為先增大后減小,均為正態分布。在隨機分布軸(Wi=0.50)上,混交度頻率值之和最大,為57.4%,在均勻分布狀態(Wi=0.25)時,頻率值為21.2%,兩者的頻率值總和為78.6%。在很均勻分布(Wi=0),林木株數最少,所占頻率值為0.7%,其次為聚集(很不均勻)分布狀態(Wi=1),頻率值為3.8%。在混交度(Mi=0.00)和角尺度(Wi=0.00)的組合上頻率值為0.000 02,表明只有5株林木在零度混交時均勻分布。最大頻率值出現在混交度(Mi=1.00)和角尺度(Wi=0.50)的結構組合上,頻率值為42.9%,即樣地中大部分林木個體處于隨機分布且極強度混交狀態,這也從參照樹同種的最近鄰體(Mi=0.00→0.50)的分布頻率不足8%得到證明。
3.3 BCI大樣地冠層的空間結構二元分布
以中度混交軸(Mi=0.50)和中庸軸(Ui=0.50)為界(圖2D),在零度混交(Mi=0.00)時,林木株數幾乎為0,在低混交等級(Mi=0.00→0.25)上,冠層林中優勢木(Ui=0.00,Ui=0.25)頻率幾乎為0。說明冠層林木中參照樹周圍最近4株相鄰木中極少同種的個體存在。而在高混交等級(Mi=0.75→1.00)時,優勢木頻率最高。在高度混交(Mi=1.00)時,各等級大小比數林木株數均較多,達到78%。在同等級大小比數上,林木株數隨著混交度的增大而增大。
由圖2E可知,隨著大小比數等級的增加角尺度頻率值為先增大后減小,大小比數均在Wi=0.5時頻率值最大,各等級大小比數之和在Wi=0.5軸上的總和為57.5%,冠層中各等級大小比數的林木株數差異不顯著。從圖2F可知,在相同的混交度等級上,角尺度的頻率值為正態分布,即先增大后減小。在Wi=0.50時,混交度各頻率值之和最大為57.5%,說明冠層有1/2以上的林木屬于隨機分布。而Wi=0.00和Wi=1.00的比例為1%和1.6%,說明BCI樣地冠層林木個體中較少處于很均勻和很不均勻分布的狀態。
3.4 BCI大樣地下層的空間結構二元分布
BCI樣地(2010年)下層林木的混交度和大小比數(M-U)的組合中(圖2G),零度混交軸(M=0.00)的大小比數的頻率值之和接近于0,其余大部分林木處于高度混交狀態。混交度和大小比數的最大頻率值為(Mi=1.00,Ui=0.00)的組合為16.4%,即下層中處于極強度混交的劣勢木的株數較多。圖2H為下層林木角尺度和大小比數(W-U)的二元分布,各等級角尺度的林木株數隨著大小比數等級的增加而增大,說明下層中處于劣勢狀態的林木較多。各等級大小比數的頻率值與林分整體和冠層的趨勢相似,也表現為正態分布。而且在隨機分布狀態時,頻率值之和最大,為57.4%,處于很均勻分布和聚集分布的林木較少,分別為0.7%和3.8%。下層的林木個體處于零度混交的株數極少(圖2I),處于弱度混交(Mi=0.25)的林木個體株數僅占下層總株數的1.8%。各等級混交度的頻率值隨著角尺度等級的增大表現為先增加后減小,在隨機分布狀態(Wi=0.50)時,混交度頻率值之和最大為57.4%。混交度和角尺度的最大頻率值出現在(Mi=1.00,Wi=0.50)的組合上為42.1%,為其它24個組合頻率均值的17.5倍。
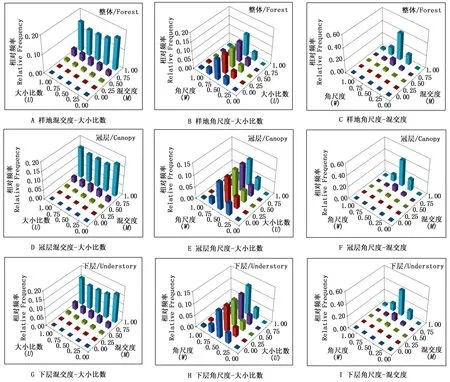
圖2 BCI樣地第7次(2010年)普查空間結構二元分布Fig.2 The bivariate distribution of spatial structure in BCI plot
4 討論
(1)在自然條件下,種群的結構和空間分布往往是種群生物學特性、種內種間關系以及環境條件長期相互適應、相互作用的結果[14]。采用基于最近相鄰木的空間結構參數對BCI大樣地第7次調查(2010年)活立木進行分析,林分整體表現為隨機分布。在混交度方面,整體林分表現為大部分林木處于強度和極強度混交狀態,而且處于極強度混交的優勢木株數較多。樣地內大多數林木DBH在5 cm以下,說明林分中幼苗儲備充足,樣地中處于劣勢的林木較多。
