hMASP2 DNA納米脂質(zhì)體對(duì)BCG感染小鼠肺組織T細(xì)胞亞群和AKT/pAKT蛋白表達(dá)的影響
2017-11-13 05:33:46張國(guó)超董信芳雒艷萍車團(tuán)結(jié)馬興銘
轉(zhuǎn)化醫(yī)學(xué)電子雜志 2017年10期
關(guān)鍵詞:小鼠
高 琪,張國(guó)超,何 琦,董信芳,雒艷萍,車團(tuán)結(jié),馬興銘
(1蘭州大學(xué)基礎(chǔ)醫(yī)學(xué)院,2甘肅省功能基因組與分子診斷重點(diǎn)實(shí)驗(yàn)室,甘肅蘭州730000)
hMASP2 DNA納米脂質(zhì)體對(duì)BCG感染小鼠肺組織T細(xì)胞亞群和AKT/pAKT蛋白表達(dá)的影響
高 琪1,張國(guó)超1,何 琦1,董信芳1,雒艷萍1,車團(tuán)結(jié)2,馬興銘1
(1蘭州大學(xué)基礎(chǔ)醫(yī)學(xué)院,2甘肅省功能基因組與分子診斷重點(diǎn)實(shí)驗(yàn)室,甘肅蘭州730000)
目的:構(gòu)建hMASP2 DNA納米脂質(zhì)體復(fù)合物,探討hMASP2 DNA納米脂質(zhì)體對(duì)BCG感染小鼠肺組織T細(xì)胞亞群和AKT/pAKT蛋白表達(dá)的影響.方法:構(gòu)建pcDNA3.1?hMASP2重組表達(dá)質(zhì)粒,以脂質(zhì)體作為載體,制備質(zhì)粒DNA納米脂質(zhì)體復(fù)合物并檢測(cè)Zeta電位和粒徑;經(jīng)鼻滴入1×109CFU BCG 100 μL,建立結(jié)核分枝桿菌感染小鼠模型,實(shí)驗(yàn)分DNA納米脂質(zhì)體組(pcDNA3.1?hMASP2組)和對(duì)照組,在感染當(dāng)天小鼠尾靜脈分別注射pcDNA3.1?hMASP2 200 μL(25 μg DNA)和PBS 200 μL,每3天注射一次,連續(xù)治療三周后處死小鼠;分離肺組織淋巴細(xì)胞,流式細(xì)胞儀檢測(cè)T細(xì)胞亞群;肺組織免疫組化分析AKT、pAKT蛋白表達(dá).結(jié)果:pcDNA3.1?hMASP2 DNA納米脂質(zhì)體復(fù)合物Zeta電位52.6 mV、平均粒徑279.9 nm;與對(duì)照組相比,pcDNA3.1?hMASP2組小鼠肺組織CD4+T細(xì)胞亞群百分比明顯升高(P<0.05)、CD4+PD?1+T細(xì)胞亞群百分比降低(P<0.05);pcDNA3.1?hMASP2組小鼠肺組織AKT和pAKT的陽(yáng)性細(xì)胞百分比明顯增高(P<0.05).結(jié)論:構(gòu)建穩(wěn)定的pcDNA3.1?hMASP2 DNA納米脂質(zhì)體復(fù)合物,pcDNA3.1?hMASP2納米脂質(zhì)體增加BCG感染小鼠的肺組織的CD4+T細(xì)胞、促進(jìn)肺組織pAKT蛋白的表達(dá),降低肺組織的PD?1+T細(xì)胞亞群,從而上調(diào)小鼠T細(xì)胞功能.
