尼奧羅非魚雜種性別類型與性腺發育初步研究
2017-12-18 03:05:02趙永華趙金良陳曉武魏繼海傘桂寶簡偉業
水產科學 2017年4期
趙永華,趙金良,陳曉武,王 燕,魏繼海,傘桂寶,簡偉業
( 1.上海海洋大學 農業部淡水水產種質資源重點實驗室,上海 201306; 2.廣州市番禺區農業科學研究所,廣東 廣州 511450; 3.茂名市偉業羅非魚良種場,廣東 茂名 525000 )
尼奧羅非魚雜種性別類型與性腺發育初步研究
趙永華1,趙金良1,陳曉武1,王 燕1,魏繼海1,傘桂寶2,簡偉業3
( 1.上海海洋大學 農業部淡水水產種質資源重點實驗室,上海 201306; 2.廣州市番禺區農業科學研究所,廣東 廣州 511450; 3.茂名市偉業羅非魚良種場,廣東 茂名 525000 )
通過組織學方法,研究了0.5~4.5月齡尼奧羅非魚雜種(尼羅羅非魚♀×奧利亞羅非魚♂)的性腺發育特征與性別類型。結果表明,0.5月齡尼奧羅非魚原始性腺尚未分化;1月齡,精巢中原始生殖細胞開始分化成精原細胞;2.5月齡出現初級精母細胞;3月齡出現次級精母細胞;3.5~4月齡精巢中存在不同發育階段的生殖細胞,壺腹空腔中分散著稀疏的精子;4.5月齡精巢壺腹腔內密集大量精子。在2.5、3.5、4、4.5月齡抽樣中,均檢測到雌魚,2.5~3.5月齡卵巢以第Ⅱ時相卵母細胞為主;4~4.5月齡卵巢以第Ⅲ時相卵母細胞為主。研究結果初步表明,尼奧羅非魚雜種中觀察到雄性、雌性兩種性別類型,以雄性性別為主;雜種雌、雄魚的性腺均能正常發育。
尼奧羅非魚;性別類型;性腺發育;組織切片
羅非魚(Oreochromis)因其具有食性雜、生長快、繁殖力和抗病力強等優點,現已成為世界性養殖魚類[1]。由于羅非魚雄性比雌性生長速度快,為此,生產上一般采用雌性尼羅羅非魚(O.niloticus)和雄性奧利亞羅非魚(O.aureus)種間雜交的方式獲得雄性率高的雜交一代——尼奧羅非魚[2]。
