何首烏及其主要成分二苯乙烯苷對非酒精性脂肪肝大鼠腸道短鏈脂肪酸產(chǎn)生量的影響△
2017-12-26 01:47:06王艷芳林佩陸建美張美李莉楊興鑫俞捷
中國現(xiàn)代中藥 2017年9期
王艷芳,林佩,陸建美,張美,李莉,楊興鑫,俞捷
(云南中醫(yī)學(xué)院 中藥學(xué)院,云南 昆明 650500)
·基礎(chǔ)研究·
何首烏及其主要成分二苯乙烯苷對非酒精性脂肪肝大鼠腸道短鏈脂肪酸產(chǎn)生量的影響△
王艷芳a,林佩a,陸建美,張美,李莉,楊興鑫,俞捷*
(云南中醫(yī)學(xué)院 中藥學(xué)院,云南 昆明 650500)
目的探討何首烏及其主要成分二苯乙烯苷(TSG)對非酒精性脂肪肝模型大鼠腸道內(nèi)微生物發(fā)酵碳水化合物產(chǎn)生短鏈脂肪酸量的影響。方法SD大鼠隨機分為正常組、高脂飲食組和高脂飲食給藥組(分別給予何首烏水提物和 TSG)。采用氣相色譜法檢測各組大鼠糞便中短鏈脂肪酸(以乙酸、丙酸、丁酸為代表)含量。COD-PAP法測定血液和肝臟總膽固醇(TC)、甘油三酯(TG)含量,鱟試驗法檢測肝門靜脈血內(nèi)毒素。結(jié)果何首烏總提物及TSG各劑量都能不同程度的降低高脂飲食大鼠腸道內(nèi)的總短鏈脂肪酸(SCFA)含量,但其調(diào)節(jié)作用可能存在性別差異。生何首烏及TSG能夠顯著降低高脂飲食雄性大鼠腸道內(nèi)乙酸、丙酸、丁酸含量,同時降低實驗動物肝臟脂質(zhì)含量和內(nèi)毒素水平。生何首烏下調(diào)高脂飲食雌性大鼠腸道內(nèi)丙酸含量,降低肝臟脂質(zhì)水平;而TSG低劑量能夠升高乙酸含量,同時降低血脂和內(nèi)毒素含量。然而制何首烏對雌、雄鼠腸道內(nèi)SCFA含量的調(diào)節(jié)作用均不顯著。結(jié)論何首烏及其活性成分 TSG 能夠調(diào)節(jié)腸道微生物發(fā)酵產(chǎn)生的短鏈脂肪酸含量,這種調(diào)節(jié)活性可能與其對非酒精性脂肪肝的治療作用有關(guān)。
何首烏;二苯乙烯苷;短鏈脂肪酸;非酒精性脂肪肝;氣相色譜
非酒精性脂肪性肝病(nonalcoholic fatty liver disease,NAFLD)是代謝綜合征的肝表現(xiàn),常合并肥胖、血脂異常及胰島素抵抗[1]。近年來,隨著生活方式和飲食結(jié)構(gòu)的改變,NAFLD的患病率范圍從超重個體的57%到非糖尿病肥胖患者的98%[2]。NAFLD發(fā)病機制尚未完全清楚,經(jīng)典解釋為Day等[3]提出的二次打擊學(xué)說:初次打擊主要是胰島素抵抗和脂質(zhì)代謝紊亂所導(dǎo)致的單純性脂肪肝;二次打擊是內(nèi)毒素(lipopolysaccharide,LPS)腸滲漏,引發(fā)氧化應(yīng)激和炎癥反應(yīng),導(dǎo)致脂肪變性的肝細胞發(fā)生炎癥和壞死。越來越多的證據(jù)已經(jīng)表明腸道菌群的改變與肥胖相關(guān)的NAFLD患者和動物模型的發(fā)展相關(guān)聯(lián),認為腸道菌群通過腸-肝軸促進NAFLD的發(fā)展[4]。短鏈脂肪酸(short chain fatty acids,SCFA)多是由腸道菌群發(fā)酵結(jié)腸中未被宿主消化吸收的碳水化合物和蛋白質(zhì)而產(chǎn)生。SCFA是腸道菌群而非宿主的重要代謝產(chǎn)物[5],主要包括乙酸、丙酸、異丁酸、丁酸、異戊酸、戊酸,其中乙酸、丙酸、丁酸所占比例高達85%[6]。SCFA不僅能為腸黏膜細胞提供能量,促進細胞的代謝、生長[7];還可以調(diào)節(jié)腸道內(nèi)環(huán)境pH值,防止腸道功能紊亂[8];此外,SCFA還在調(diào)節(jié)炎癥反應(yīng)[9]、抗腫瘤[10]和調(diào)控基因表達[8]等方面發(fā)揮重要的作用。同時,有文獻報道稱腸道菌群主要代謝產(chǎn)物SCFA可能通過多種途徑(腸道、脂肪組織、肝臟)介導(dǎo)機體炎癥反應(yīng),直接或間接影響NAFLD的發(fā)生發(fā)展[11]。