納米脂質(zhì)體;MASP2;結(jié)核病;PD?1;AKT
0 引言
結(jié)核病(tuberculosis,TB)現(xiàn)已成為備受大家關(guān)注的全球性健康問(wèn)題.2015年,世界各地估計(jì)有1040萬(wàn)例新發(fā)活動(dòng)性結(jié)核病,同年全球造成約140萬(wàn)人死亡[1].結(jié)核病是嚴(yán)重危害人類健康的經(jīng)呼吸道傳播的慢性傳染病,近年來(lái),我國(guó)每年報(bào)告肺結(jié)核發(fā)病人數(shù)約100萬(wàn),始終位居全國(guó)甲乙類傳染病的前列;耐多藥肺結(jié)核危害日益凸顯,每年新發(fā)患者人數(shù)約12萬(wàn),未來(lái)數(shù)年內(nèi)可能出現(xiàn)以耐藥菌為主的結(jié)核病流行態(tài)勢(shì)[2].現(xiàn)如今,在藥物治療的基礎(chǔ)上,免疫學(xué)療法為結(jié)核病的治療提供了新方法和新思路,其中結(jié)核分枝桿菌疫苗和IL?12使用最多,DNA疫苗以Ag85B效果最好,它能夠刺激肉芽腫組織淋巴細(xì)胞的應(yīng)答,達(dá)到延長(zhǎng)動(dòng)物壽命的目的[3].IL?12能夠推進(jìn)CTL細(xì)胞成熟、引誘Th1分化、刺激T細(xì)胞、釋放IFN?γ,免疫治療具有較大應(yīng)用潛力.
補(bǔ)體系統(tǒng)被認(rèn)為是在炎癥過(guò)程中具有重要作用的先天免疫系統(tǒng)的重要組成部分.可以通過(guò)經(jīng)典途徑、替代途徑或凝集素途徑激活,產(chǎn)生具有廣泛免疫功能的C3a和C5a,以及能夠直接裂解靶細(xì)胞的末端補(bǔ)體裝置膜攻擊復(fù)合物[4].甘露糖結(jié)合凝集素相關(guān)絲氨酸蛋白酶2(mannan?binding lectin associated ser?ine proteases,MASP2)是啟動(dòng)補(bǔ)體凝集素途徑的關(guān)鍵效應(yīng)酶,其基因位于染色體1p36上,由12個(gè)外顯子組成,長(zhǎng)約20 kb[5-6].甘露糖結(jié)合凝集素(mannan?binding lectin,MBL)識(shí)別結(jié)合病原體后,與之結(jié)合的MASP2被激活,引發(fā)補(bǔ)體系統(tǒng)的抗體非依賴性活化途徑,裂解C4和C2并激活后續(xù)補(bǔ)體級(jí)聯(lián)反應(yīng),從而誘導(dǎo)炎癥反應(yīng),并破壞并清除病原體[5],因此,凝集素途徑效應(yīng)酶MASP2在發(fā)揮抗細(xì)菌感染方面具有重要的作用.
本實(shí)驗(yàn)室曾對(duì)MASP2蛋白進(jìn)行表達(dá)和純化,但其性質(zhì)不穩(wěn)定,在30 min即可降解,當(dāng)MASP2蛋白濃度>3 μg/L時(shí),在無(wú)MBL參與的情況下可發(fā)生自行激活,使得MASP2蛋白難以大量純化[7].在前期的研究中,以腺病毒為載體構(gòu)建的rAd?hMASP2對(duì)小鼠結(jié)核性肉芽腫的治療具有明顯治療效果[8-11],但腺病毒載體存在靶向性差、免疫原性強(qiáng)、半衰期短等缺點(diǎn),限制了其在治療中的應(yīng)用[12].為此,我們構(gòu)建MASP2 DNA納米脂質(zhì)體復(fù)合物,由磷脂、膽固醇等為膜材包合而成的脂質(zhì)體進(jìn)入人體內(nèi),被網(wǎng)狀內(nèi)皮系統(tǒng)吞噬而激活機(jī)體自身免疫功能,脂質(zhì)體容易浸染細(xì)胞,并能夠改變藥物的體內(nèi)分布,使藥物主要在肝、肺等組織器官中積蓄,提高藥物治療效率.因此,在本實(shí)驗(yàn)構(gòu)建hMASP2重組質(zhì)粒,以脂質(zhì)體為載體,制備DNA納米脂質(zhì)體復(fù)合物,評(píng)價(jià)其對(duì)BCG感染小鼠肺組織T細(xì)胞亞群和AKT/pAKT蛋白表達(dá)的影響.