有關尼奧羅非魚雄性率高的性別形成機制,早期認為,尼羅羅非魚是雄性異質型性染色體,雄性由異質的XY性染色體決定,雌性由同質的XX性染色體決定[3];奧利亞羅非魚是雌性異質型性染色體,雄性由同質的ZZ性染色體決定,雌性由異質的ZW性染色體決定[4],因此,雜交一代為XZ型雄性。除性染色體外,Avtalion等[5]提出,一條常染色體上的一對等位基因(AA或aa)影響性染色體,尼羅羅非魚雌性為AAXX,雄性為AAXY;奧利亞羅非魚雌性為aaZW,雄性為aaZZ,這個基因平衡模型假設,當性別決定有關的染色體和常染色體位點的作用超過某一閾值時,個體發育為雄性,低于這一閾值時則為雌性。
目前研究表明,大多數魚類不存在性染色體,性腺分化和發育受相關基因共同調控。羅非魚中已發現與性別分化相關的基因有很多,且定位于不同的染色體上,如尼羅羅非魚位于LG12染色體上的dmrt1基因,LG23染色體上的amh基因,LG1染色體上的cyp19a1基因,以及一些未成功定位的基因,這些單個或者多個基因共同參與羅非魚性別決定與分化調控[6]。此外,環境因子如溫度[7]、類固醇激素[8]等對羅非魚性別分化和發育也都有顯著影響。
已有的研究表明,尼羅羅非魚早期性腺發育中出現兩性性腺現象[9],也有學者提出雌性致死假設觀點,為進一步探明尼奧羅非魚雜交種性別偏離及性腺發育,本研究通過對0.5~4.5月齡尼奧羅非魚性腺組織學特征連續觀察,以期揭示尼奧羅非魚的性別類型、性腺發育特征。
1 材料與方法
尼奧羅非魚魚苗2015年5月28日采自廣州市番禺區農業科學研究所良種繁殖場,于上海海洋大學羅非魚種質資源試驗站的7.8 m×3.9 m×1.5 m水泥池中養殖,定時足量投喂,每個水泥池放養500尾,用羅非魚專用配合飼料養殖。
每隔15 d采樣一次,每次隨機采集30~40尾。常規生物學測量體長、體質量。幼魚前期,性腺細小無法剝取,截取性腺所在軀干部位,波恩氏液固定;當性腺可以剝取時,取完整性腺固定。組織經石蠟包埋,制成5~8 μm切片,蘇木精—伊紅染色,中性樹膠封片,于顯微鏡(Nikon DS-Ril)觀察拍照。用目微尺測量性腺橫截面的寬度和厚度以及生殖細胞的大小、核徑。卵巢、精巢的發育分期參照劉筠[10]分期標準。