本課題組前期研究發(fā)現(xiàn),何首烏水提物及二苯乙烯苷(2,3,5,4′-tetrahydroxy-stilbene-2-O-β-D-glucoside,TSG)能抑制NAFLD大鼠腸道內(nèi)厚壁菌門與擬桿菌門比例的升高,調(diào)節(jié)腸道菌群組分的比例,改善腸道微生態(tài)紊亂狀況,抑制后期腸源性內(nèi)毒素等一系列炎癥因子的釋放,表明何首烏水提物及TSG可在NAFLD發(fā)病的二次打擊環(huán)節(jié)中起到改善調(diào)節(jié)作用[12]。依據(jù)本課題組前期研究基礎(chǔ),我們推測何首烏水提物及TSG良好的脂代謝調(diào)節(jié)活性可能與其影響腸道內(nèi)SCFA含量相關(guān)。為了闡明脂代謝調(diào)節(jié)與SCFA之間可能的關(guān)系,進一步發(fā)掘何首烏水提物及TSG的降血脂機制,我們就其對高脂飲食誘導(dǎo)NAFLD模型大鼠腸道內(nèi)SCFA含量的影響展開了研究。
1 材料
1.1 動物與飼料
清潔級健康Spraque-Dawley(SD)大鼠,(180±20)g,雌雄各半,購自成都達碩生物科技有限公司(編號:0016254);實驗條件和方法經(jīng)云南中醫(yī)學(xué)院動物實驗中心倫理委員會審查合格(動物倫理學(xué)審查文件編號:R-062014012)。
基礎(chǔ)飼料配方:面粉20%、米粉10%、玉米20%、麩皮25%、豆料20%、骨粉2%、魚粉2%。高脂飼料配方:基礎(chǔ)飼料79%、膽固醇1%、豬油10%、蛋黃10%。
1.2 材料與試劑
實驗所用的何首烏購自云南省祿勸縣,經(jīng)云南中醫(yī)學(xué)院俞捷副教授鑒定為蓼科植物何首烏P.multiflorumThunb.的干燥塊根。樣品存于云南中醫(yī)學(xué)院中藥炮制研究中心。按《中華人民共和國藥典》2015版記載方法用黑豆汁對生何首烏進行炮制得到制何首烏。經(jīng)高效液相色譜法測定,實驗用生何首烏及炮制品中二苯乙烯苷質(zhì)量分數(shù)分別為5.99%及2.94%,均大于1.0%,符合《中華人民共和國藥典》標準。
TSG(南京景竹生物科技有限公司,經(jīng)HPLC檢測純度為98%);標準品乙酸(A116165)、丙酸(D110443)、丁酸(B110439)、己酸(H103630)均為色譜純(上海晶純生化科技股份有限公司),己酸為內(nèi)標物;二水合草酸(四川西隴化工有限公司);疊氮化鈉(美國Amresco公司);甘油三酯(Triglycerides,TG)試劑盒(COD-PAP法,中生北控生物科技股份有限公司);總膽固醇(Cholesterol,CHO)試劑盒(COD-PAP法,中生北控生物科技股份有限公司);內(nèi)毒素定量鱟試劑盒(廈門市鱟試劑實驗廠有限公司)。
1.3 儀器與設(shè)備
GC 7890B型氣相色譜(美國安捷倫科技公司);HR/T16M型臺式高速冷凍離心機(湖南赫西儀器裝備有限公司);TP 214型電子天平(北京賽多利斯儀器有限公司);ZHWY-103D型恒溫振蕩器(上海智城分析儀器制造有限公司);Milli-Q 超純水機(法國Millipore SAS);DNM-9602G型酶標分析儀(北京普朗新技術(shù)有限公司);DY89-Ⅱ型電動玻璃勻漿機(寧波新芝生物科技股份有限公司)。
2 方法
2.1 樣品的制備
稱取150 g生何首烏和315 g制何首烏,分別加10倍量水煎煮1 h,過濾,
藥渣加8倍量水煎煮40 min,過濾,藥渣再加6倍量水煎煮40 min,過濾,3次濾液合并,濃縮至流浸膏即可。得生何首烏水提濃縮液為600 mL,制何首烏水提濃縮液560 mL;即生何首烏相當于原生藥濃度0.250 g·mL-1,制何首烏相當于原生藥質(zhì)量濃度0.563 g·mL-1。
2.2 動物分組及給藥