1 材料和方法
1.1 hMASP2DNA納米脂質(zhì)體制備hMASP2全基因質(zhì)粒pYr?ads?1?hMASP2由本實(shí)驗(yàn)室構(gòu)建,E.coli DH5α和E.coli BL?21為本實(shí)驗(yàn)室保存;真核表達(dá)載體pcDNA3.1購(gòu)自蘇州金唯智有限公司;hMASP2的上下游引物由華大基因科技有限公司合成.以本實(shí)驗(yàn)室pYr?ads?1?hMASP2為模板進(jìn)行PCR擴(kuò)增,獲取hMASP2全基因,上游引物序列GGGGTACCGCCAC?CATGAGGCTGCTGACCCTCCT,下游引物序列GGAATTCTTAAAAATCACTAATTATGTTCTC,酶切位點(diǎn)EcoRⅠ和KpnⅠ,將構(gòu)建好的重組質(zhì)粒交由華大基因科技有限公司測(cè)序,結(jié)果顯示序列匹配,表明重組質(zhì)粒構(gòu)建成功.參考文獻(xiàn)方法[13]構(gòu)建hMASP2 DNA納米脂質(zhì)體復(fù)合物:用實(shí)驗(yàn)室已經(jīng)制備的脂質(zhì)體,調(diào)整其及質(zhì)粒DNA的濃度,將脂質(zhì)體與DNA 3∶1混合,剩余量用5%葡萄糖補(bǔ)足,用加樣槍反復(fù)吹打混勻,置于4℃冰箱中過(guò)夜,次日用于小鼠尾靜脈注射.
1.2 hMASP2 DNA納米脂質(zhì)體表征hMASP2 DNA納米脂質(zhì)體用Malvern Zetasizer Nano?ZS(Malv?ern Instruments,Worcestershire,UK)儀器檢測(cè)MASP2其Zeta電位和粒徑大小,判斷其穩(wěn)定性和納米脂質(zhì)體大小.
1.3 小鼠BCG感染模型建立與干預(yù)SPF級(jí)6~8周齡雌性昆明種小鼠(蘭州大學(xué)實(shí)驗(yàn)動(dòng)物中心),溫濕度適宜、干凈清潔的條件下飼養(yǎng).擴(kuò)增培養(yǎng)牛型結(jié)核分枝桿菌減毒株BCG D1331菌株(蘭州大學(xué)結(jié)核病研究中心惠贈(zèng)),第0天小鼠經(jīng)鼻滴入1×109CFU 100 μL,建立小鼠結(jié)核病模型并隨機(jī)將小鼠分為hMASP2脂質(zhì)體復(fù)合物組(pcDNA3.1?hMASP2組)和PBS對(duì)照組,每組四只.BCG感染當(dāng)天分別給予尾靜脈注射pcDNA3.1?hMASP2脂質(zhì)體復(fù)合物和等體積PBS,注射體積為200 μL(25 μg DNA),每隔三天注射一次.在第21天處死小鼠,取左肺組織分離淋巴細(xì)胞,流式細(xì)胞儀分析CD4、CD8、PD?1淋巴細(xì)胞亞群,右肺組織免疫組化染色檢測(cè)AKT和pAKT的表達(dá)情況,并計(jì)算其陽(yáng)性細(xì)胞百分比.
1.4 肺淋巴細(xì)胞分離與流式細(xì)胞術(shù)參考文獻(xiàn)方法[14]分離肺組織淋巴細(xì)胞,即用40%的Percoll(Pharmacia,America)分離小鼠肺組織的淋巴細(xì)胞,用D?Hanks液洗滌兩次,棄去上清后用PBS洗滌,100 μLPBS重懸后細(xì)胞計(jì)數(shù).調(diào)整細(xì)胞濃度,取2×106個(gè)淋巴細(xì)胞進(jìn)行染色,其中PE?抗CD4單克隆抗體(0.625 μL),APC?抗CD8單克隆抗體(0.625 μL),F(xiàn)ITC?抗PD?1單克隆抗體(0.8 μL)加入后(三株熒光抗體購(gòu)自eBioscience公司),輕輕渦旋混勻,4℃避光孵育30 min,然后加入預(yù)冷的PBS 1 mL進(jìn)行重懸洗滌,上機(jī)檢測(cè)(BD FACSCalibur),結(jié)果由FlowJo 7.6.1統(tǒng)計(jì)分析.