試驗所得數據用Excel 2013軟件進行統計學分析,結果以平均值±標準差表示。
2 結 果
2.1 尼奧羅非魚的生長
0.5~4月齡尼奧羅非魚體長與體質量隨月齡的增加而顯著增加,體質量增長量隨月齡的增加而增大(表1)。
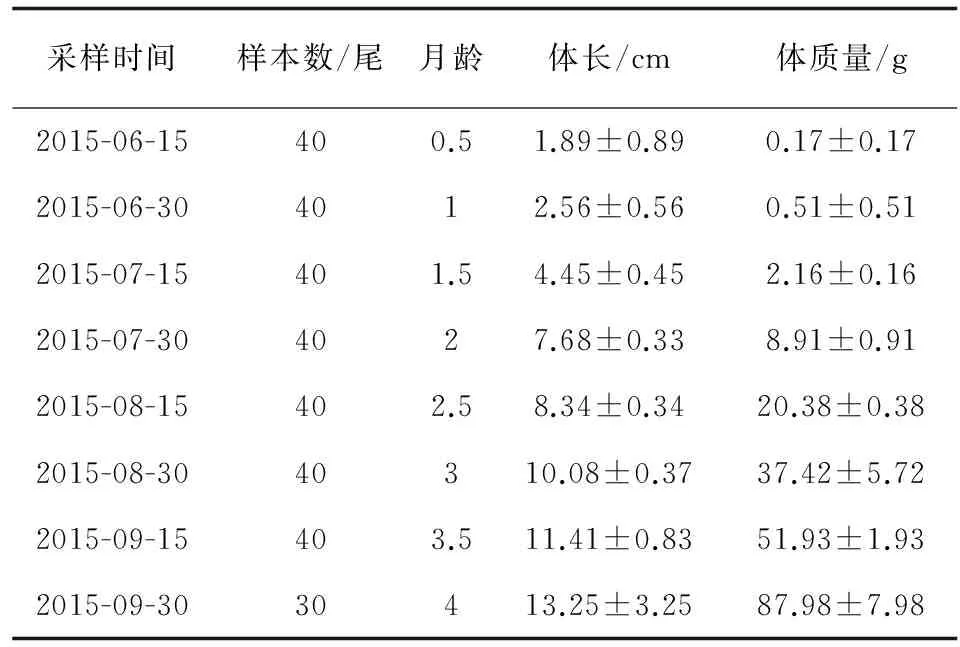
表1 尼奧羅非魚生長
2.2 尼奧羅非魚性腺發育
在9次采樣中,絕大多數為雄性個體,雌性個體僅在2.5、3.5、4、4.5月齡中檢測到,且在這4次采樣中分別只采集到1尾雌性個體,雄性率高達98.8%。尼奧羅非魚性腺大小隨生長而增大,同一批次采樣的卵巢體積比精巢大(表2)。
2.2.1 性腺未分化期
0.5月齡:性腺細小,肉眼無法辨別,性腺處于原始性腺階段。在顯微鏡下可觀察到性腺原基上布滿原始生殖細胞(3.21~5.15 μm),不能明顯區分精巢、卵巢(圖1a)。
2.2.2 精巢發育
Ⅰ期精巢(1~1.5月齡):精巢肉眼無法分辨,體積較小,白色。橫切面呈橢圓形,生殖細胞核較大,染色較深,細胞質色質較淡。1月齡,原生殖細胞開始分裂為精原細胞(圖1b);1.5月齡,精原細胞數目增多,無定向的分散在結締組織中,精原細胞直徑7.25~8.15 μm(圖1c)。
Ⅱ期精巢(2月齡):精巢體積增大1倍,呈白色線狀。部分單層精原細胞集中排列,形成精細小管,部分有管腔。精細小管大多呈圓形或橢圓形,直徑13.19~18.21 μm,其內的精原細胞細胞核位于中央或稍微偏離中心,精原細胞直徑6.15~7.94 μm,核徑1.6~2.3 μm(圖1d)。
Ⅲ期精巢(2.5月齡):精巢成肉色圓柱狀,表面有細小血管分布,體積繼續增大。精巢中產生數量較多的精原細胞,精小葉內出現空腔,精原細胞、初級精母細胞沿精小葉邊緣單層或多層排列,初級精母細胞直徑6.32~8.25 μm,核徑3.21~4.19 μm。精巢中形成1個大型空腔和1~2個小型空腔(圖1e)。