取SD大鼠98只,雄雌各半,實驗前喂養(yǎng)基礎(chǔ)飼料7 d后,空腹稱重。根據(jù)體重、性別隨機分為7組,每組雌雄各7只:正常對照組(CON)、高脂對照組(MOD)、TSG低劑量組(TSG.L)、TSG中劑量組(TSG.M)、TSG高劑量組(TSG.H)、生何首烏水提液組(PMR)、制何首烏水提液組(PMRP)。實驗大鼠自由進食及飲水,實驗期為12周。除CON組喂食基礎(chǔ)飼料外,各組均連續(xù)喂食高脂飼料至實驗結(jié)束。TSG.L組、TSG.M組和 TSG.H組分別按12、24、48 mg·kg-1的劑量灌胃給予二苯乙烯苷單體。PMR組、PMRP組分別按405、810 mg·kg-1的劑量灌胃給予生、制何首烏水提液(按《中華人民共和國藥典》2015版劑量分別為4.5、9.0 g折算)。
2.3 血液和肝臟組織樣品的采集及保存
采用眼內(nèi)眥取血的方法,實驗開始時采集1次,給藥后每隔6天采集1次,每次每只取血量約為1.5~2.0 mL,所取血液用高速臺式冷凍離心機以10 000 r·min-1的轉(zhuǎn)速、4 ℃離心15 min,取上清液采用試劑盒測定TC、TG 含量。
實驗12周后,以水合氯醛溶液麻醉大鼠,使用一次性真空靜脈采血管,采取肝門靜脈血,2000 r·min-1離心10 min取上層血漿,采用鱟試劑檢測內(nèi)毒素(LPS)含量。而后取出大鼠肝臟,稱重,取部分肝臟剪碎并稱取1 g破碎肝臟,置于勻漿管中加入9 mL 0.9%氯化鈉溶液,用電動勻漿機勻漿。勻漿后將離心管放入臺式高速冷凍離心機中,在4 ℃ 4000 r·min-1條件下離心10 min,取上清液,采用試劑盒檢測肝臟細胞中TC、TG的含量。
2.4 糞便樣品的采集及保存
在實驗周期內(nèi),給藥前一天采集1次糞便,給藥后每隔6 d收集1次,于-80 ℃冰箱保存,備用。
2.5 短鏈脂肪酸的測定
2.5.1 標準曲線的制作 取乙酸、丙酸、丁酸對照品溶液分別置于100 mL 容量瓶中,用超純水定容,配制乙酸濃度分別為1.749、5.246、10.49、20.98、26.22 mmol·L-1,丙酸濃度分別為0.534 6、3.207、6.415、12.83、16.04 mmol·L-1,丁酸濃度分別為0.217 9、0.653 7、1.308、2.615、3.269 mmol·L-1。己酸溶液作為內(nèi)標物,其濃度為0.1 mmol·L-1,置于以上各濃度對照品溶液中,混勻,進行氣相色譜分析。以對照品的峰面積與內(nèi)標物峰面積比值(Y)為縱坐標,以濃度(X)為橫坐標,繪制標準曲線。
2.5.2 氣相色譜條件 采用Agilent 122-7032:DB-WAX(30 m×250 μm,0.25 μm)色譜柱進行分析。程序升溫,初溫120 ℃,保持2.5 min,然后以8 ℃·min-1速度升至 130 ℃,保持4 min,再以30 ℃·min-1的速度升至210 ℃,保持2 min。FID溫度為250 ℃,進樣口溫度為200 ℃;進樣量為1 μL。氮氣為載氣,流速為3 mL·min-1;分流比為20∶1[13]。
2.5.3 短鏈脂肪酸的提取 每組每只大鼠各取糞便一粒(162±23.55)g,進行研磨混勻,而后準確稱取糞便樣品500 mg于15 mL離心管中,加入3 mL萃取液(含 0.1 mol·L-1草酸、40 mol·L-1疊氮化鈉和0.1 mol·L-1己酸),將混合物溶液放置于搖床上60 min,然后以12 000 r·min-1轉(zhuǎn)速離心10 min;取上清液經(jīng)有機系0.45 μm微孔濾膜過濾置于安瓿瓶中,進行GC分析。根據(jù)標準曲線計算得出短鏈脂肪酸含量[14]。
2.6 數(shù)據(jù)處理
3 結(jié)果與分析
3.1 標準曲線結(jié)果
在氣相色譜條件下進樣測定,3種SCFA及內(nèi)標物的標準品和樣品色譜圖見圖1,分離效果好。同時3種SCFA的線性關(guān)系結(jié)果列于表1。從線性關(guān)系可以看出,乙酸、丙酸、丁酸在對應(yīng)的濃度范圍內(nèi)線性關(guān)系良好,r均達到0.999 0。
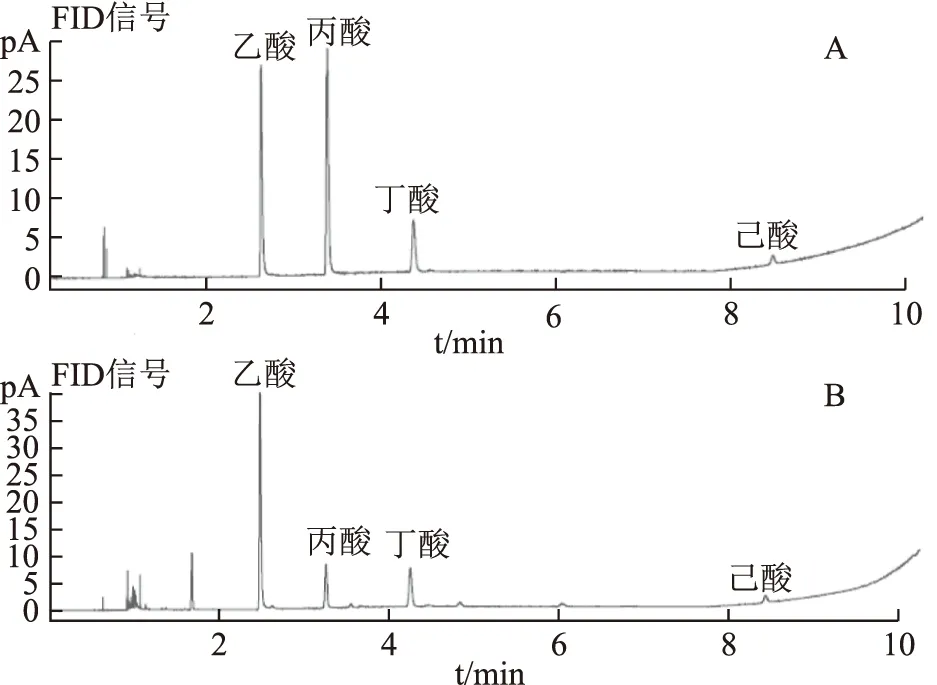
注:A.對照品;B.樣品。圖1 3種SCFA和己酸內(nèi)標的標準品及樣品GC色譜圖
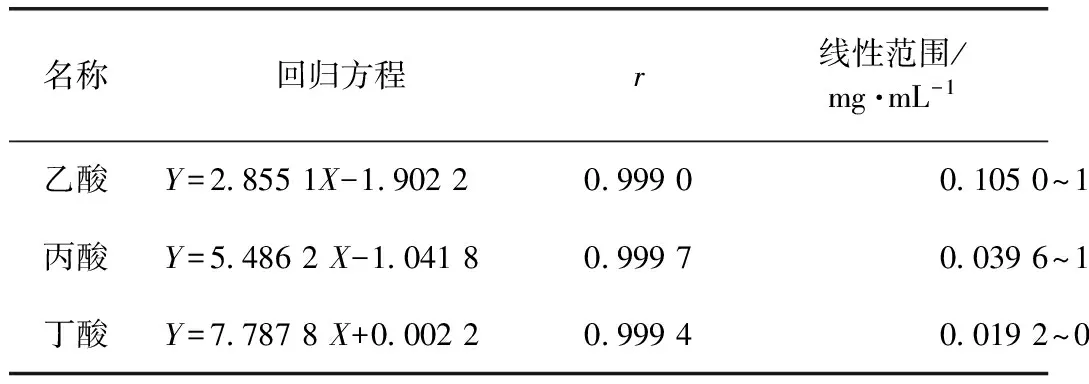