1.5 肺組織免疫組織化學(xué)染色用常規(guī)SABC免疫組化法檢測(cè)小鼠右肺組織中AKT、pAKT的表達(dá)水平,常規(guī)石蠟切片脫蠟,乙醇水化,檸檬酸鈉修復(fù),3%H2O2孵育10 min,加兔抗鼠一抗(1∶100)(抗AKT抗體購(gòu)自Bioss公司,抗pAKT抗體購(gòu)自bioswamp公司),4℃過(guò)夜孵育,加Maxvision二抗(1∶100)37℃孵育20 min,PBS沖洗2 min×3次,DAB顯色,蘇木素復(fù)染,脫水,封片,顯微鏡下觀察,自上向下隨機(jī)取10個(gè)高倍視野(×400),用Image?Pro plus 6.0軟件進(jìn)行分析,以細(xì)胞核內(nèi)出現(xiàn)棕黃色顆粒為陽(yáng)性信號(hào),計(jì)算陽(yáng)性細(xì)胞百分比.
1.6 統(tǒng)計(jì)學(xué)處理采用SPSS18.0軟件學(xué)進(jìn)行數(shù)據(jù)分析,計(jì)算出各組的M±SD,同時(shí)采用單因素方差分析計(jì)算P值,組間分析采用t檢驗(yàn),當(dāng)P<0.05時(shí)認(rèn)為差異有統(tǒng)計(jì)學(xué)意義.實(shí)驗(yàn)數(shù)據(jù)制圖使用Graph Pad Prism軟件.
2 結(jié)果
2.1 hMASP2 DNA納米脂質(zhì)體表征hMASP2 DNA納米脂質(zhì)體經(jīng)Malvern Zetasizer Nano?ZS(Malv?ern Instruments,Worcestershire,UK)儀器檢測(cè),測(cè)得其平均Zeta電位為52.6 mV,平均粒徑大小為279.9 nm(圖1).制備的hMASP2 DNA納米脂質(zhì)體復(fù)合物性質(zhì)穩(wěn)定(Zeta>40 mV,粒徑100~300 nm)[9],可用于小鼠尾靜脈注射.
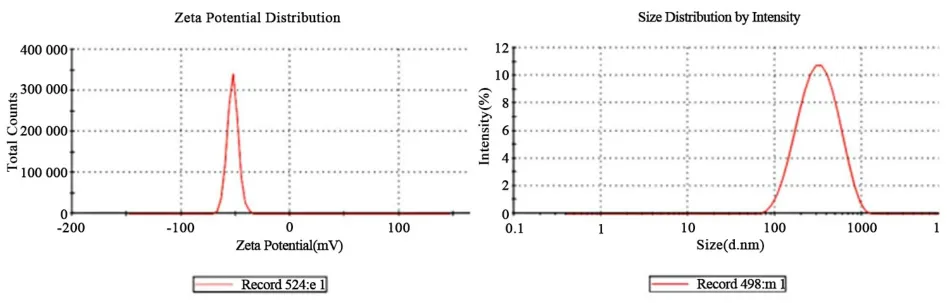
圖1 MASP2 DNA納米脂質(zhì)體Zeta電位和粒徑峰值
2.2 肺組織T細(xì)胞亞群流式細(xì)胞儀檢測(cè)小鼠肺組織T細(xì)胞數(shù)量、亞群及百分比結(jié)果見圖2~4.兩組小鼠肺組織T細(xì)胞數(shù)、CD8+T細(xì)胞亞群和CD8+PD?1+T細(xì)胞亞群百分比無(wú)統(tǒng)計(jì)學(xué)差異(P>0.05)(圖2、3).與對(duì)照組相比,pcDNA3.1?hMASP2組小鼠肺組織CD4+T細(xì)胞百分比明顯增多,差異具有統(tǒng)計(jì)學(xué)意義(P=0.028);pcDNA3.1?hMASP2組小鼠肺組織CD4+PD?1+T細(xì)胞百分比明顯降低,差異具有統(tǒng)計(jì)學(xué)意義(P=0.023).表明hMASP2 DNA納米脂質(zhì)體能夠促進(jìn)小鼠肺組織淋巴細(xì)胞的浸潤(rùn)和募集,并抑制CD4+T淋巴細(xì)胞膜表面抑制性分子PD?1的表達(dá).