Ⅳ期精巢(3~4月齡):3月齡,精巢肉白色,扁平三棱柱狀,表面血管分布明顯。精小葉增大,其內可見由初級精母細胞、次級精母細胞及精細胞組成的精小囊,次級精母細胞直徑3.14~4.21 μm,核徑2.05~3.13 μm,此時小葉腔中已存在少量的精子,小型空腔數有所增加(圖1f)。3.5~4月齡,精巢呈肉色或肉白色,精巢背部帶有縱橫血管。細胞分裂較為明顯,部分精子通過精細小管壁經結締組織向小葉空腔游動,腔內分散著稀疏的精子(圖1g,h)。
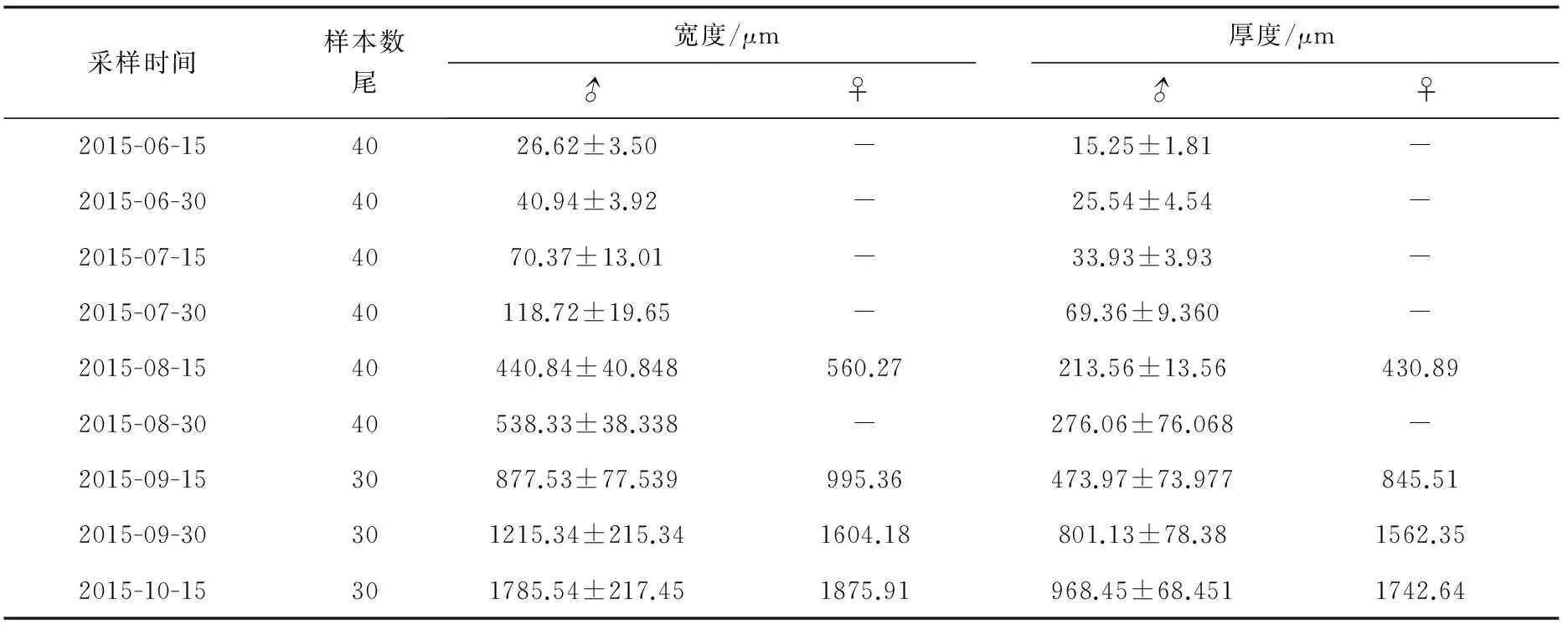
表2 尼奧羅非魚不同發育階段性腺寬度和厚度
Ⅴ期精巢(4.5月齡):精巢體積顯著增大,呈乳白色,擠壓腹部可擠出精子。小葉腔中一些相鄰精小囊壁互相融合,小葉空腔很大,其內充滿成熟的精子。精子嗜堿性極強,被蘇木精染成深藍色,直徑2.1~2.5 μm(圖1i)。
2.2.3 卵巢發育
Ⅱ期卵巢(2.5~3.5月齡):2.5月齡,卵巢呈白色細線狀,肉眼無法辨認,性腺末端未見血管分布。卵母細胞大小均勻,圓形不規則排列,細胞核居中央,占卵細胞一半左右,卵細胞直徑36.18~44.07 μm,核徑16.79~22.23 μm,核仁10個以上,不規則地分散在細胞核中,卵細胞外具有一層濾泡膜,以第Ⅱ時相卵母細胞為主(圖1j)。在3.5月齡卵巢中,卵母細胞直徑33.18~57.07 μm,核徑17.83~26.15 μm,出現少量卵黃核(圖1k)。
Ⅲ期卵巢(4~4.5月齡):4月齡,卵巢增大,呈囊狀,表面可見血管,肉眼已能夠看清細小的卵粒。卵母細胞不規則的排列在卵巢基質中,卵徑50~110 μm,核徑20~50 μm,核仁數增多沿核膜分布,外側由一層濾泡膜發育成二層濾泡膜,開始出現輻射帶,切片中以第Ⅲ時相卵母細胞為主,有少部分第Ⅱ時相卵母細胞(圖1l)。4.5月齡,卵巢表面血管增多,卵徑為50~200 μm,核徑40~95 μm,切片上出現少數Ⅳ時相卵母細胞,輻射帶增厚,細胞質中開始出現許多空泡,卵黃顆粒數量和體積增加(圖1m)。
3 討 論
3.1 性腺未分化期