名稱回歸方程r線性范圍/mg·mL-1乙酸Y=28551X-190220999001050~1575丙酸Y=54862X-104180999700396~1188丁酸Y=77878X+000220999400192~0288
3.2 腸道內(nèi)容物SCFA含量與肝臟、血清中TG、TC及LPS含量的相關(guān)性分析
對雌、雄鼠各組腸道內(nèi)SCFA含量與肝臟、血清中的TC、TG及LPS含量進行Pearson相關(guān)性分析,結(jié)果見表2,雄鼠腸道內(nèi)產(chǎn)生的SCFA不僅與肝臟中的TC、TG呈正相關(guān),還與LPS呈正相關(guān),提示雄性大鼠肝臟中脂質(zhì)含量的降低以及內(nèi)毒素水平的下降與腸道內(nèi)SCFA含量的降低有顯著的相關(guān)性。其中乙酸含量與肝臟TG含量相關(guān),r=0.917(P< 0.01);丙酸、丁酸的含量與肝臟中TC、TG及LPS含量呈明顯正相關(guān)(P< 0.01,P< 0.05)。
雌鼠腸道內(nèi)僅乙酸含量與血清中的TC及LPS含量呈負相關(guān),相關(guān)系數(shù)分別為0.927、0.920(P< 0.01),提示腸道內(nèi)容物總SCFA含量的變化與其NAFLD程度無明顯相關(guān)性,但何首烏提取物給藥能使腸道內(nèi)乙酸含量升高,這可能與其降低血清TC含量的功效相關(guān)。
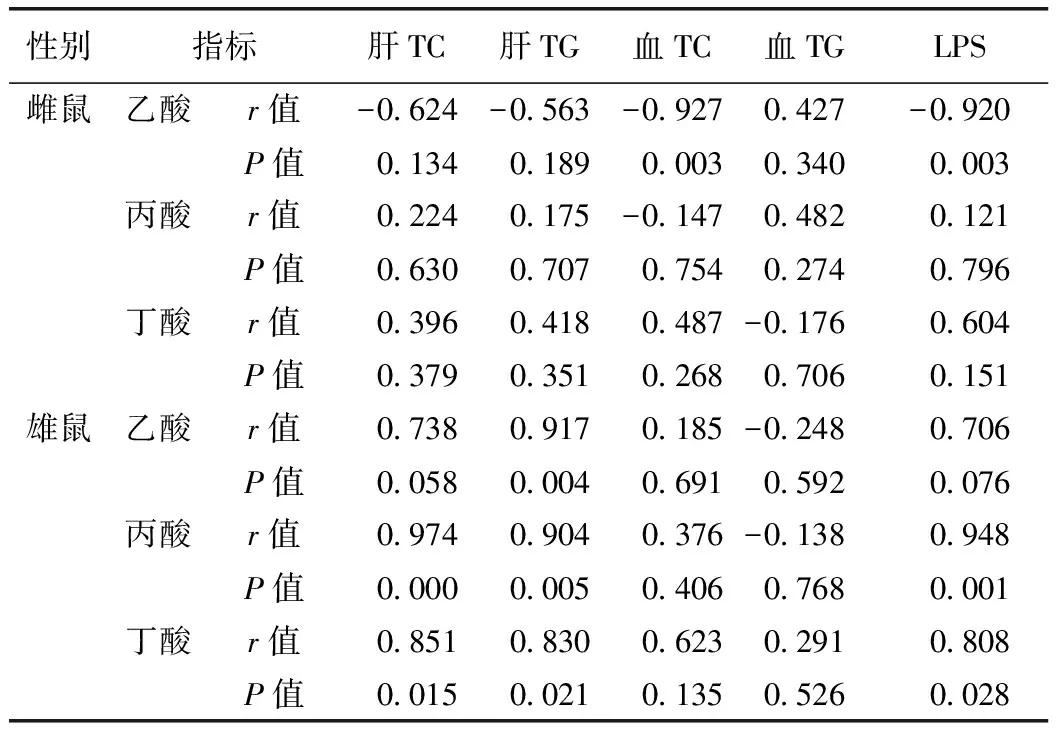
表2 腸道內(nèi)容物SCFA含量與TG、TC及LPS間相關(guān)性分析結(jié)果
注:表中數(shù)值是第12周所有組別指標數(shù)據(jù)進行相關(guān)性分析的結(jié)果。
3.3 何首烏總提物和TSG對血液、肝臟中TC、TG及LPS含量的影響
如表3雌鼠血脂及肝臟脂質(zhì)結(jié)果顯示,飼喂高脂飼料12周后,雌鼠模型組肝臟組織勻漿TC、TG含量分別升高89.8%、40.5%,雌鼠NAFLD模型復(fù)制成功。同時,雌鼠模型組血液中TC、LPS含量顯著高于正常組(P<0.001,P<0.01),但TG含量無明顯變化。藥物治療各組對雌鼠肝臟或血清脂質(zhì)含量均有不同程度的改善,其中以TSG低劑量組對血液中TC、LPS的調(diào)節(jié)改善效果最佳,其次是高劑量組;而生何首烏調(diào)節(jié)肝臟脂質(zhì)含量的效果優(yōu)于其他組,其次是TSG低劑量組。
如表4雄鼠血脂及肝臟脂質(zhì)結(jié)果分析顯示,雄鼠模型組肝臟勻漿TC、TG含量升高幅度分別為81.7%、63.3%,雄鼠NAFLD模型復(fù)制成功。同時,雄鼠血清TC、LPS含量顯著高于正常組(P<0.001,P<0.01)。給予何首烏總提物及TSG治療的非酒精性脂肪肝模型都得到不同程度的改善,其中以生何首烏對肝臟脂質(zhì)含量的調(diào)節(jié)作用最佳,其次是TSG中劑量組。TSG高劑量組調(diào)節(jié)LPS的作用優(yōu)于其他組,其次是生何首烏組。
3.4 SCFA含量測定結(jié)果
3.4.1 何首烏總提物及TSG對雌鼠腸道內(nèi)SCFA含量的影響 實驗選取給藥前(0周)、給藥后1周及實驗最后一周(12周)雌性大鼠糞便樣本進行檢測,雌鼠的食量變化見圖2。如圖3結(jié)果顯示,高脂飼料飼喂1周后,正常組雌鼠腸道內(nèi)乙酸、丙酸、丁酸含量變化幅度不大,而其余各組雌鼠腸道內(nèi)產(chǎn)生的乙酸、丙酸、丁酸含量急劇增加,而這一周高脂飼料組大鼠的攝食量也明顯增加(見圖2),考慮這種腸道內(nèi)產(chǎn)短鏈脂肪酸量的明顯增加可能與高脂飼料在實驗初期對實驗動物攝食行為影響較大相關(guān)。隨著飼喂時間的延長,雌鼠進食量出現(xiàn)下降(見圖2),而到實驗的第12周各組乙酸、丙酸、丁酸的含量均有明顯降低(見圖3),但是組別之間總SCFA的含量受食量影響的關(guān)系不明顯。
表3 雌鼠各組血液及肝臟中TC、TG測定結(jié)果 mmol·L-1