(2)以BCI整體林分為例,通過一元分布可知林分中林木個體大多數處于極強度混交,但不知處于極強度混交的林木個體處于哪種分布格局或在林分中處于怎樣的態勢。通過混交度-大小比數(圖2A)可知BCI樣地中處于極強度混交狀態的林木在大小比數各等級的林木株數相差不大;角尺度-混交度(圖2C),可以很直觀的展示處于極強度混交狀態的林木個體在隨機分布狀態的頻率值為0.429,說明林分中處于極強度混交狀態的林木大部分處于隨機分布狀態。
5 結論
(1)BCI 樣地(2010年)整體林分為隨機分布格局,處于很均勻分布的林木較少。林分處于極強度混交狀態,處于中低度混交等級的林木較少,在零度、弱度和中度3個等級的頻率之和不足8%。處于零度混交狀態且處于均勻分布的林木最少(頻率值為0.000 02),大部分林木處于隨機分布狀態且極強度混交狀態。林木株數在各大小比數等級上的頻率差別不大,處于極強度混交和絕對優勢狀態的林木較多,占總株數的17%。處于絕對劣勢狀態且隨機分布的林木株樹最多。
(2)BCI 樣地(2010年)冠層林木大多數處于高度混交狀態,在零度混交狀態的林木極少。在同一大小比數等級上,不同混交狀態的林木株數存在顯著差異。冠層林木大多數處于隨機分布狀態,在團狀聚集和很均勻分布狀態的林木較少。冠層林木在各等級優劣程度上的株樹相差不大,大致為1∶1∶1∶1。
(3)BCI 樣地(2010年)下層林木分布格局為隨機分布,大部分林木處于高度混交狀態,處于弱度混交狀態的林木株樹也較少,處于極強度混交狀態且處于隨機分布狀態的林木株數是其它混交度-角尺度組合的17.5倍。下層林木中處于劣勢狀態的林木株數相對較多。
綜上所述, BCI 樣地(2010年)林分整體、冠層林木、下層林木分布格局均表現為隨機分布。林分整體、冠層林木和下層林木大部分林木均處于極強度混交狀態,處于絕對劣勢的林木株數最多,處于劣勢的林木株數相對較少;整體表現為中庸偏劣勢的狀態。
[1] 趙中華,惠剛盈,胡艷波,等. 基于大小比數的林分空間優勢度表達方法及其應用[J]. 北京林業大學學報, 2014, 36(1):78-82.
[2] Pommerening A. Approaches to quantifying forest structures[J]. Foresty, 2002, 75(3):305-324.
[3] 周紅敏,彭 輝,柳新紅,等.萌生杉木林經營后林分空間結構變化[J]. 浙江林業科技, 2015, 35(4):41-46.
[4] 巫志龍,周成軍,周新年,等. 杉闊混交人工林林分空間結構分析[J]. 林業科學研究, 2013, 26(5):609-615.
[5] 馬 琳,梁 軍,周國英,等. 昆崳山天然赤松次生林空間結構特征的研究[J]. 中南林業科技大學學報, 2013, 33(9):50-54.
[6] 周紅敏,惠剛盈,趙中華,等. 森林結構調查中最適樣方面積和數量的研究[J]. 林業科學研究, 2009, 22(4):482-495.
[7] 惠剛盈,李 麗,趙中華. 林木空間分布格局分析方法[J]. 生態學報, 2007, 27(11):4717-4728.
[8] 周紅敏,何必庭,彭 輝,等.萌生杉木林空間結構特征研究[J]. 林業科學研究, 2015, 28(5):686-690.
[9] Chisholm R A, Condit R, Rahman K A,etal. Temporal variability of forest communities: empirical estimates of population change in 4000 tree species[J]. Ecology Letters, 2014, 17(7):855-65.
[10] Hubbell S P, Ahumada J A, Condit R,etal. Local neighborhood effects on long-term survival of individual trees in a Neotropical forest[J]. Ecological Research, 2001, 16(5):859-875.