性腺形成的標志是原始生殖細胞的出現。本研究中,0.5月齡尼奧羅非魚的性腺原基上已布滿原始生殖細胞,原始性腺已經形成,尚處于性腺未分化期,此時還不能區分精巢和卵巢,性腺平均寬度為(26.62±3.50) μm,平均厚度為(15.25±1.81) μm,略低于同期的尼羅羅非魚、莫桑比克羅非魚(O.mossambicus)性腺大小[11-12]。研究表明,孵化后10日齡,尼羅羅非魚性腺原基分化明顯,至15日齡,原始性腺已形成[11];莫桑比克羅非魚在孵化后10~11日齡,性腺原基分化明顯,生殖細胞數目迅速增加,逐漸形成原始性腺[12]。這些表明,尼奧羅非魚原始性腺形成與其他羅非魚較為接近,形成時間約為孵化后10~15 d完成。
3.2 性腺分化期

3.3 性腺分化發育完整性
在尼羅羅非魚性腺早期發育研究中,劉筠等[9,11]在Ⅰ、Ⅱ期精巢和卵巢中發現有精母細胞和卵母細胞共存的現象。但在本研究中,1~4.5月齡尼奧羅非魚的精巢(或卵巢)中均未觀察到兩性細胞共存的現象,性腺明顯分化為精巢或卵巢,未發現有“假雄”性別或“中間”性別類型。根據尼奧羅非魚成熟精巢中存在不同發育階段的生殖細胞,Ⅱ、Ⅲ期卵巢中同時存在多個時相的卵母細胞,初步認為,尼奧羅非魚的精巢、卵巢也屬于非同步發育型。
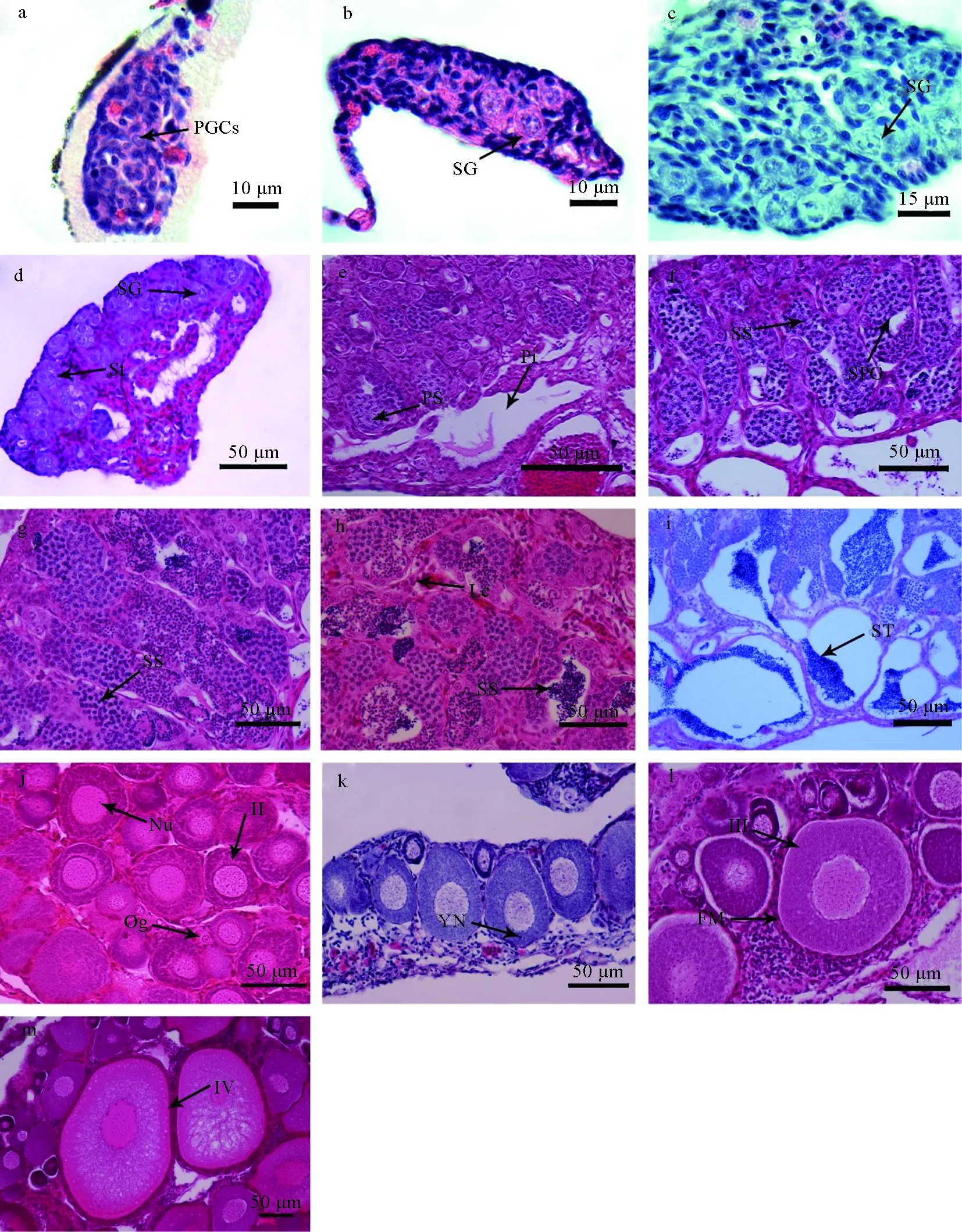
圖1 尼奧羅非魚性腺分化和發育
a:0.5月齡原始性腺,×100;b:1月齡Ⅰ期精巢,×100;c:1.5月齡精巢,×100;d:2月齡,Ⅱ期精巢,×40;e:2.5月齡,Ⅲ期精巢,×40;f:3月齡精巢,×40;g:3.5月齡精巢,×40;h:4月齡精巢,×40;i:4.5月齡,Ⅳ期精巢;j:2.5月齡,Ⅱ期卵巢,×40;k:3.5月齡卵巢,×40;l:4月齡,Ⅲ期卵巢,×40;m:4.5月齡,Ⅲ期卵巢,×40;PGCs:原始生殖細胞;SG:精原細胞;PS:初級精母細胞;SPG:精小囊;SS:次級精母細胞;St:精細小管;ST:精子細胞;Pi:壺腹腔;Og:卵原細胞;Nu:核仁;YN:卵黃核;FM:濾泡膜.