注:與模型組相比,*P<0.05,**P<0.01,***P<0.001;下同。
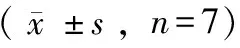
表4 雄鼠各組血液及肝臟中TC、TG測定結(jié)果 mmol·L-1
如圖3所示,與正常組相比,模型組乙酸含量降低,而丙酸、丁酸含量升高。給予藥物治療后,生何首烏降低雌鼠腸道內(nèi)丙酸含量的效果優(yōu)于其他組,降幅為26.67%;低劑量TSG升高雌鼠腸道內(nèi)乙酸含量的效果較佳,比模型組升高18.21%;TSG中劑量降低腸道內(nèi)丁酸含量的效果最佳,較模型組降低63.16%。從調(diào)節(jié)總SCFA的角度來看,TSG低劑量組調(diào)節(jié)效果優(yōu)于高劑量組,而TSG的調(diào)節(jié)作用優(yōu)于總提物。
3.4.2 何首烏總提物及TSG對雄鼠腸道內(nèi)SCFA含量的影響 雄鼠的食量變化見圖2,對雄鼠0周、1周、12周糞便中SCFA含量的檢測結(jié)果如圖4顯示,雄鼠腸道內(nèi)各時間段的總SCFA含量均高于雌鼠腸道內(nèi)的SCFA含量,飼喂高脂飼料1周的雄鼠腸道內(nèi)總SCFA含量顯著高于飼喂正常飼料的大鼠含量。隨著喂養(yǎng)時間的延長,各組大鼠腸道內(nèi)的總SCFA含量均有明顯降低。
綜上,實驗初期雌性、雄性動物腸道內(nèi)SCFA含量受其進食量影響較明顯,而隨著實驗動物對高脂飼料的適應(yīng)及進食量的下降,腸道內(nèi)SCFA含量與進食量間不呈現(xiàn)明顯關(guān)聯(lián)。
第12周末,模型組大鼠腸道內(nèi)乙酸、丙酸、丁酸含量均顯著高于正常組;給予生、制何首烏以及TSG治療后,均能顯著降低 SCFA 的含量,以生何首烏降低乙酸、丙酸、丁酸的效果最佳,降幅分別為56.47%、76.53%、46.94%;其次是TSG低劑量組降低乙酸、高劑量組降低丙酸、中劑量組降低丁酸的效果優(yōu)于其他組。從調(diào)節(jié)總SCFA的角度來看,生何首烏總提物的調(diào)節(jié)作用優(yōu)于TSG,而TSG低劑量組的調(diào)節(jié)效果優(yōu)于中、高劑量組。
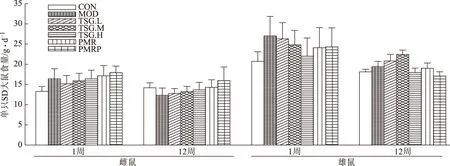
圖2 雌、雄鼠的食量變化
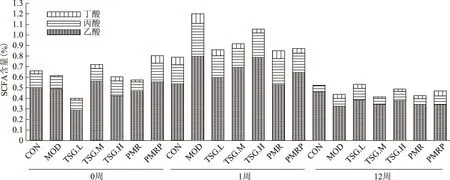
圖3 雌鼠腸道內(nèi)SCFA含量變化
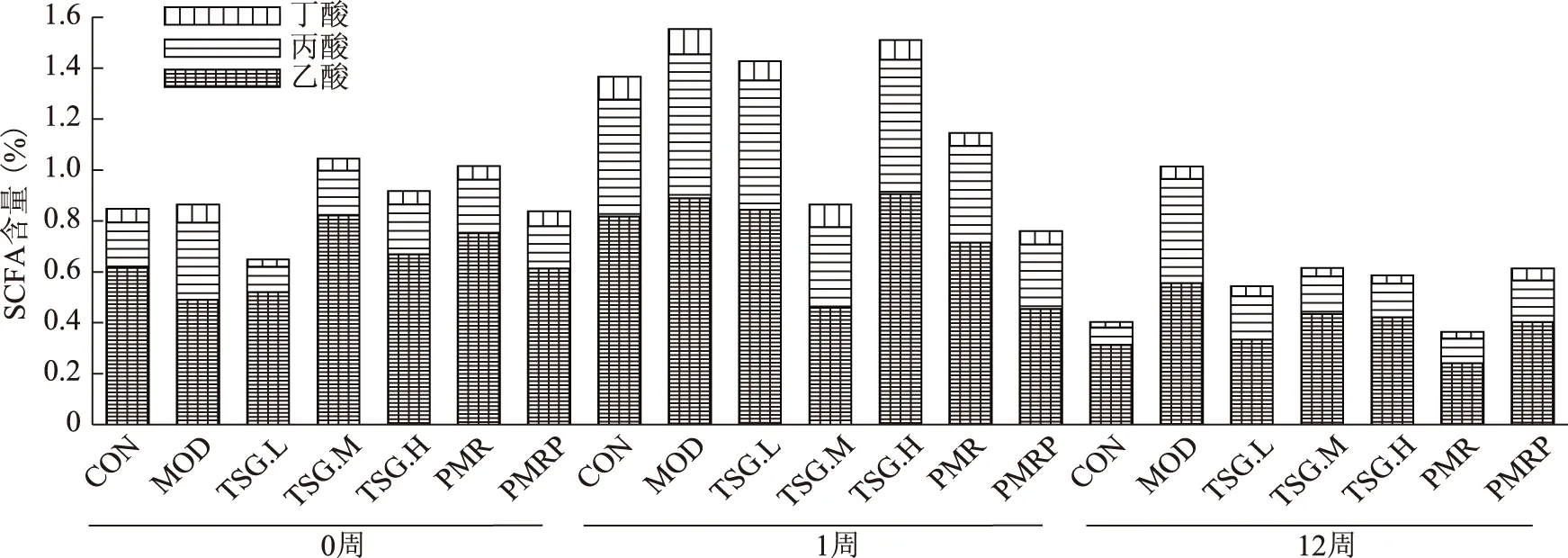
圖4 雄鼠腸道內(nèi)SCFA含量變化
4 討論
SCFA是腸道菌群的主要發(fā)酵產(chǎn)物,在人類結(jié)腸中,正常腸道菌群產(chǎn)生50~100 mmol·L-1的SCFA[15],主要包括乙酸、丙酸、丁酸。相關(guān)文獻顯示關(guān)于SCFA對肥胖和NAFLD發(fā)展的貢獻存在爭議。有文獻報道表明SCFA可能通過多種機制影響肥胖的發(fā)生,同時保護肝細胞。丙酸可使體外培養(yǎng)的肝細胞合成膽固醇的量以及離體肝細胞培養(yǎng)液中HMG-CoA還原酶活性顯著下降[16]。人類炎癥腸道疾病與腸道內(nèi)SCFA的減少有關(guān),炎癥能夠增強腸黏膜通透性,使得菌群產(chǎn)物LPS通過門脈循環(huán)進入肝臟誘發(fā)疾病,因此通過增加SCFA抑制炎癥在保護肝臟健康中扮演了重要角色[17]。Kimura等[18]認為SCFA/GPR43在腸道及腸道外均能發(fā)揮作用,鼠的脂肪組織中SCFA能通過GPR43受體抑制胰島素信號的傳遞,從而抑制脂肪堆積。
另一些研究報道則認為SCFA的增加能夠促進肥胖的發(fā)展。向無菌動物移植腸道菌群的研究結(jié)果顯示,腸道菌群產(chǎn)生的SCFA所提供的能量是從飲食中所攝取能量的30%[19]。乙酸是多數(shù)細菌發(fā)酵的主要代謝產(chǎn)物,也是膽固醇合成的最主要底物,在機體內(nèi),大部分乙酸被吸收入血,進入肝臟的代謝,作為外周組織的能量來源[20]。丙酸在很大程度上由肝臟吸收,是糖異生、脂質(zhì)合成、蛋白質(zhì)合成很好的前體[21-22]。Schwiertz等[23]發(fā)現(xiàn)在肥胖人群的腸道中擬桿菌門與厚壁菌門的比例升高;雖然兩種門類菌群都產(chǎn)生SCFA,但擬桿菌門(如譜氏菌屬)產(chǎn)生的SCFA更多。