圖2 兩組小鼠肺組織淋巴細(xì)胞總數(shù)
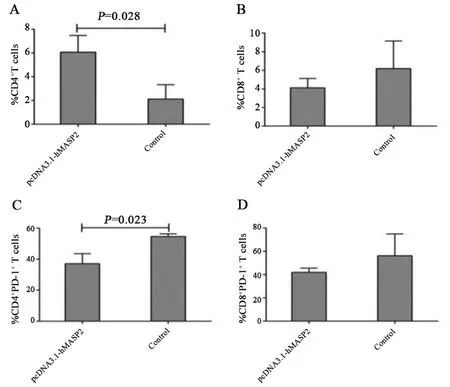
圖3 兩組小鼠肺組織CD4+、CD8+、CD4+PD?1+、CD8+PD?1+T細(xì)胞亞群百分比
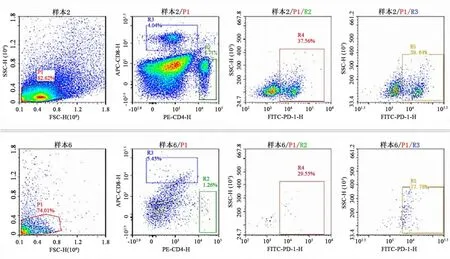
圖4 兩組小鼠肺組織淋巴細(xì)胞亞群流式細(xì)胞術(shù)檢測(cè)代表圖
2.3 肺組織免疫組化檢測(cè)每個(gè)切片觀察4個(gè)高倍鏡視野,每高倍鏡視野計(jì)數(shù)100個(gè)細(xì)胞,綜合判斷陽(yáng)性細(xì)胞染比例和染色強(qiáng)度.小鼠肺組織免疫組化檢測(cè)結(jié)果見圖5、6.與對(duì)照組相比,pcDNA3.1?hMASP2組AKT和pAKT陽(yáng)性細(xì)胞百分比明顯增高,差異具有統(tǒng)計(jì)學(xué)意義(P<0.05).對(duì)照組小鼠肺組織中有較少的棕黃色陽(yáng)性細(xì)胞分布,而hMASP2 DNA納米脂質(zhì)體組小鼠肺組織中有較多、集中分布的棕黃色陽(yáng)性細(xì)胞.hMASP2DNA納米脂質(zhì)體組小鼠肺組織AKT、pAKT的表達(dá)水平均顯著增加,表明hMASP2 DNA納米脂質(zhì)體能夠促進(jìn)小鼠肺組織細(xì)胞AKT表達(dá),并增加磷酸化的pAKT水平.
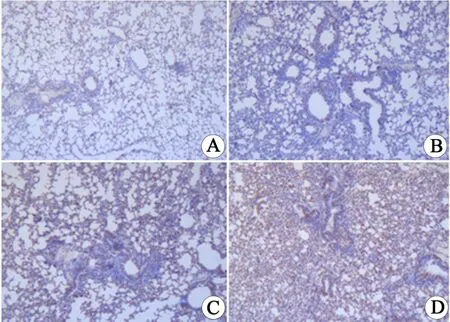
圖5 小鼠肺組織中AKT和pAKT免疫組化結(jié)果(IHC×100)
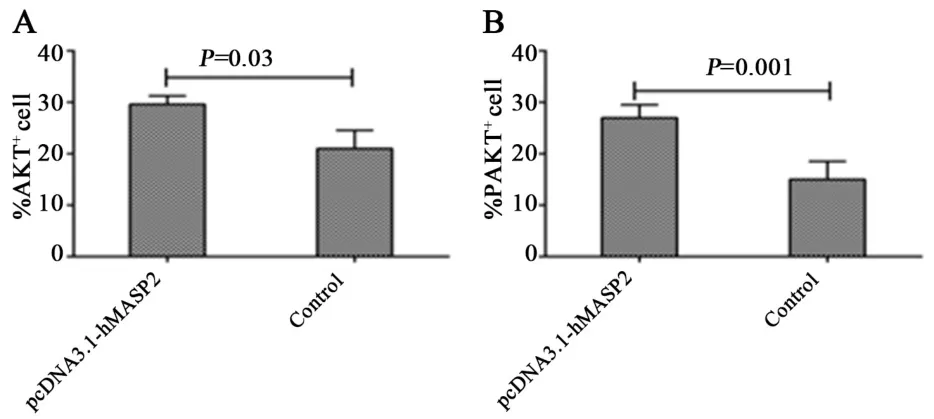
圖6 小鼠肺組織中AKT和pAKT陽(yáng)性細(xì)胞百分比
3 討論
程序性死亡因子(PD?1/CD279)是共刺激分子CD28家族中的受體,主要表達(dá)于T細(xì)胞表面,通過(guò)與其配體PDL?1/2結(jié)合,抑制淋巴細(xì)胞增殖和細(xì)胞因子產(chǎn)生[15].PD?1/PD?L1對(duì)調(diào)節(jié)T細(xì)胞功能起著至關(guān)重要的作用.在慢性感染、腫瘤患者體內(nèi),T細(xì)胞高表達(dá)PD?1分子,被認(rèn)為是T細(xì)胞耗竭的標(biāo)志[16-17].結(jié)核患者的免疫力受損與巨噬細(xì)胞和T細(xì)胞活化受損、IFN?γ的產(chǎn)生減少密切相關(guān)[18],結(jié)核分枝桿菌特異性CD4+T細(xì)胞在活動(dòng)性肺結(jié)核中的作用比CD8+T細(xì)胞更重要[19],并且活動(dòng)性肺結(jié)核CD4+T細(xì)胞高表達(dá)PD?1分子[20].故通過(guò)下調(diào)T細(xì)胞PD?1分子的表達(dá),改善患者的T細(xì)胞功能,是治療慢性感染的策略之一.