本研究結果初步表明,尼奧羅非魚存在雌、雄兩種性別類型,以雄性性別占主導(98%),雄魚中精原細胞、初級精母細胞、次級精母細胞、精子細胞正常發育,能夠產生成熟精子。據此認為,種間雜交對尼奧羅非魚的性腺分化與生殖細胞發育并未產生明顯影響,但雜交對雜種的精子數量、活力影響尚需進一步分析。許玉德等[17]對尼羅羅非魚、奧利亞羅非魚及尼奧羅非魚精巢組織學觀察,發現尼奧羅非魚精巢內生殖細胞密度比親本略疏,間質組織相對發達,間質細胞數目相對較多。萬英杰等[18]對體質量400~500 g尼奧羅非魚雌魚解剖觀察發現,卵巢分正常型和萎縮型,萎縮型的雌魚數量約為正常型雌魚的3倍,由于萎縮型雌魚不能正常排卵,表現出“雄性化”現象。本研究中,由于雌魚數量過少,也未能觀察到“萎縮型”卵巢。今后,還需擴大樣本數量,分析探討雜交對雜種卵巢后期發育及卵母細胞成熟是否產生影響,理解尼奧羅非魚雜種雄性率高的生理機制。
在尼奧羅非魚性腺解剖中也發現,大多數個體的性腺為雙側對稱型,也有些個體為雙側不對稱型,一側短小,甚至還出現單側性腺發育,這類性腺比正常型短小,這種現象是與羅非魚性腺不同步發育有關,或是由于雜交引起的雜種不育性,還需進一步研究確定。
[1] 張紅燕, 袁永明, 賀艷輝, 等. 中國羅非魚生產與貿易現狀分析及建議[J].中國漁業經濟,2015,33(3):95-100.
[2] 梁仁杰, 王曉斌. 奧尼魚苗種雄性率的鑒定與生長測定試驗[J]. 水利漁業,2004,24(4):33.
[3] Müller-Belecke A, H?rstgen-Schwark G. Sex determination in tilapia (Oreochromisniloticus): sex ratios in homozygous gynogenetic progeny and their offspring[J]. Aquaculture,1995,137(1):57-65.
[4] Desprez D, Mélard C. Effect of ambient water temperature on sex determinism in the blue tilapia,Oreochromisaureus[J]. Aquaculture, 1998,162(1):79-84.
[5] Avtalion R R, Hammerman I S. Sex-determination inSarotherodon(Tilapia) Ⅰ.Introduction to the theory of autosomal influences[J]. The Israeli Journal of Aquaculture Bamidgeh,1978,30(4):110-115.
[6] 鄭堯, 王在照, 陳家長. 調控魚類性腺分化基因的研究進展[J]. 水生生物學報,2015, 39(4):798-809.
[7] 黨廣成,劉羽清,付佩勝,等. 高溫誘導對尼羅羅非魚性別分化及生長的影響[J]. 漁業科學進展,2011,32(5):32-37.