另一方面,高脂飼料對雌、雄大鼠脂代謝的影響可能存在性別差異,本課題組前期研究表明高脂飲食誘導(dǎo)使雌、雄鼠血脂異常類型方面表現(xiàn)出顯著的性別差異[24]。本研究結(jié)果也顯示SCFA與NAFLD相關(guān)性可能存在一定性別差異,雄鼠相關(guān)性結(jié)果支持SCFA的增加能夠促進肥胖發(fā)展這一觀點,即雄鼠腸道內(nèi)SCFA量與NAFLD發(fā)病正相關(guān);而雌鼠結(jié)果顯示乙酸含量的增加減少血液TC和LPS含量,表明雌鼠腸道內(nèi)SCFA量可能與NAFLD發(fā)病負相關(guān)。
有研究表明腸道菌群中的腸桿菌、腸球菌、雙歧桿菌、乳酸桿菌等多種菌屬與脂代謝有著密切的關(guān)系,而脂代謝的異常會直接影響腸道菌群的數(shù)量和分布[25];然而腸道內(nèi)發(fā)酵所產(chǎn)生的SCFA的種類和數(shù)量受發(fā)酵基質(zhì)的數(shù)量、類型、降解速率、降解程度,以及腸道菌群和宿主生理狀態(tài)等因素的影響[26]。本研究結(jié)果發(fā)現(xiàn),高脂飲食雄鼠糞便中乙酸、丙酸、丁酸的含量均明顯高于正常組,而高脂飲食雌鼠糞便中乙酸的含量低于正常組;提示高脂飲食大鼠的腸道微生態(tài)系統(tǒng)出現(xiàn)紊亂,可能存在著生成乙酸、丙酸和丁酸的菌屬增加或減少,致使腸道菌群對宿主代謝產(chǎn)生異常影響,引發(fā)代謝紊亂。
給予何首烏總提物及TSG治療后,雄鼠給藥組的總SCFA含量有所降低,其中生何首烏、TSG低劑量組調(diào)節(jié)效果最佳;同時也發(fā)現(xiàn)生何首烏和TSG低劑量組能夠顯著降低肝臟脂質(zhì)及血液內(nèi)毒素。有研究稱短鏈脂肪酸主要被腸上皮細胞吸收,增加體內(nèi)能量的儲備,同時通過門脈循環(huán)進入肝臟,經(jīng)肝臟代謝轉(zhuǎn)變?yōu)槿8视停寡缴撸龠M脂肪的積累[27]。這從降血脂方面對本實驗結(jié)果提供了一個有力的佐證。對于高脂飲食雌鼠,生何首烏顯著降低丙酸含量,TSG低劑量升高乙酸含量,且總SCFA含量趨近于正常組;與此同時生何首烏顯著降低肝臟脂質(zhì),TSG低劑量降低血液中的TC和LPS。近年來,“肝-腸軸”概念的提出為尋找肝病的發(fā)病機制和診療方法提供了新思路。一系列研究顯示,腸源性內(nèi)毒素血癥在NAFLD的發(fā)生發(fā)展中起重要作用[28]。然而本研究在檢測SCFA含量時,由于樣品量不足,通過組內(nèi)合并樣品進行檢測,并且只檢測了一次,結(jié)果沒有標準差值而對組間進行了比較,但是課題組對SCFA含量的測定方法進行了穩(wěn)定性、精密度、相對回收率等方法學(xué)驗證,其結(jié)果均滿足分析要求,表明數(shù)據(jù)的準確性及可靠性。這是本研究的不足之處,在日后的實驗中,應(yīng)多收集糞便樣品或采取所需樣品量少的方法對每個樣品進行SCFA含量的檢測,也許能夠得出更好的結(jié)論。
綜上所述,何首烏總提物及TSG能夠調(diào)節(jié)腸道內(nèi)SCFA含量,促使通過門脈循環(huán)進入肝臟的SCFA適宜,能夠減輕脂質(zhì)在肝細胞內(nèi)的蓄積,阻止腸源性內(nèi)毒素易位,有效防護NAFLD的發(fā)生發(fā)展。然而,SCFA是如何調(diào)控NAFLD宿主外周組織調(diào)控能量代謝、抑制炎癥反應(yīng)的信號轉(zhuǎn)導(dǎo),尚需進一步研究。
[1] Rinella M E,Sanyal A J.NAFLD in 2014:Genetics,diagnostics and therapeutic advances in NAFLD[J].Nat Rev Gastroenterol Hepatol,2015,12(2):65-66.