在本實(shí)驗(yàn)中,pcDNA3.1?hMASP2治療組CD4+T細(xì)胞比例明顯升高、CD4+PD?1+T細(xì)胞比例降低,表明hMASP2 DNA納米脂質(zhì)體能夠促進(jìn)小鼠肺組織CD4+T細(xì)胞的浸潤(rùn)和募集,并抑制PD?1分子的表達(dá),從而上調(diào)CD4+T細(xì)胞功能.磷酸肌醇三磷酸激酶?絲氨酸?蘇氨酸蛋白激酶(phosphatidylinsitol 3?OH kinase?RAC?alphaserine/threonine?protein kinase,PI3K?AKT)信號(hào)通路廣泛存在于細(xì)胞中,是細(xì)胞內(nèi)重要信號(hào)通路之一,通過(guò)影響下游多種效應(yīng)分子的活化狀態(tài)發(fā)揮其作用[21].AKT也稱為蛋白激酶B,一種絲氨酸/蘇氨酸蛋白激酶,主要通過(guò)絲氨酸473和蘇氨酸308的磷酸化激活,在多種細(xì)胞過(guò)程中起重要作用[22].AKT是PI3K/AKT通路的中心環(huán)節(jié),可被PI3K激活成為具有磷酸激酶活性的pAKT(磷酸化AKT),pAKT可誘導(dǎo)干擾細(xì)胞凋亡功能的信號(hào),并且可能通過(guò)激活哺乳動(dòng)物的雷帕霉素靶標(biāo)和阻滯靶細(xì)胞通過(guò)正常的細(xì)胞周期等來(lái)促進(jìn)細(xì)胞存活,增殖和運(yùn)動(dòng)[23-24].在本實(shí)驗(yàn)中,pcDNA3.1?hMASP2組棕黃色的陽(yáng)性細(xì)胞分布更廣泛,并且AKT和pAKT陽(yáng)性細(xì)胞百分比明顯增高,其在小鼠肺組織中顯著表達(dá).一方面hMASP2 DNA納米脂質(zhì)體抑制BCG感染誘導(dǎo)的肺組織細(xì)胞凋亡,有利于肺組織修復(fù).另一方面,肺肉芽組組織中有大量浸潤(rùn)T細(xì)胞,hMASP2 DNA納米脂質(zhì)體通過(guò)影響AKT和pAKT表達(dá),促進(jìn)浸潤(rùn)T細(xì)胞增殖、活化,拮抗PD?1對(duì)T細(xì)胞的下調(diào)作用,可能是其下調(diào)PD?1分子的表達(dá)機(jī)制之一,繼而增強(qiáng)T細(xì)胞抗感染的效應(yīng)功能.總之,結(jié)核病的發(fā)生、發(fā)展、治療及預(yù)后是一個(gè)復(fù)雜的多因素參與的過(guò)程,本實(shí)驗(yàn)主要通過(guò)檢測(cè)PD?1和AKT水平來(lái)初步評(píng)價(jià)hMASP2 DNA納米脂質(zhì)體對(duì)小鼠結(jié)核性肉芽腫的影響.hMASP2 DNA納米脂質(zhì)體作用于小鼠后,T細(xì)胞PD?1表達(dá)水平降低,AKT和pAKT表達(dá)均增加,表明hMASP2 DNA納米脂質(zhì)體對(duì)BCG感染小鼠肺組織局部免疫具有上調(diào)作用.但hMASP在小鼠體內(nèi)的表達(dá)、hMASP2 DNA納米脂質(zhì)體對(duì)PD?1和AKT水平的調(diào)控機(jī)制需要進(jìn)一步研究.