[8] Sun L N, Jiang X L, Xie Q P, et al. Transdifferentiation of differentiated ovary into functional testis by long-term treatment of aromatase inhibitor in Nile tilapia[J].Endocrinology,2014,155(4):1476-1488.
[9] 劉筠,劉國安, 陳淑群,等. 尼羅羅非魚性腺發育的研究[J]. 水生生物學集刊,1983,7(1):17-32.
[10] 劉筠. 魚類繁殖生物學[M]. 北京:中國農業出版社,1993:23-32.
[11] 王令玲, 仇潛如, 吳福煌. 尼羅羅非魚性腺發育的研究[J]. 淡水漁業,1986(2):1-4.
[12] 陳玉琳, 胡秀敏, 朱雅珠. 莫桑比克羅非魚幼魚的性腺發育與分化[J]. 水產學報,1980,4(4):313-320.
[13] 馬學坤, 柳學周, 溫海深, 等. 半滑舌鰨性腺分化的組織學觀察[J]. 海洋水產研究,2006,27(2):55-61.
[14] 代麗, 劉彬, 王志堅. 稀有鮈鯽性腺分化的組織學觀察[J]. 西南師范大學學報,2013,38(1):55-61.
[15] 蘇敏, 林丹軍, 尤永隆, 等. 黑脊倒刺鲃性腺早期分化的研究[J]. 淡水漁業,2008,38(1):62-66.
[16] 汪學杰, 田媛, 劉奕, 等. 雙須骨舌魚性腺發育的組織學觀察[J]. 水產學報,2014,38(4):524-530.
[17] 許玉德, 鐘建興, 鄭森林, 等. 單性羅非魚及其親本精巢的組織學和LDH同工酶的比較[J]. 廈門大學學報:自然科學版,1999,38(4):628-632.
[18] 萬英杰, 周之武. 雄性化奧尼魚卵巢生物學的初步研究[J]. 水利漁業,1992(2):28-29.
GendersandGonadalDevelopmentinHybridsofNileTilapiaOreochromisniloticus♀×O.aureus♂
ZHAO Yonghua1, ZHAO Jinliang1, CHEN Xiaowu1, WANG Yan1, WEI Jihai1,SAN Guibao2,JIAN Weiye3
( 1. Laboratory of Freshwater Fisheries Germplasm Resources, Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China; 2. Guangzhou Panyu Institute of Agricultural Sciences, Guangzhou 511450,China;3. Maoming Weiye Tilapia Seed Farm, Maoming 525000, China )
The gender type and gonadal development of 0.5—0.45 month old Nile tilapiaOreochromisniloticus(♀)×O.aureus(♂) were observed by microscopic histology. The results showed that primordial gonad of the tilapia was undifferentiated at 0.5-month-old.The primordial germ cells in the testis differentiated into spermatogonia around 1 month, primary spermatocytes occurred at the age of 2.5 month, while secondary spermatocytes presented around 3-month-old. The germ cells were present in different developmental stages in the testis and sparse sperm dispersed in ampulla cavity at the age of 3.5-4.5 months,while intensive sperm were dispersed in ampulla cavity around 4.5 months. Among the samples, female individuals were also detected at 2.5,3.5,4 and 4.5 months, respectively,the oocytes were in phase Ⅱat 2.5-3.5 months ovarian,and phase Ⅱat 4 and 4.5 month-old.The findings suggested that there were two kinds of gender in hybrids with the male dominated, and that the testis and ovarian in the hybrids developed normally.
Oreochromisniloticus♀×O.aureus♂;gender type; gonadal development; histological method
10.16378/j.cnki.1003-1111.2017.04.011
S965.125
A
1003-1111(2017)04-0467-05
2016-04-08;
2016-11-28.
國家現代農業產業技術體系專項資金資助項目(CARS-49);水產動物遺傳育種中心上海市協同創新中心資助項目(ZF1206);廣東省水產良種體系建設專項(2016).
趙永華(1990-),男,碩士研究生;研究方向:水產動物遺傳育種與繁殖. E-mail:1099875884@qq.com. 通訊作者:趙金良(1969-),男,教授;研究方向:動物遺傳育種與繁殖.E-mail:jlzhao@shou.edu.cn.