[2] Vernon G,Baranova A,Younossi Z M.Systematic review:the epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults[J].Aliment Pharmacol Ther,2011,34:274-285.
[3] Day C P,James O F.Steatohepatitis:a tale of two“hits”?[J].Gastroenterology,1998,114(4):842-845.
[4] Compare D,Coccoli P,Rocco A,et al.Gut-liver axis:the impact of gut microbiota on non-alcoholic fatty liver disease[J].Nutr Metab Cardiovasc Dis,2012,22(6):471-476.
[5] Mcorist A L,Abell G C,Cooke C,et a1.Bacterial population dynamics and faecal short-chain fatty acid(SCFA)concentrations in healthy humans[J].Br J Nutr,2008,100(1):138-146.
[6] Macfarlane G T,Gibson G R.Carbohydrate fermentation,energy transduction and gas metabolism in the human large intestine[J].Gastrointest Microbiol,1997,269-317.
[7] 王子花,申瑞玲,李文全.短鏈脂肪酸的產(chǎn)生及作用[J].畜牧獸醫(yī)科技信息,2007(2):12-13.
[8] 劉小華,李舒梅,熊躍玲.短鏈脂肪酸對腸道功效及其機制的研究進展[J].腸外與腸內(nèi)營養(yǎng),2012,19(1):56-58.
[9] Marco A R V,Hosana G R,Elaine H,et a1.Suppressive effect of short chain fatty acids on production of proinflammatory mediators by neutrophils[J].J Nutr Biochem,2011,22(9):849-855.
[10] Piekarska J,Mista D,Houszka M,et al.Trichinella spiralis:The influence of short chain fatty acids on the proliferation of lymphocytes,the goblet cell count and apoptosis in the mouse intestine[J].Exp Parasitol,2011,128(4):419-426.
[11] 梁蔭基,林琛蒞,王少娜,等.肥胖相關(guān)非酒精性脂肪性肝病防治的新靶點:短鏈脂肪酸及其受體信號通路的保護作用[J].重慶醫(yī)科大學(xué)學(xué)報,2016,41(6):628-631.
[12] Lin P,Lu J M,Wang Y F,et al.Naturally occurring stilbenoid TSG reverses non-alcoholic fatty liver diseases via gut-liver axis[J].PLoS ONE,2015,10(10):1-14.
[13] K.Sch?fer.Analysis of Short Chain Fatty Acids from different intestinal samples by capillary gas chromatography[J].Chromatographia,1995,40(9/10):550-556.