[1] Yates TA,Khan PY,Knight GM,et al.The transmission of Myco?bacterium tuberculosis in high burden settings[J].Lancet Infect Dis,2016,16(2):227-238.
[2] 劉英姿.艾滋病患者中結(jié)核病的患病情況及發(fā)病趨勢(shì)分析[J].中國(guó)醫(yī)藥指南,2013,11(24):261-262.
[3]黃委剛.腫瘤壞死因子?α在結(jié)核病免疫中的研究進(jìn)展[J].醫(yī)藥前沿,2013(9):58-59.
[4] Cernoch M,Viklicky O.Complement in kidney transplantation[J].Front Med(Lausanne),2017,4:66.
[5] Wang Y,Yan J,Shi Y,et al.Lack of association between polymor?phisms of MASP2 and susceptibility to SARS coronavirus infection[J].BMC Infect Dis,2009,9:51.
[6] 王 倩,陳慧昱,陸小鈴,等.甘露糖結(jié)合凝集素相關(guān)絲氨酸蛋白酶2的研究進(jìn)展[J].細(xì)胞與分子免疫學(xué)雜志,2015,31(1):129-133.
[7] 張國(guó)超,雒艷萍,陸小鈴,等.重組腺病毒載體pAd?4?hMASP?2的構(gòu)建[J].中國(guó)生物制品學(xué)雜志,2016,29(11):1141-1145.
[8] Dong X,Luo Y,Gao Q,et al.Effects of MBL?associated serine pro?tease?2(MASP?2)on liquefaction and ulceration in rabbit skin mod?el of tuberculosis[J].Microb Pathog,2016,99:282-286.
[9] Gao Q,Dong X,Luo Y,et al.Construction of human MASP?2?CCP1/2SP,CCP2SP,SP plasmid DNA nanolipoplexes and the effects on tuberculosis in BCG?infected mice[J].Microb Pathog,2017,109:200-208.
[10] Xu X,Lu X,Dong X,et al.Effects of hMASP?2 on the formation of BCG infection?induced granuloma in the lungs of BALB/c mice[J].Sci Rep,2017,7(1):2300.
[11] Fu J,Wang J,Luo Y,et al.Association between MASP?2 gene pol?ymorphism and risk of infection diseases:A meta?analysis[J].Mi?crob Pathog,2016,100:221-228.
[12] 胡奇嬋,陳 玥,王 麗,等.腺病毒載體用于基因治療的研究進(jìn)展[J].解放軍醫(yī)藥雜志,2011,23(5):76-80.
[13] Templeton NS,Lasic DD,F(xiàn)rederik PM,et al.Improved DNA:lipo?some complexes for increased systemic delivery and gene expression[J].Nat Biotechnol,1997,15(7):647-652.
[14] Umemura M,Yahagi A,Hamada S,et al.IL?17?mediated regulation of innateand acquired immune responseag ainstpulmonary Mycobacte?rium bovis bacille Calmette?Guerin infection[J].J Immunol,2007,178(6):3786-3796.
[15] Freeman GJ,Wherry EJ,Ahmed R,et al.Reinvigorating exhausted HIV?specific T cells via PD?1?PD?1 ligand blockade[J].J Exp Med,2006,203(10):2223-2227.
[16] Pauken KE,Wherry EJ.Overcoming T cell exhaustion in infection and cancer[J].Trends Immunol,2015,36(4):265-276.
[17] Patel SP,Kurzrock R.PD?L1 expression as a predictive biomarker in cancer immunotherapy[J].Mol Cancer Ther,2015,14(4):847-856.
[18] Asante?Poku A,Yeboah?Manu D,Otchere ID,et al.Mycobacterium africanumis associated with patient ethnicity in Ghana[J].PLoS Negl Trop Dis,2015,9(1):e3370.