[11] Wang X G, Wiegand T, Wolf A,etal. Spatial patterns of tree species richness in two temperate forests[J]. Journal of Ecology, 2011, 99(6):1382-1393.
[12] 馬克平.生物多樣性與生態系統功能的實驗研究[J]. 生物多樣性, 2013, 21(3): 247-248.
[13] 王慧杰,常順利,張毓濤,等. 天山雪嶺云杉森林群落的密度制約效應[J]. 生物多樣性, 2016, 24(3):252-261.
[14] Condit R, Ashton P S,Baker P,etal.Spatial patterns in the distribution of tropical species[J]. Science, 2000, 288(5470):1414-1418.
[15] Condit R, Ashton P, Bunyavejchewin S,etal. The importance of demographic niches to tree diversity[J]. Science, 2006, 313(5783):98.
[16] Nathan R. Long-distance dispersal of plants[J]. Science, 2006, 313(5788):786-788.
[17] 郝占慶,李步杭,張 健,等.長白山闊葉紅松林樣地(CBS):群落組成與結構[J]. 植物生態學報, 2008, 32 (2):238-250.
[18] Bohlman S A. Species Diversity of Canopy versus Understory Trees in a Neotropical Forest: Implications for Forest Structure, Function and Monitoring[J]. Ecosystems, 2015, 18(4):658-670.
[19] Croat T R. Flora of Barro Colorado Island[M]. Stanford, CA: Stanford University Press, 1978.
[20] Leigh E G, Jr Rand S A, Windsor D M. The Ecology of a Tropical Forest: Seasonal Rhythms and Long- Term Changes[M]. Washington, DC: Smithsonian Institution Press, 1982.
[21] Hubbell S P, Foster R B. Diversity of canopy trees in a neotropical forest and implications for conservation[M]// Sutton S L, Whitmore T C, Chadwick A C. Tropical Rain Forest: Ecology and Management. Oxford: Blackwell Scientific Publications, 1983: 25-41.
[22] Condit R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and Comparison with Other Plots[M]. Berlin: Springer, 1998.
[23] Condit R, Chisholm R A, Hubbell S P. Thirty years of forest census at Barro Colorado and the importance of immigration in maintaining diversity[J]. PLoS ONE, 2012, 7(11):1-6.
[24] Steege H, Pitman N C A, Sabatier D,etal. Hyperdominance in the Amazonian tree flora[J]. Science, 2013, 342(6156):1243092.
[25] LaFrankie J V, Ashton P S, Chuyong G B,etal. Contrasting structure and composition of the understory in species-rich tropical rain forests[J]. Ecology, 2006, 87(9):2298-305.
[26] LI Y F,HUI G Y,ZHAO Z H,etal.The bivariate distribution characteristics of spatial structure in natural Korean pine broadleaved forest[J].Journal of Vegetation Science, 2012, 23(6):1180-1190.
[27] 李遠發. 林分空間結構參數二元分布的研究[D].北京: 中國林業科學研究院, 2013.
[28] Wiegand T, Moloney K A.Rings, circles and null-models for point pattern analysis in ecology[J]. Oikos, 2004, 104(2):209-229.
[29] Wright S J. Plant diversity in tropical forests: a review of mechanisms of species coexistence[J]. Oecologia, 2002, 130(1):1-14.
[30] Barot S, Gignoux J, Menaut J C. Demography of a savanna palm tree: predictions from comprehensive spatial pattern analyses[J]. Ecology, 1999, 80(6):1987-2005.
ResearchonSpatialStructureof50hm2ForestPlotatBarroColoradoIslandin2010
PENGHui1,2,ZHOUHong-min3,WANGHong-xiang1,ZHAOZhong-hua1,HUIGang-ying1
(1.Key Laboratory of Tree Breeding and Cultivation, State Forestry Administration; Research Institute of Forestry,Chinese Academy of Forestry, Beijing 100091, China; 2.Fengyangshan National Nature Reserve, Longquan 323700, Zhejiang, China; 3.Longquan Forestry Research Institute, Longquan 323700, Zhejiang, China)
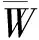
BCI plot;spatial structure;bivariate distribution
10.13275/j.cnki.lykxyj.2017.06.002
2017-03-20
國家自然科學基金項目(31370638)
彭 輝,工程師。
* 通訊作者:惠剛盈,研究員。E-mail:hui@caf.ac.cn
S718.54
A
1001-1498(2017)06-0895-07
彭南軒)