[14] Andreas Schwiertz,David Taras,Klaus Sch?fer,et al.Microbiota and SCFA in lean and overweight healthy subjects[J].Obesity,2010,18(1):190-195.
[15] Duncan S H,Louis P,Thomson J M,et al.The role of pH in determining the species composition of the human colonic microbiota[J].Environ Microbiol,2009,11(8):2112-2122.
[16] Nakamura N,Hamazaki T,Jokaji H,et a1.Effect of HMG-CoA reductase inhibitors on plasma polyunsaturated fatty acid concentrations in patients with hyperlipidemia[J].Int J Clin Lab Res,1998,28(3):192-195.
[17] Treem W R,Ahsan N,Shoup M,et a1.Fecal short-chain fatty acids in children with inflammatory bowel disease[J].J Pediatr Gastroenterol Nutr,1994,18(2):159-164.
[18] Kimura I,Ozawa K,Inoue D,et al.The gut microbiota suppresses insulin mediated fat accumulation via the short-chain fatty acid receptor GPR43[J].Nat Commun,2013,4:1829.
[19] Wostmann B S,Larkin C,Moriarty A,et a1.Dietary intake,energy metabolism,and excretory losses of adult male germfree Wistar rats[J].Lab Anim Sci,1983,33(1):46-50.
[20] Wolever T M,Brighenti F,Royall D,et a1.Effect of rectal infusion of short chain fatty acids in human subjects[J].Am J Gastroenterol,1989,84(9):1027-1033.
[21] Wolever T M,Spadafora P,Eshuis H.Interaction between colonic acetate and propionate in humans[J].Am J Clin Nutr,1991,53(3):681-687.
[22] Vernay M.Origin and utilization of volatile fatty acids and lactate in the rabbit:influence of the faecal excretion pattern[J].Br J Nutr,1987,57(3):371-381.
[23] Schwiertz A,Taras D,Sch?fer K,et al.Microbiota and SCFA in lean and overweight healthy subjects[J].Obesity(Silver Spring),2010,18(1):190-195.
[24] 張婷,林佩,陸建美,等.高脂飲食誘導(dǎo)大鼠高脂血癥模型的性別差異研究[J].實驗動物學(xué),2015,32(2):7-13.
[25] 易園驪,姚萍.脂代謝與腸道菌群關(guān)系的研究進展[J].醫(yī)學(xué)綜述,2013,19(6):991-994.
[26] 劉松珍,張雁,張名位,等.腸道短鏈脂肪酸產(chǎn)生機制及生理功能的研究進展[J].廣東農(nóng)業(yè)科學(xué),2013,(11):99-103.
[27] 鄭軍.腸道菌群失調(diào)與脂肪肝患者血脂和丙氨酸氨基轉(zhuǎn)移酶的變化研究[J].中國微生態(tài)學(xué)雜志,2007,19(2):161-162.
[28] 曹毅,沈峰,徐雷鳴,等.腸道菌群和內(nèi)毒素血癥與非酒精性脂肪性肝病[J].實用肝臟病雜志,2012,15(2):163-165.
EffectofRadixPolygoniMultifloriandTSGonShort-chainFattyAcidsinIntestinalTractofNAFLDRats△
WANG Yanfanga,LIN Peia,LU Jianmei,ZHANG Mei,LI Li,YANG Xingxin,YU Jie*
(YunnanUniversityofTraditionalChineseMedicine,Kunming650500,China)
Objective:To study the effect of Radix Polygoni Multiflori and its active ingredient,TSG,on short-chain fatty acids (SCFA),which were gut microbiota fermentation products of carbohydrates,in rats with nonalcoholic fatty liver disease.MethodsSD rats were randomly distributed into seven groups,mice were fed with either standard chow diet or high-fat diet,plus daily administration of water or water extract of Radix Polygoni Multiflori or TSG.The contents of acetic acid,propionic acid and butyric acid in stool samples were determined by gas chromatography.TC,TG contents in liver and blood sample,and endotoxin contents in hepatic portal venous blood were tested.ResultsThe water extract of Radix Polygoni Multiflori and TSG could reduce the contents of total SCFA in the intestinal tract of NAFLD mice fed by HFD,however,gender differences might existed.PMR and TSG could reduce acetic acid,propionic acid,butyric acid in HFD-fed male mice.At the same time,they relived high contents of TG,TC in liver tissue and LPS level in portal venous.In female rats,the propionic acid content was lower in PMR group compared with NAFLD rats.PMR also decreased the TC、TG level in hepatic tissues of female rats.However,low dose TSG raised acetic acid content,while reduced blood lipid and LPS level.In the meantime,regulation effect of PMRP on intestinal SCFA content of both female and male mice was not significant.ConclusionRadix Polygoni Multiflori and its active ingredient TSG affected SCFA level produced by gut microbial fermentation.This regulatory activity may contribute to their treatment activity to NAFLD.
Radix Polygoni Multiflori;TSG;short-chain fatty acids;non-alcoholic fatty liver;gas chromatography
10.13313/j.issn.1673-4890.2017.9.009
國家自然科學(xué)基金(81060337,81460623);云南省應(yīng)用基礎(chǔ)研究重點項目(2014FA035);云南省教育廳南藥協(xié)同創(chuàng)新中心項目(30270100500);云南省中青年學(xué)術(shù)和技術(shù)帶頭人后備人才項目(2015HB053);云南省應(yīng)用基礎(chǔ)研究計劃項目青年基金(2016FD050)
*
俞捷,副教授,研究方向:藥理學(xué)和藥物分析學(xué);Tel:(8710)65918055,E-mail:cz.yujie@gmail.com
a共同第一作者
2016-09-12)