[19] Gao Y,Zhang S,Ou Q,et al.Characterization of CD4/CD8+αβ and VY2Vδ2+T cells in HIV?negative individuals with different My?cobacterium tuberculosis infection statuses[J].Hum Immunol,2015,76(11):801-807.
[20] Barber DL,Wherry EJ,Masopust D,et al.Restoring function in ex?hausted CD8 T cells during chronic viral infection[J].Nature,2006,439(7077):682-687.
[21] Landgren H,Curtis MA.Locating and labeling neural stemcells in the brain[J].J Cell Physiol,2011,226(1):1-7.
[22] Nicholson KM,Anderson NG.The protein kinase B/Akt signalling pathway in human malignancy[J].Cell Signal,2002,14(5):381-395.
[23] Xue G,Hemmings BA.PKB/Akt?dependent regulation of cell motil?ity[J].J Natl Cancer Inst,2013,105(6):393-404.
[24] Al?Bazz YO,Underwood JC,Brown BL,et al.Prognostic significance of Akt,phospho?Akt and BAD expression in primary breast cancer[J].Eur J Cancer,2009,45(4):694-704.
Effects of hMASP2 DNA nano-liposomes on T-lymphocytesubsetsandexpressionof AKT/pAKT protein in lung tissue of BCG-infected mice
GAO Qi1,ZHANG Guo-Chao1,HE Qi1,DONG Xin-Fang1,LUO Yan-Ping1,CHE Tuan-Jie2,MA Xing-Ming11Department of Immunology,School of Basic Medical Science,Lanzhou University,2Gansu Key Laboratory of Functional Genom?ics and Molecular Diagnosis,Lanzhou 730000,China
AIM:To evaluate the potential impacts of human MASP2(hMASP2)DNA nano liposomes on T?lymphocyte sub?sets and AKT/pAKT protein expression in lung tissue of BCG?in?fected mice through constructing the hMASP2 DNA nano?lipo?somes complex.METHODS:The pcDNA3.1?hMASP2 recombi?nant plasmids were constructed and then the hMASP2 DNA nano?liposomes complex were prepared with a carrier of liposome.The zeta potential and particle size of hMASP2 DNA nano?liposomes were detected.After infection with 100 μL of 1×109CFU BCG,mice were randomly divided into two groups and
the fol?lowing treatments:the mice of hMASP2 DNA nano?liposomes group were injected by tail vein with 200 μL(25 μg DNA)of DNA nano?liposomes and the mice of control group were received PBS(200 μL)by tail vein.All mice were injected every three days and treated for 3 weeks,then were killed.Lymphocytes were isolated from lung tissue and PD?1+T cell subsets were detected by flow cytometry.The expression of AKT/pAKT protein was detected with immunohistochemical staining.RESULTS:The zeta potential of pcDNA3.1?hMASP2 DNA nano?liposome complex was 52.6 mV and the average particle size was 279.9 nm.Com?pared with the control group,the percentage of CD4+T?lympho?cyte subsets in the lung tissue of mice treated with hMASP2 DNA nano?liposomes were significantly increased(P<0.05)and the percentage of CD4+PD?1+T?lymphocyte subsets were significantly decreased(P<0.05).The percentage of AKT and pAKT positive cells in hMASP2 DNA nano?liposomes group were significantly increased(P<0.05).CONCLUSION:A stable pcDNA3.1?hMASP2 DNA nano?liposome complex is firstly constructed,which shows the positive efficacy in increasing the number of CD4+T cells,reducing the number of PD?1+T?lymphocyte sub?sets,and promoting the expression of pAKT of BCG?infected mice.These findings provide experimental evidence that hMASP2 DNA nano?liposomes display a potential role of up?regulating T?cell?mediated immunity in tuberculosis.
nano?liposomes;MASP2;tuberculosis;PD?1;AKT
R730.45
A
2017-07-21;接受日期:2017-08-03
蘭州大學(xué)中央高校基本科研業(yè)務(wù)費(fèi)專項(xiàng)資金(lzujbky-2017-117);蘭州市科技計(jì)劃項(xiàng)目(2016-3-107)
高 琪.碩士.E?mail:gaoqijy@163.com
雒艷萍.副教授.馬興銘(共同通訊作者).教授.E?mail:maxm@lzu.edu.cn
2095?6894(2017)10?35?04
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
